熱脅迫下添加脯氨酸對(duì)菜心生長(zhǎng)和生理生化特性的影響
2020-11-16 02:06:21莊澤鑫周玲艷潘偉明
農(nóng)業(yè)與技術(shù) 2020年20期
關(guān)鍵詞:生長(zhǎng)
莊澤鑫 周玲艷 潘偉明

摘 要:本實(shí)驗(yàn)以菜心品種4號(hào)和31號(hào)為材料,測(cè)定了高溫脅迫以及高溫脅迫下添加外源脯氨酸對(duì)植株生長(zhǎng)情況如株高、根長(zhǎng)、鮮重、干重以及生理生化特性如葉綠素含量、丙二醛含量(MDA)過氧化氫酶( CAT) 和過氧化物酶( POD) 和超氧化物歧化酶(SOD)活性等各項(xiàng)指標(biāo)。結(jié)果表明:熱脅迫下,菜心品種31號(hào)的生長(zhǎng)狀態(tài)較好,葉綠素和脯氨酸含量相對(duì)較高,抗氧化酶POD、CAT酶活性較高,MDA活性、電導(dǎo)率相對(duì)較低;當(dāng)添加外源脯氨酸時(shí),可提高菜心的生長(zhǎng)指標(biāo)、葉綠素和脯氨酸含量以及POD、CAT、SOD酶活性,降低MDA活性和電導(dǎo)率,提升和降低的比例對(duì)于4號(hào)菜心品種而言更為明顯,說明脯氨酸含量可以作為菜心耐熱性的篩選指標(biāo)。
關(guān)鍵詞:菜心 熱脅迫 脯氨酸;生長(zhǎng);生理生化
中圖分類號(hào):S-3 ? ? ? 文獻(xiàn)標(biāo)識(shí)碼:A
DOI:10.19754/j.nyyjs.20201030004
菜心又稱菜苔,十字花科、蕓苔屬,一、二年生的草本植物。適合播種于南方溫暖地區(qū),是我國(guó)南方特產(chǎn)蔬菜之一[1]。菜心喜涼爽溫和氣候,生長(zhǎng)發(fā)育的適宜溫度為15~25℃,30℃以上高溫下菜心的生長(zhǎng)將受到較大的影響,會(huì)導(dǎo)致菜心纖細(xì),產(chǎn)量低,質(zhì)量差[2]。南方6—8月份高溫,嚴(yán)重影響菜心的生長(zhǎng),篩選和選育耐熱菜心品種非常必要,以滿足南方菜心的周年生產(chǎn)需求。
在旱、鹽堿、熱、冷、凍等逆境條件下,植物體內(nèi)脯氨酸的含量顯著增加[3-5]。在作物、蔬菜、花卉等多種植物中研究發(fā)現(xiàn),植物體內(nèi)脯氨酸含量在一定程度上反映了植物的抗逆性,抗逆性強(qiáng)的品種往往積累較多的脯氨酸[6-10]。廖飛雄等[11]發(fā)現(xiàn),通過在培養(yǎng)基中加入外源脯氨酸可降低熱脅迫下的愈傷組織細(xì)胞的相對(duì)電解質(zhì)滲出,減輕褐變,促進(jìn)鮮重的增加。本研究通過分析不同菜心品種高溫脅迫下脯氨酸含量以及電導(dǎo)率、葉綠素含量、丙二醛含量、抗氧化酶活性的變化,旨在為菜心耐熱品種的選育及抗逆栽培提供依據(jù)。
1 材料與方法
1.1 材料
供試材料為4號(hào)和31號(hào)菜心品種,購(gòu)買于廣東農(nóng)科院種子門市部。
