大氣低溫脅迫對中國褐飛虱年內初始遷入的影響
2020-11-13 02:02:10包云軒蒼薪竹楊詩俊謝曉金陸明紅劉萬才
生態學報 2020年20期
包云軒,蒼薪竹,楊詩俊,陳 粲,謝曉金,陸明紅,劉萬才
1 氣象災害預報預警與評估協同創新中心/南京信息工程大學, 南京 210044 2 江蘇省農業氣象重點實驗室/南京信息工程大學, 南京 210044 3 全國農業技術推廣服務中心/農業農村部, 北京 100125
稻飛虱是一類危害極重的專食性水稻害蟲,褐飛虱(Nilaparvatalugens(St?l))是三種主要稻飛虱中產生危害最嚴重的一種,它們可借助高空氣流實現種群大規模遠距離的遷飛,由此產生毀滅級的災害[1- 2]。自20世紀60年代初期,東亞和東南亞熱帶和亞熱帶地區實行“綠色革命”以來,在感蟲性水稻品種種植面積快速擴增、產量快速增長的同時,于20世紀60年代后期至90年代初出現了第一輪蟲害爆發周期。之后這些地區濫用殺蟲劑、過量施用氮肥等行為導致褐飛虱于21世紀00年代中期至今進入第二輪大爆發[3]。常年從東南亞地區頻繁且大量遷入到中國境內的褐飛虱嚴重地影響了中國水稻的品質和產量[4-5],由此造成中國水稻年均減產可達數億噸[6],嚴重威脅了中國的糧食安全。
褐飛虱年內初期遷入我國的情況往往可以決定整年蟲害的發生程度[7]。華南稻區和西南稻區作為每年年初境外褐飛虱種群遷入我國的第一站,其褐飛虱的春季發生量與全國發生面積、發生程度有著至關重要的聯系[8],遷入期早、遷入峰期提前也加大了我國南方稻區早稻上褐飛虱種群爆發的風險[9]。
大氣溫度對褐飛虱遷入期的早遲有著十分重要的影響,這是因為大氣溫度可以限制褐飛虱完整的遷飛行為[10]。例如,近地面溫度在13℃以下時大部分褐飛虱不會起飛,起飛后的褐飛虱種群也多集中在溫度范圍在16—18℃的對流層大氣中飛行[11- 12],遇低溫則出現集中降落的行為等。其中,促使褐飛虱發生降落行為的低溫屏障往往是南北方的冷暖氣團交匯形成的低溫墻[13]。穩定的低溫層結可迫使褐飛虱逐漸接近地面或向上遇冷停止振翅從而導致其直接降落。有研究表明,全球變暖引發的溫度普遍升高間接使褐飛虱年內初期遷入我國的時間提前,其中一部分原因是蟲源地溫度的升高更利于褐飛虱種群的遷出[14];而另一部分原因則是我國西南、華南兩稻區地處熱帶與亞熱帶的過渡地帶,初春時節氣溫變化復雜,極易產生低溫層結,而溫度的升高極有可能弱化了低溫屏障的限制作用。因此,研究大氣低溫脅迫對境外褐飛虱年內初始遷入中國的影響對我國褐飛虱種群早期入侵的預警具有重要的理論價值和實踐意義。但目前相關的研究多以春季遷入個例為研究對象[13, 15],幾乎沒有對多年褐飛虱初始遷入我國過程中的低溫脅迫發生特征進行過系統性、規律性的總結與討論。
在現今全球變暖愈演愈烈、極端氣候事件頻發的大背景下,氣候異常增加了溫度對褐飛虱遷飛行為影響的不確定性,同時,大氣溫度對褐飛虱遷飛過程的影響程度和作用機理也趨于復雜化。因此,系統性地了解歷史上褐飛虱年內初始遷入我國時的低溫脅迫發生規律,明確當前發生特征,對預測未來氣候變化情景下可能的發生趨勢顯得十分重要和迫切。本文擬利用中國多年褐飛虱遷入的蟲情數據,結合HYSPLIT軌跡模式,對2000—2017年境外褐飛虱年內初始遷入中國華南和西南稻區的發生特征進行統計分析和遷入過程模擬,以探明大氣低溫對褐飛虱遷入的影響,揭示低溫脅迫作用的時空差異。研究將有助于監測境外褐飛虱初始遷入我國的動態,為全球變暖趨勢下褐飛虱遷入發生的準確預測奠定基礎。
1 材料與方法
1.1 資料來源
(1)蟲情資料:稻飛虱蟲情資料由全國農業技術推廣服務中心提供,文中主要使用了云南省、廣西壯族自治區、廣東省2000—2017年褐飛虱逐日燈誘數據。
(2)氣象資料:本研究所用的氣象資料為美國國家環境預報中心(National Center for Environmental Prediction,NCEP)發布的全球6 h一次(UTC 00、06、12和18時),空間分辨率為1°×1°的FNL氣象再分析資料。
(3)其他資料:全球行政區劃底圖(1∶4000000),來源于中國國家基礎地理信息中心;30 m全球地表覆蓋數據,來源于中國地理信息資源目錄服務系統。
1.2 研究方法
1.2.1調查方法
褐飛虱僅在我國華南南部和西南南部的少數地區存在越冬蟲源,而在每年早春遷入我國南方稻區的蟲源大多是從中南半島等地越境遷入[16]。因此,選取緊鄰中南半島的西南稻區(以云南省為代表)、華南稻區(以廣東省、廣西壯族自治區為代表)為研究區,使用稻區內各植保站點的每日燈誘蟲情數據進行大氣低溫脅迫對褐飛虱年內初始遷入行為影響的研究(圖1)。