1.2 方法
1.2.1 菜心種子的發(fā)芽和處理
取健康、飽滿的菜心種子,以質(zhì)量分?jǐn)?shù)2.0%的次氯酸消毒15min后,再用無(wú)菌去離子水洗3~4次,每次約30s。然后以無(wú)菌去離子水浸泡6h后,將適量的種子均勻的接于培養(yǎng)基中,將4號(hào)(A)和31號(hào)(B)菜心種子各分4組(A1、A2、A3、A4;B1、B2、B3、B4),具體情況見表1。其中,每組3瓶,總共24瓶。分別置于25℃和35℃的溫度培養(yǎng)箱中進(jìn)行光照培養(yǎng),每天光照12h,光照強(qiáng)度3000Lux。
1.2.2 菜心幼苗生長(zhǎng)指標(biāo)測(cè)定
菜心幼苗株高的測(cè)量采用百分尺從菜心的根部到葉尖。根長(zhǎng)的測(cè)量是從根尖到根部。以濾紙吸干菜心幼苗表面水分,用電子天平稱量鮮重。將菜心幼苗102℃殺青10 min后,置于70℃下烘干至恒重,用電子天平稱量幼苗的干重。每組濃度處理的菜心幼苗隨機(jī)抽取10株,計(jì)算平均值。
1.2.3 菜心幼苗生理生化指標(biāo)測(cè)定
葉綠素含量、游離脯氨酸含量、細(xì)胞膜透性、SOD、POD、CAT活性和MDA含量參照李合生等[12]方法。
1.2.4 游離脯氨酸含量的測(cè)定
參照張殿忠等[13]方法。
1.3 數(shù)據(jù)統(tǒng)計(jì)及分析
采用excel軟件計(jì)算,再對(duì)數(shù)據(jù)進(jìn)行作圖。
2 結(jié)果分析
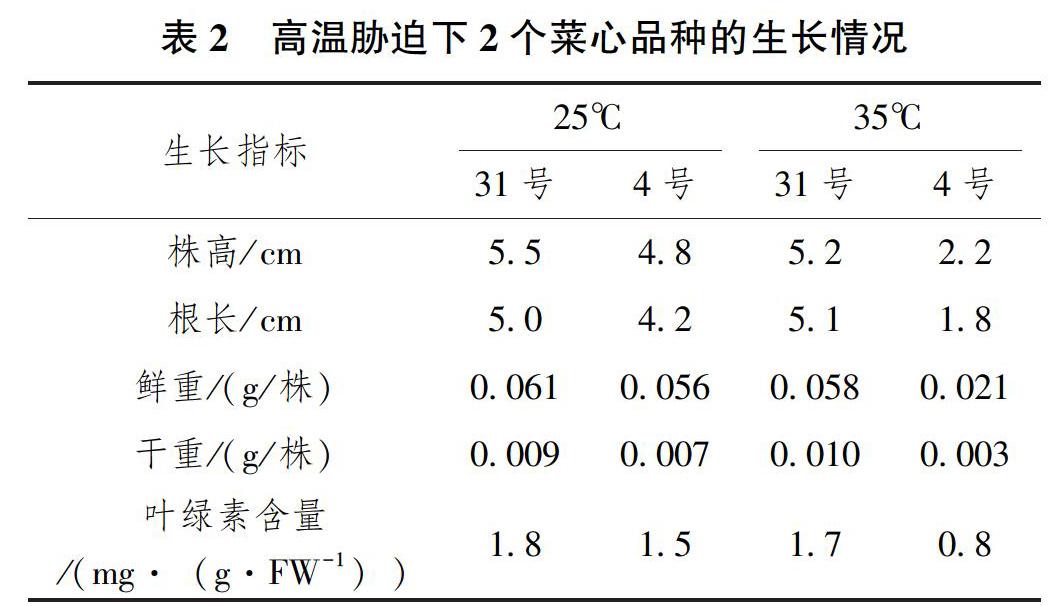
2.1 高溫脅迫對(duì)2個(gè)菜心品種生長(zhǎng)和葉綠素含量的影響 ?逆境脅迫往往影響植物的生長(zhǎng)發(fā)育,本研究將菜心種子接種于1/2MS培養(yǎng)基中,培養(yǎng)溫度分別為25℃和35℃,培養(yǎng)時(shí)間為5d,觀察和分析高溫脅迫下2個(gè)菜心品種31號(hào)和4號(hào)的生長(zhǎng)情況。結(jié)果表明,在正常培養(yǎng)溫度(25℃)下,2個(gè)菜心品種的生長(zhǎng)情況和生長(zhǎng)發(fā)育相關(guān)指標(biāo)相差不大,而在 35℃高溫培養(yǎng)下,菜心品種4號(hào)的生長(zhǎng)情況明顯差于31號(hào);且31號(hào)菜心品種與正常溫度(25℃)培養(yǎng)相比,差異較小,而4號(hào)菜心與正常溫度(25℃)培養(yǎng)相比,株高、根長(zhǎng)、鮮重、干重、葉綠素含量降低了54.2%、57.1%、62.5%、57.1%、46.7%(表2)。
2.2 高溫脅迫下2個(gè)菜心品種游離脯氨酸含量的變化
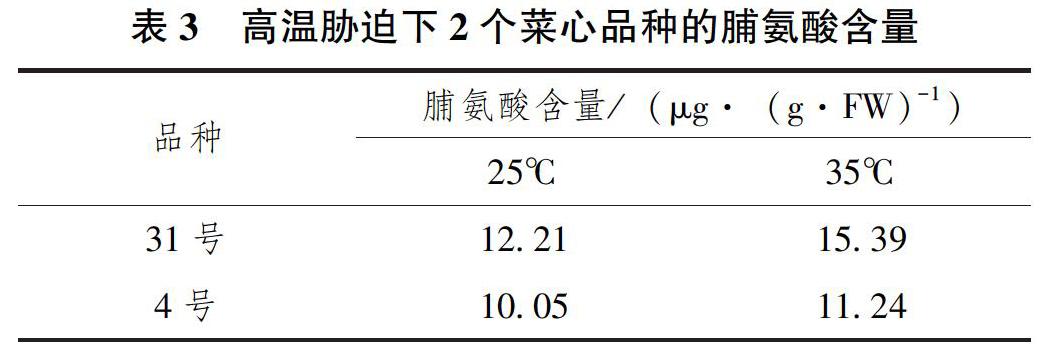