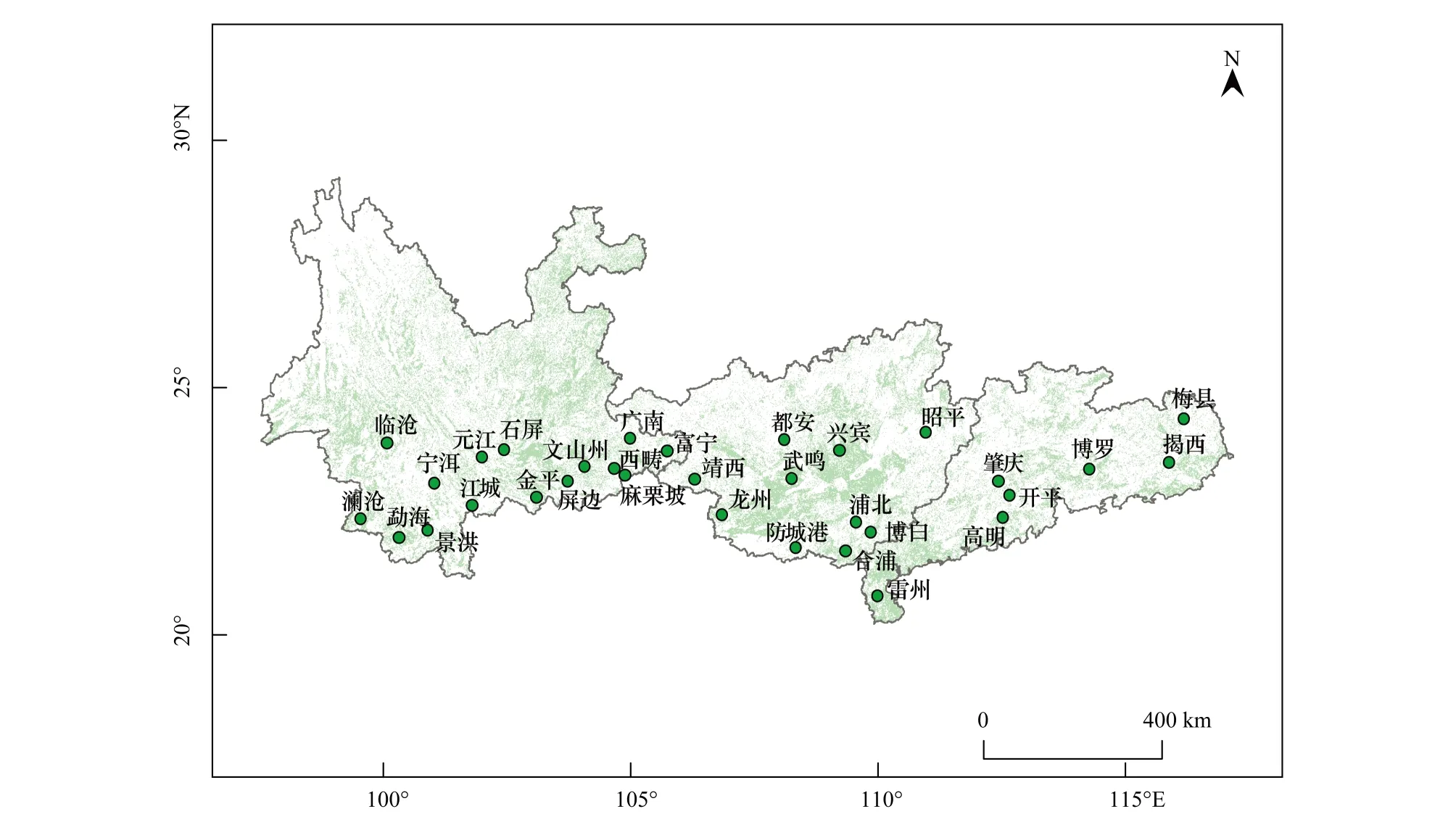
圖1 西南、華南稻區褐飛虱監測植保站點分布Fig.1 Distribution of N. lugens′ monitoring plant protection stations in the rice-growing regions of the southwest China and the south China
綜合考慮年內初始遷入的蟲情發生情況,并參考2009年發布的《稻飛虱測報調查規范》對始見期及高峰日的定義,將研究區年內褐飛虱首次單燈單日有蟲的日期定為始見期,年內單燈單日蟲量首次突增超過100頭為首次遷入峰發生日,并以此統計出2000—2017年兩個稻區的始見期和首次遷入峰日期的情況。
1.2.2軌跡分析方法
軌跡回推分析采用的是美國國家海洋和大氣管理局(National Oceanic and Atmospheric Administration,NOAA)與澳大利亞氣象局(Bureau of Meteorology,BOM)開發的HYSPLIT(Hybrid Single Particle Lagrangian Integrated Trajectory)模型。參數設置如下:(1)褐飛虱順風遷飛,其主動飛行能力是可忽略項[17-18];(2)軌跡回推到的蟲源附近曾存在理論上可提供蟲源取食的稻區;(3)褐飛虱在日出前1 h和日落后1 h內起飛[19-20]。西南、華南兩稻區3—4月份日出時刻在BST 07:00左右,日落時刻在BST 19:00左右。春季早晚氣溫較低,褐飛虱一般集中在較溫暖的下午飛行,屬于“日間單峰起飛”型[20]。故設定燈下見蟲日當晚20:00、燈下見蟲第二日清晨06:00兩個起始時刻,分別回推24 h、10 h至前一日晚20:00。(4)褐飛虱的盛行遷飛高度位于海拔1500 m左右[21],特別說明:在模型中設置的起始高度為離地高度。西南稻區地勢高且水稻種植區在海拔幾百米至幾千米的范圍內均有所分布[22],在離地高度相同的條件下,大氣溫度遠低于同緯度地區。而褐飛虱無法在環境溫度過低的條件下飛行,因此,模型中設置的西南稻區的離地高度相對較低。可見,褐飛虱的遷飛高度因受到溫度、地形等條件限制,需要根據季節及站點海拔的不同進行相應的調整,模型中兩稻區遷飛軌跡起始高度設定如表1。

表1 基于HYSPLIT模型的西南、華南稻區遷飛軌跡起始高度設定
1.2.3數據統計方法
為了解年內初始遷入過程中低溫脅迫發生的整體情況,文中使用Excel等軟件統計2000—2017年中低溫屏障發生頻率、低溫屏障強度、低溫屏障幅度、低溫屏障出現時間分布等特征,并作相關描述。各特征具體判定方法如下:(1)低溫屏障:假設褐飛虱所在飛行高度層環境溫度低于16℃時,褐飛虱即停止飛行[11];(2)假設降蟲區整晚均有降蟲。針對不同高度層,若降蟲區在降蟲當晚最低氣溫小于16℃,即判定存在低溫屏障。18 a中出現低溫屏障的次數除以總年數記為低溫屏障發生頻率。僅針對受低溫脅迫的降蟲過程計算低溫強度和降溫幅度。(3)低溫強度:針對不同高度層,在降蟲區降蟲當晚20:00、次日02:00、次日08:00(記蟲)三個時刻中出現的最低溫度記為低溫強度;降溫幅度:將盛行遷飛高度層(西南稻區800 hPa,華南稻區850 hPa)上降蟲區降蟲當晚的最大溫差記為降溫幅度。
2 結果與分析
2.1 初始遷入時期蟲情統計結果
2.1.1始見期統計結果
2000—2017年間,褐飛虱在我國西南稻區最早于3月10日始見,華南稻區最早于3月1日始見(表2)。在西南稻區內所有站點的平均始見期為4月24日±2.15日(平均始見期平均值±標準誤差),華南稻區所有站點的平均始見期為4月10日±2.72日。在18 a間,褐飛虱始見于華南稻區的日期較同緯度的西南稻區整體早9—14 d。2001—2008年西南稻區內最早始見期出現在4月10日前后,2009—2014年提前至4月初,到2015—2017年則穩定在3月10日。2000—2003年華南稻區內最早始見期在4月初,2004—2009年提前至3月上旬,到2010—2017年則穩定在3月1日。如圖2中所示,在同一稻區的始見期,盡管年間存在波動,整體上仍呈顯著提前的趨勢(始見期與年份呈顯著負相關,L1:r1=-0.803,P<0.01;L2:r2=-0.801,P<0.01)。相關研究表明,這種提前趨勢可能與近30 a的全球氣候變暖有關[14]。
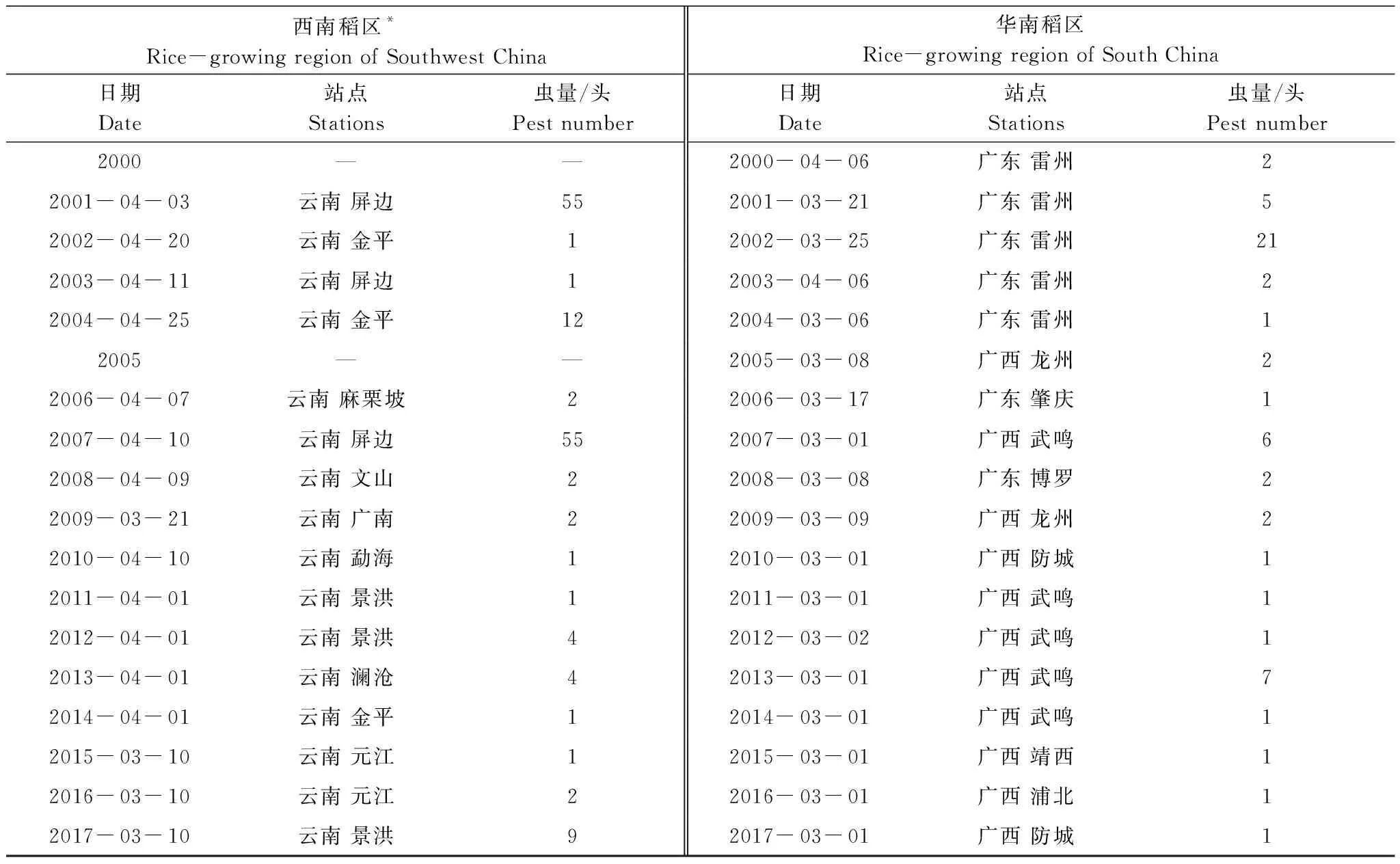
表2 2000—2017年西南、華南稻區的褐飛虱遷入始見期