將常溫(25℃)發(fā)芽5d后的2個(gè)菜心品種放置于35℃培養(yǎng)箱中培養(yǎng)12h后,測(cè)定其游離Pro的含量。結(jié)果顯示,正常培養(yǎng)溫度(25℃)下,菜心品種31號(hào)的游離脯氨酸含量高于4號(hào)品種(21.5%);且高溫脅迫下,2個(gè)菜心品種的游離脯氨酸含量均有不同程度的增加,相比較對(duì)照(25℃),菜心品種31號(hào)的脯氨酸含量提升了26%,而4號(hào)品種只提升了11.8%(表3)。
2.3 高溫脅迫下外源脯氨酸對(duì)2個(gè)菜心品種生長(zhǎng)的影響 ?將菜心種子分別接種于1/2MS培養(yǎng)基(對(duì)照組)和添加1mg·L-1脯氨酸的1/2MS培養(yǎng)基(實(shí)驗(yàn)組)中發(fā)芽,培養(yǎng)條件均為35℃,培養(yǎng)時(shí)間為5d,然后測(cè)定2個(gè)菜心品種的各項(xiàng)生長(zhǎng)指標(biāo)和脯氨酸含量。結(jié)果表明,在較長(zhǎng)時(shí)期的高溫脅迫下,添加脯氨酸培養(yǎng)基中生長(zhǎng)的2個(gè)菜心品種的生長(zhǎng)指標(biāo)均高于對(duì)照組(表4),其中,31號(hào)的株高、根長(zhǎng)、鮮重、干重、葉綠素含量分別增加了15.4%、2.0%、17.2%、20%、17.6%,4號(hào)的分別增加了170%、167%、152%、133%、137%,且實(shí)驗(yàn)組在高溫脅迫下能夠正常生長(zhǎng)并保持一定的活力,而對(duì)照組則表現(xiàn)為部分枯萎。
2.4 高溫脅迫下外源脯氨酸對(duì)2個(gè)菜心品種脯氨酸含量的影響 ?將菜心種子分別接種于1/2MS培養(yǎng)基(對(duì)照組)和添加1mg·L-1脯氨酸的1/2MS培養(yǎng)基(實(shí)驗(yàn)組)中,25℃下發(fā)芽5d后,置于35℃培養(yǎng)箱中培養(yǎng)12h,測(cè)定其游離Pro的含量。結(jié)果表明,培養(yǎng)基中添加外源脯氨酸的情況下,31號(hào)和4號(hào)菜心品種的游離脯氨酸含量增高,分別提高了56%和66.5%(表5),從而提高了菜心的耐熱性。
2.5 熱脅迫下添加外源脯氨酸對(duì)2個(gè)菜心品種膜保護(hù)酶活性的影響 ?將菜心種子分別接種于1/2MS培養(yǎng)基(對(duì)照組)和添加1 mg·L-1脯氨酸的1/2MS培養(yǎng)基(實(shí)驗(yàn)組)中發(fā)芽,培養(yǎng)條件均為35℃,培養(yǎng)時(shí)間為5d,然后測(cè)定2個(gè)菜心品種的膜保護(hù)酶活性。計(jì)算處理測(cè)定值(添加脯氨酸)與對(duì)照測(cè)定值(不添加脯氨酸)的相對(duì)值,當(dāng)相對(duì)值大于1 時(shí)代表處理測(cè)定值高于對(duì)照測(cè)定值,相對(duì)值小于1 時(shí)代表處理測(cè)定值低于對(duì)照測(cè)定值,相對(duì)值等于1時(shí)代表處理測(cè)定值等于對(duì)照測(cè)定值。圖1結(jié)果表明,35℃高溫脅迫下,不添加外源脯氨酸時(shí),31號(hào)菜心的POD、CAT活性遠(yuǎn)遠(yuǎn)高于4號(hào)菜心(比值遠(yuǎn)遠(yuǎn)大于1),SOD酶活性相差不是很大,但MDA、電導(dǎo)率遠(yuǎn)遠(yuǎn)低于4號(hào)菜心(比值遠(yuǎn)遠(yuǎn)小于1),說明4號(hào)菜心耐熱性差,熱脅迫下,所受傷害大;當(dāng)添加外源脯氨酸時(shí),2個(gè)菜心品種的抗氧化酶活性及電導(dǎo)率差距大大縮小(比值接近1)。圖2結(jié)果表明,當(dāng)添加外源脯氨酸時(shí),2個(gè)品種的POD、CAT、SOD酶活性增加(比值均大于1),而MDA活性和電導(dǎo)率下降(比值均大于1),其中,4號(hào)菜心品種的POD、CAT、SOD酶分別是對(duì)照(沒有添加脯氨酸)的1.33、1.13、1.23倍,MDA和電導(dǎo)率分別0.7、0.62,31號(hào)菜心品種的POD、CAT、SOD酶分別是對(duì)照(沒有添加脯氨酸)的2.36、2.0、1.23倍,MDA和電導(dǎo)率分別0.51、0.28,4號(hào)菜心的升降幅度大于31號(hào)菜心。
3 討論
生物量是反映作物長(zhǎng)勢(shì)的重要農(nóng)藝性狀和形成作物產(chǎn)量的基礎(chǔ)[14],李榮華研究發(fā)現(xiàn),耐熱性強(qiáng)的菜心品種高溫脅迫下的生物量減少顯著低于耐熱性弱的菜心品種,并認(rèn)為生物量可以作為評(píng)價(jià)菜心耐熱性強(qiáng)弱的關(guān)鍵指標(biāo)[15]。本研究發(fā)現(xiàn),高溫脅迫下,耐熱性較好的菜心品種31號(hào)的株高、根長(zhǎng)、鮮重、干重均顯著優(yōu)于菜心品種4號(hào)。
葉綠素是進(jìn)行光合作用的主要色素。有研究表明,高溫脅迫下植物葉綠素含量下降,但耐熱性好的植物品種的葉綠素含量往往保持較高的水平[16-17]。本研究發(fā)現(xiàn),耐熱性較好的菜心品種31號(hào)的葉綠素含量高于菜心品種4號(hào),這可能也是31號(hào)菜心品種鮮重、干重較優(yōu)的原因。
逆境脅迫下植物將產(chǎn)生滲透調(diào)節(jié)物質(zhì),以提高對(duì)逆境的耐性。游離脯氨酸是重要的滲透調(diào)節(jié)物質(zhì)[18],具有提高細(xì)胞液濃度、維滲透壓、保護(hù)膜系統(tǒng)組分的作用[19]。脯氨酸含量一定程度上可以反映植物的耐熱性。毛靜等[20]發(fā)現(xiàn)高溫脅迫下鳶尾葉片脯氨酸含量不同程度的積累增加,且耐熱性較強(qiáng)的鳶尾品種無(wú)論常溫或高溫脅迫下的脯氨酸含量均高于耐熱性較弱的品種。靳路真等[18]在研究高溫脅迫對(duì)不同耐性大豆品種生理生化的影響時(shí)也有同樣的結(jié)論。本研究發(fā)現(xiàn),高溫脅迫下,耐熱性較好的菜心品種31號(hào)的脯氨酸含量高于菜心品種4號(hào),這與廖飛雄和潘瑞熾[11]的研究結(jié)果一致。