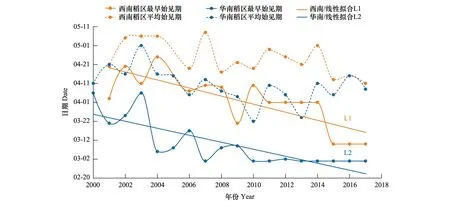
圖2 2000—2017年西南、華南稻區褐飛虱始見期Fig.2 Annual immigration beginning dates of N. lugens in the rice-growing regions of the southwest China and the south China during 2000 to 2017
2.1.2首次遷入峰統計結果
2000—2017年間,境外褐飛虱遷入峰首次出現在西南稻區的日期最早為3月21日,華南稻區最早為3月6日(表3)。在西南稻區內全部站點的平均首遷峰日為5月29日±4.82日,華南稻區全部站點的平均首遷峰日為6月12日±3.96日,結果表明春季褐飛虱首次遷入西南稻區站點的時間在平均水平上反而早于華南稻區。為了排除因兩稻區站點分布差異對結果分析造成的影響,選取兩稻區緯度在22°N—24°N內的所有站點,調查其平均首遷峰日,所得結果相同。這顯示在每年的4—7月褐飛虱種群開始大量遷入我國境內時,若以百頭蟲量為準,則褐飛虱遷入西南稻區的時間更早。其原因一方面,在北半球南亞和東南亞低緯度地區,西南季風爆發早于南海季風,同時,在陸上越靠近印度低壓的地區西南季風越強,就會越早出現有利于褐飛虱北遷的風場[23-24]。因此,從地理經度和緯度上講,西南稻區在褐飛虱初始遷入風場上比華南稻區更有利;另一方面,在每年4—5月不僅從緬甸、泰國、越南等蟲源區到中國的西南、華南稻區的空中溫場條件都能適應于褐飛虱的北遷,且此時正為緬甸旱季稻的黃熟期,為西南稻區提供了大量蟲源[13]。這說明相較于大氣溫度,此時蟲源量大小與是否存在適宜攜蟲的風場已成為影響褐飛虱遷入時間的主要因素。
綜合整個初始遷入期的統計結果來看,褐飛虱初始遷入我國華南稻區的日期整體上要早于西南稻區14 d左右。如圖3中所示,首次遷入峰日有提前趨勢但未形成明顯的時間差異性,主要是年度間的差異(首次遷入峰日早遲與年份負相關但不顯著,L3:r3=-0.698,P<0.01;L4:r4=-0.426,P>0.05)。例如,華南稻區2000和2001年、與2016和2017年幾乎相同;西南稻區2001和2002年與2011和2012年也幾乎相同(表3)。與首遷峰日相比,始見期在不同年份間變動的幅度更小,出現的時間更穩定。
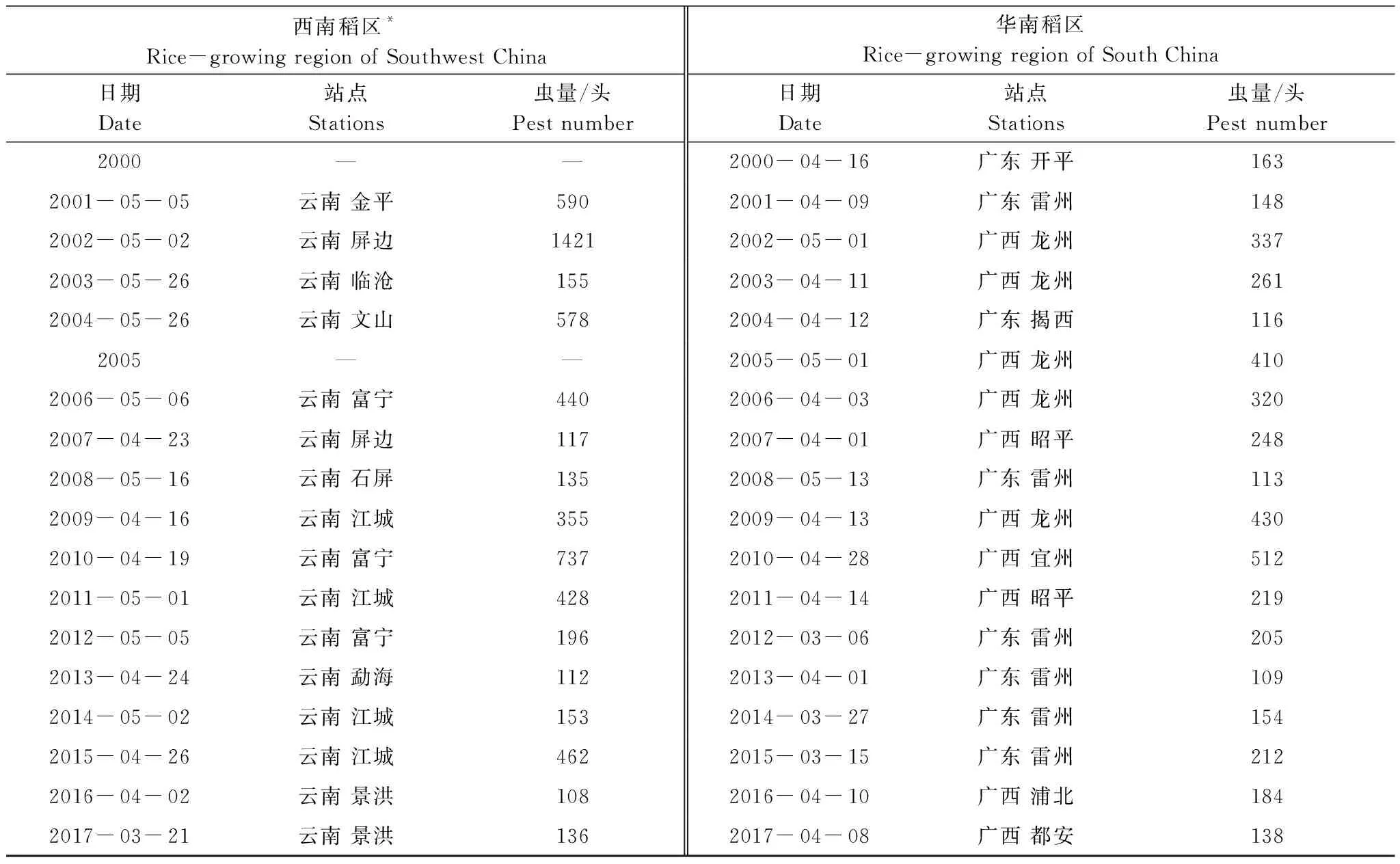
表3 2000—2017年西南、華南稻區的褐飛虱首次遷入峰日
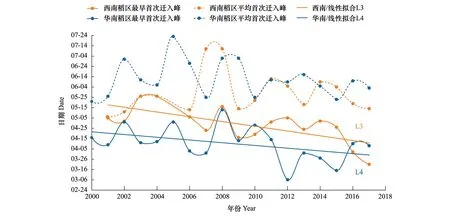
圖3 2000—2017年西南、華南稻區褐飛虱首次遷入峰日Fig.3 First immigration peak dates of N. lugens in the rice-growing regions of the southwest China and the south China during 2000 to 2017
2.2 年內初始遷入種群的蟲源地分布及其遷飛軌跡模擬
為探明多年來年內初始遷入我國褐飛虱種群的境外蟲源分布,本文對18 a中遷入中國西南、華南稻區的初始遷入過程進行了軌跡回推。按不同稻區、不同遷入期將有效回推軌跡疊加到全球地表稻田分布圖上(30 m分辨率)(圖4)。以軌跡走向及蟲源地分布為依據,將各年回推軌跡進行分類統計(表4)。
總結多年回推軌跡的蟲源地,得出年內初始遷入我國西南稻區的蟲源地主要分布在緬甸北部和中東部,華南稻區年內初始遷入蟲源主要來自老撾、越南北部和中國的海南南部。在褐飛虱年內初始遷入西南稻區的過程中,共約有87.5%遷入過程的蟲源來自緬甸北部和中東部稻區,其余少部分蟲源來自于老撾、越南。而在初始遷入華南稻區的過程中,共約有66.7%遷入過程的蟲源來自于老撾和越南的中部或北部稻區,22.2%遷入過程的蟲源來自于緬甸、老撾交界邊境以及海南島。其余少量未歸類回推軌跡的蟲源地分布在雷州半島和廣西南部稻區,為區內的短途遷飛。如圖4,在整個初始遷入過程中回推軌跡多為東—西走向或西南—東北走向,只有個別過程取偏南至偏北走向。此外,褐飛虱在首次遷入峰時期較始見期相比遷入地范圍更大,蟲源地更廣,軌跡聚合度更低。
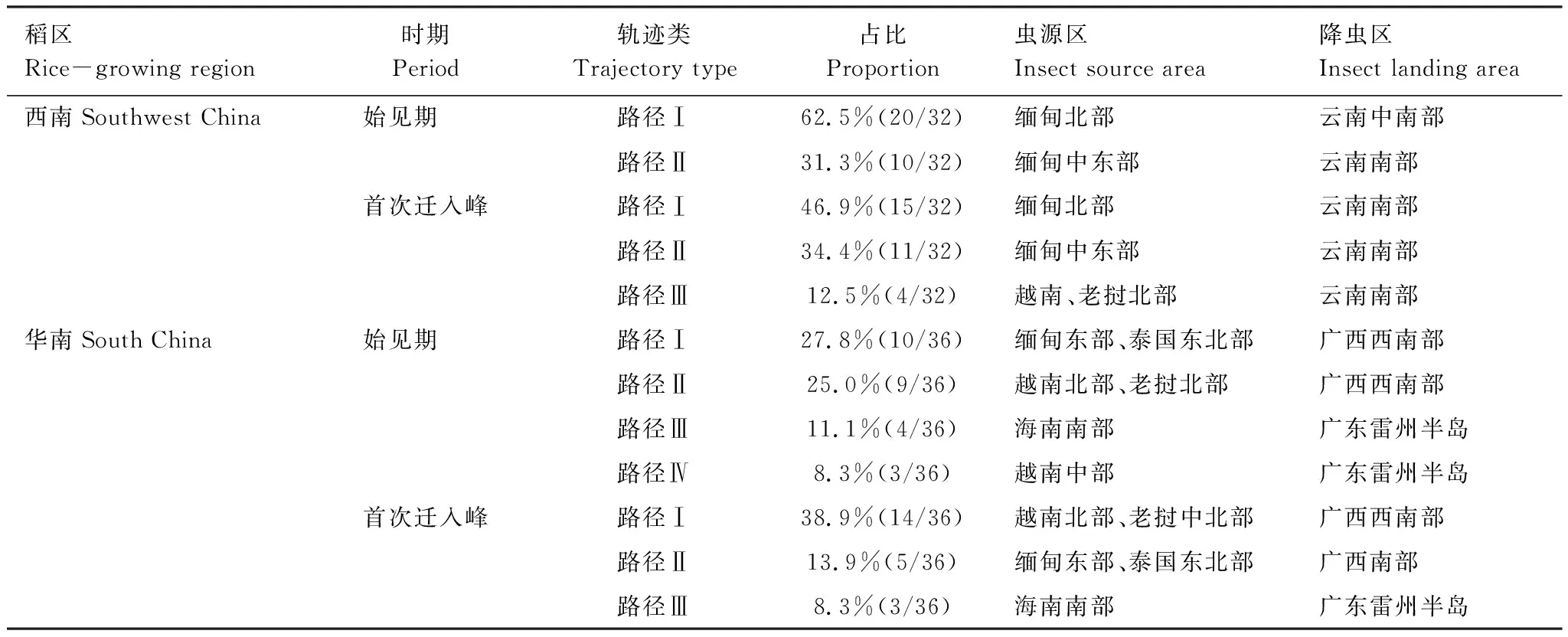
表4 2000—2017年西南、華南稻區初始蟲源地及其軌跡分類
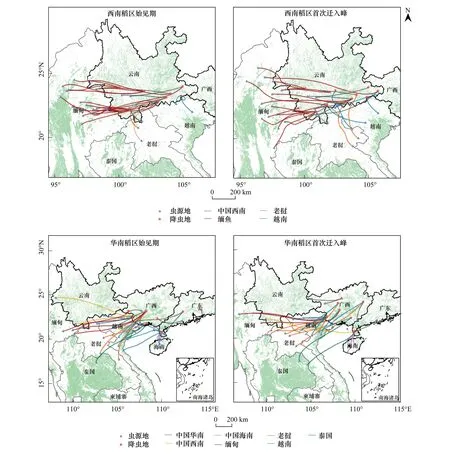