高溫脅迫下,往往導(dǎo)致大量 MDA 的產(chǎn)生,其含量多少可以直接反應(yīng)細(xì)胞膜受傷害程度。抗氧化酶SOD、POD、CAT是植物細(xì)胞內(nèi)活性氧清除系統(tǒng),具有保護(hù)細(xì)胞功能的作用[21]。靳路真等[12]發(fā)現(xiàn)高溫脅迫下,耐熱性品種SOD、POD 酶活性水平高于熱敏感品種。龐強(qiáng)強(qiáng)等[22]研究表明,隨著高溫脅迫時(shí)間的延長(zhǎng),菜心葉片中電導(dǎo)率、MDA 含量均呈上升的趨勢(shì),其中耐熱性強(qiáng)的菜心品種增幅較小,耐熱性差的菜心增幅較大,可能與耐熱性強(qiáng)的菜心葉片保護(hù)酶SOD、CAT和POD 活性顯著高于不耐熱品種有關(guān)。王宏輝等[23]在研究高溫脅迫對(duì)4個(gè)紅掌盆栽品種生理特性的影響時(shí)發(fā)現(xiàn),隨著高溫脅迫時(shí)間延長(zhǎng),4個(gè)品種的葉片相對(duì)電導(dǎo)率和MDA 呈上升趨勢(shì),SOD、CAT、POD呈現(xiàn)先上升后下降的趨勢(shì),這可能是由于高溫脅迫超過了植物能承受的極限,破壞酶活性中心,導(dǎo)致酶活性下降。廖飛雄和潘瑞熾[11]發(fā)現(xiàn),在培養(yǎng)基中添加脯氨酸后,菜心愈傷組織細(xì)胞的膜保護(hù)酶POD 、CAT 和 SOD 活性有不同程度的提高。本研究發(fā)現(xiàn)熱脅迫下,31號(hào)菜心的POD、CAT活性遠(yuǎn)遠(yuǎn)高于4號(hào)菜心,電導(dǎo)率遠(yuǎn)遠(yuǎn)低于4號(hào)菜心。當(dāng)添加外源脯氨酸時(shí),兩個(gè)菜心品種的抗氧化酶活性增加,MDA和電導(dǎo)率下降,且4號(hào)菜心的升降變化率大于31號(hào)菜心。
參考文獻(xiàn)
[1] 曾小玲,方淑桂,朱朝輝,等.不同基因型菜心游離小孢子培養(yǎng)和植株再生[J].熱帶作物學(xué)報(bào),2014,35(12):2397-2402.
[2]張振賢.蔬菜栽培學(xué)[M].北京:中國(guó)農(nóng)業(yè)大學(xué)出版社,2003.
[3]Hanson A,Hitz WD.Metabolic responses of mesophyte to plant water deficits[J].Annu Rev Plant Physiol,1982,33:163-203.
[4]湯章誠(chéng).逆境條件下植物脯氨酸積累及其可能的意義[J].植物生理學(xué)通訊,1984(01):15-21.