圖4 2000—2017年西南、華南稻區褐飛虱初始蟲源地及其軌跡分布Fig.4 Distribution of source areas and trajectories for N. lugens′ initial immigration in the rice-growing regions of the southwest China and the south China during 2000 to 2017
2.3 低溫脅迫統計特征
使用1.2.3數據統計方法中介紹的方法得到低溫脅迫統計特征值如下表(表5、表6、表7)。為簡明揭示溫度對褐飛虱整個遷飛過程的復雜影響,下面僅對褐飛虱在盛行遷飛層上的脅迫情況進行討論。統計所有受低溫脅迫影響的降落過程發現,在降蟲當晚,降溫過程有21.9%出現在20:00—次日02:00之間,25%出現在02:00—08:00之間,52.5%整晚降溫。在2000—2017年褐飛虱年內初始遷入西南稻區的過程中,約有50.0%的降落過程受到了低溫影響,低溫過程平均強度為13.71℃(表7);在初始遷入華南稻區的過程中,約有58.3%的降落過程受到了低溫影響,低溫過程平均強度為13.18℃。可見,褐飛虱降落在華南稻區受低溫脅迫影響相對更大。相比于華南稻區,西南稻區因緯度跨距大、海拔高度起伏明顯而溫度變化幅度大,但因云南稻區地形北高南低,受地形阻擋,一般冷空氣難以侵入西南南部稻區(即云南中南部稻區),導致褐飛虱種群降落對低溫脅迫的響應概率小;華南稻區因緯度跨距小、海拔高度差異小而溫度變化幅度小,南嶺山系雖呈東西帶狀分布,但其五個山嶺之間是間斷的,且海拔高度明顯低于云南北部山系,對冷空氣的阻力也明顯小于西南南部稻區,導致華南稻區受冷空影響范圍廣、發生概率高,褐飛虱種群降落對低溫脅迫的響應概率和強度都大。綜上,春季西南稻區日均溫高,溫度日較差大,褐飛虱種群降落對低溫脅迫的響應小;華南稻區日均溫低,溫度日較差小,褐飛虱降落對低溫脅迫的響應大。
不同遷入時期的降蟲過程受低溫脅迫影響的差異在兩稻區表現不同,在華南稻區表現出明顯差異,在西南稻區表現為無差異。華南稻區褐飛虱始見期受到低溫脅迫影響的概率(88.9%)遠高于西南稻區始見期(50%)。而在首次遷入峰期中,華南稻區受低溫脅迫影響的概率(27.8%)明顯低于西南稻區(50%)(表7)。對不同遷入時期的低溫脅迫發生情況統計得到,始見期約有70.6%的降落過程受低溫影響,低溫強度為12.53℃,而在首次遷入峰時期僅有38.2%的降落過程受到了低溫影響,低溫強度為14.36℃。