[5]徐同,陳翠蓮.植物抗逆性測(cè)定(脯氨酸快速測(cè)定)法[J].華中農(nóng)學(xué)院學(xué)報(bào),1983(01):94-95.
[6]Van Swaaij A C, Jacobsen E, Feenstra W J .Effect of cold hardening ,wilting and exogenously applied proline on leaf proline content and frost tolerance of several genotypes of Solanum[J].Physiol Plantarum ,1985(64):230-236.
[7]VanSwaaij A C,Jacobsen E,Kiel JAKW,et al.Selection characterization and regeneration of hydroxyproline-resistant cell lies of Solanum tuberosum:I Tolerance to NaCl and freezing stress[J].Physiol Plantarum,1986(68):359-366.
[8]Dorffling K, Dorffling H, Lesselich G.In vitro-selection and regeneration of hydroxyproline- resistant lines of winter wheat with increased proline content and increased frost tolerance[J].Journal of Plant Physiology,1993(142):222-225.
[9]孔令接,陳言博,王亞琴.不同夏菊品種的耐熱性研究.園藝學(xué)報(bào),2019,46(12):2437-2448.
[10]李威,肖熙鷗,呂玲玲.高溫脅迫下茄子耐熱性表現(xiàn)及耐熱指標(biāo)的篩選.熱帶作物學(xué)報(bào)2015,36(06):1142-1146.
[11]廖飛雄,潘瑞熾.熱脅迫下菜心脯氨酸含量變化及其在耐熱中的作用[J].華南師范大學(xué)學(xué)報(bào)(自然科學(xué)版),2001,(02):45-49.
[12]李合生,孫群,趙世杰.植物生理生化實(shí)驗(yàn)原理和技術(shù)[M].北京:高等教育出版社.2000.
[13]張殿忠,汪沛洪,趙會(huì)賢.測(cè)定小麥葉片游離氨基酸含量的方法[J].植物生理學(xué)通訊,1990(04):62-65.
[14]Bai J, Li S, Wang K, et al. Estimating above ground fresh biomass of different cotton canopy types with homogeneity models based on hyper spectrum parameters[J].Agricultural Sciences in China,2007,6(04):437-445.
[15]李榮華,郭培國(guó),張華.高溫脅迫對(duì)菜心農(nóng)藝性狀的影響[J].長(zhǎng)江蔬菜,2011(22):39-43.
[16]LiuX,Huang B.Heat stress injury in relation to membrane lipid peroxidation in creeping bent grass[J] .Crop Science,2000,40(02):503-510.
[17]靳路真,王洋,張偉.高溫脅迫對(duì)不同耐性大豆品種生理生化的影響.大豆科學(xué),2019,38(01):063-071.
[18]廉勇,崔世茂,包秀霞,等.根區(qū)溫度對(duì)辣椒幼苗生理特性的影響[J].華北農(nóng)學(xué)報(bào),2014,29(05):156-160.
[19]李春燕,陳思思,徐雯,等.苗期低溫脅迫對(duì)揚(yáng)麥葉片抗氧化酶和滲透調(diào)節(jié)物質(zhì)的影響[J].作物學(xué)報(bào),2011(37):2293-2298.
[20]毛靜,董艷芳,周媛,等.十六個(gè)德國(guó)鳶尾品種的耐熱性評(píng)價(jià)[J].北方園藝,2019(03):94-100.
[21]李珍珍,韓陽(yáng).抗壞血酸對(duì)小麥種子老化及幼苗脂質(zhì)過氧化的影響[J].遼寧大學(xué)學(xué)報(bào)(自然科學(xué)版),2000,23(05):56-61.
[22]龐強(qiáng)強(qiáng),周曼,孫曉東.菜心耐熱性評(píng)價(jià)及酶促抗氧化系統(tǒng)對(duì)高溫脅迫的響應(yīng)[J].浙江農(nóng)業(yè)學(xué)報(bào),2020,32(01):72-79.
[23]王宏輝,顧俊杰,房偉民,等.高溫脅迫對(duì)4個(gè)紅掌盆栽品種生理特性的影響[J].華北農(nóng)學(xué)報(bào),2016,31(02):139-145.
(責(zé)任編輯 周康)
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