統計發現,一般情況下,華南稻區始見期較首次遷入峰日要早2—3個月,始見期日均溫低于16℃的頻率要遠高于首次遷入峰時期,所以在始見期低溫脅迫對降蟲過程產生的影響更顯著。但是,在西南稻區的統計卻顯示:這種低溫脅迫的時間差異性很小,一方面原因可能是北遷過程中地形阻擋因素會使部分褐飛虱遷入種群在遇到低溫之前就遇到了地形動力脅迫而產生了降落,另一方面是因為云南春季氣溫變化幅度小,晝夜溫差大,故在時間維度上西南地區上空大氣溫度低于褐飛虱種群16℃飛行閾值的概率差異小,從而受到低溫脅迫影響降蟲過程的時間差異減小。因此,不同遷入期的降蟲過程受低溫脅迫影響的差異在西南稻區就很小。
綜上,褐飛虱初始遷入我國受低溫脅迫的降落過程在時間和空間分布上均存在差異,特別是在時間分布上的差異更加明顯。就始見期而言,華南稻區的降溫幅度小于西南稻區,但低溫脅迫在華南稻區的影響頻次比西南稻區高、發生時間早、強度大;而在首次遷入峰上,低溫脅迫在西南稻區的影響頻次比華南稻區高、發生時間早、強度大、降溫幅度也大(表7)。
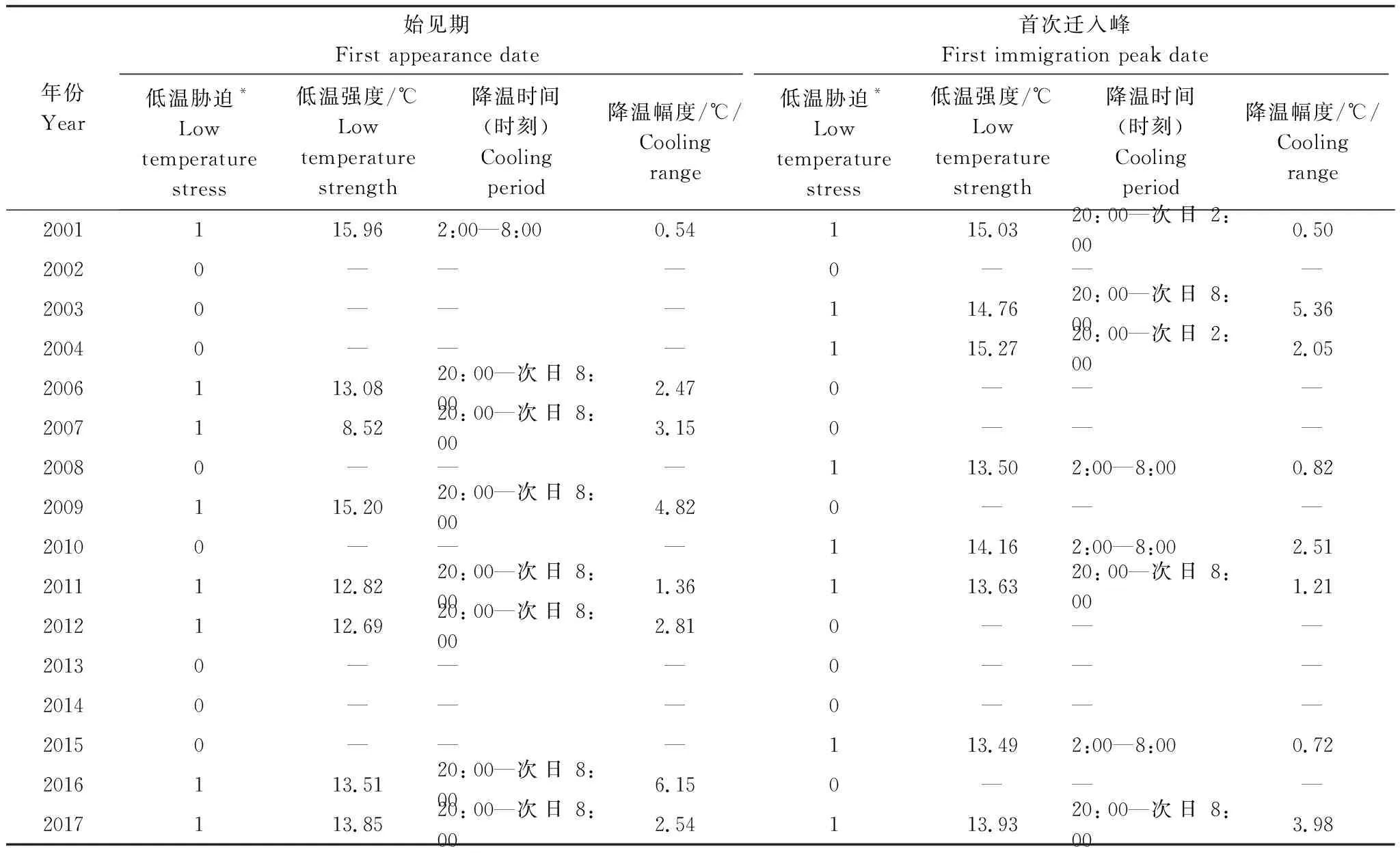
表5 2000—2017年西南稻區逐年800 hPa低溫脅迫特征值

表6 2000—2017年華南稻區逐年850 hPa低溫脅迫特征值
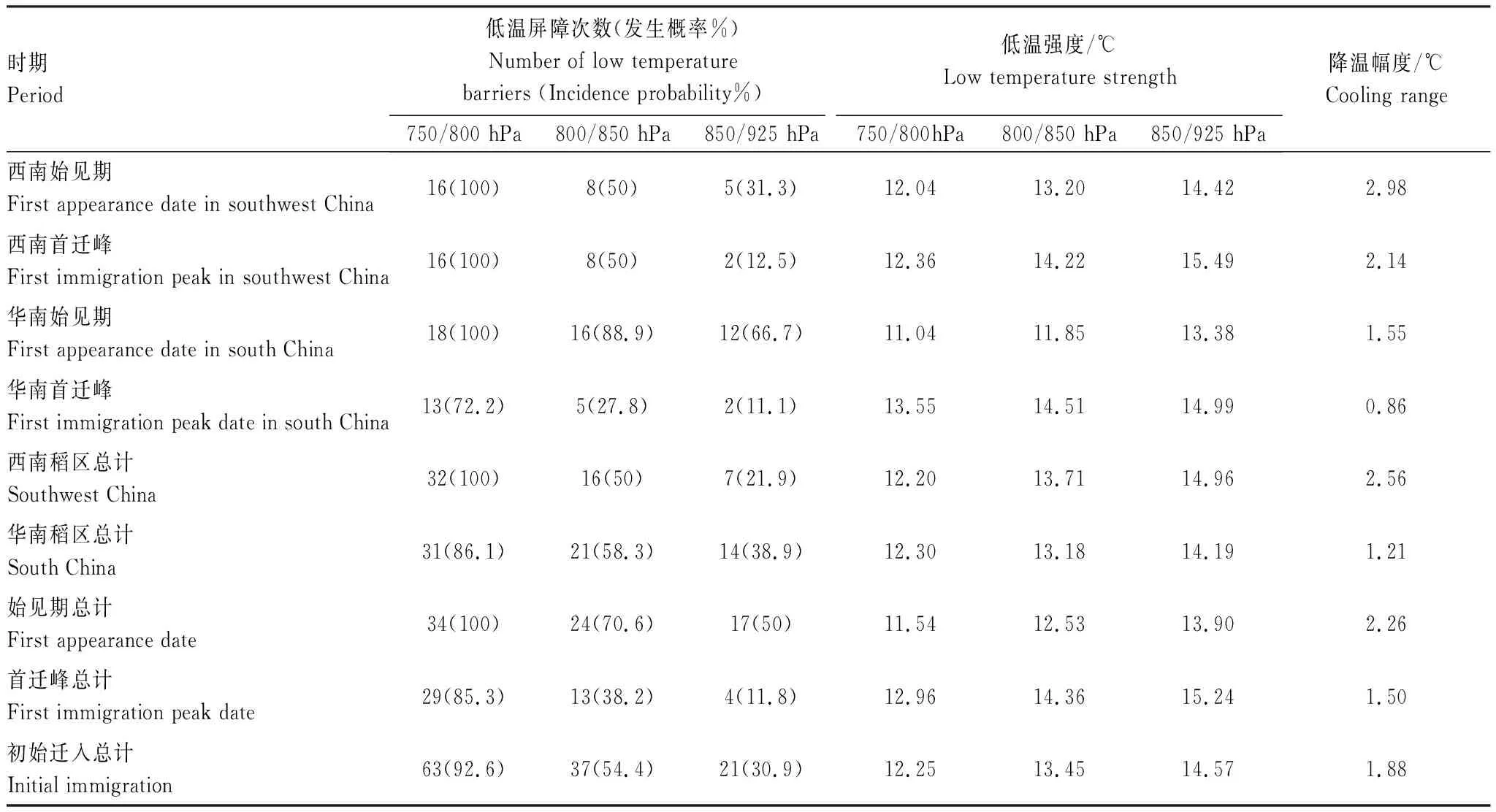
表7 2000—2017年西南、華南稻區不同高度層低溫屏障統計
2.4 遷入地溫度分析
為能更全面地了解褐飛虱遷入與低溫脅迫的關系,本文統計了在整個初始遷入時期降蟲地盛行遷飛層的日均溫,遷入日用黑色橫粗線覆蓋。從圖5可見,西南稻區整個初始遷入期日均溫低于16℃的頻次在整體趨勢上逐年減少,始見期、首次遷入峰逐年提前,這表明西南稻區褐飛虱提早遷入可能與降蟲地盛行遷飛層的溫度升高有關。圖中顯示并非在溫度符合條件后(大于16℃溫度閾值)褐飛虱立即遷入降蟲區,而是遷入時的溫度基本都符合大于16℃的條件(如圖5中2010年)。這說明溫度僅是限制攜蟲氣流遷入某地的一個重要因素,而不是影響初始遷入時期早晚的決定性因素。
為進一步了解始見期出現早遲與近年來降蟲區盛行遷飛層溫度變化間的關系,我們通過調查華南、西南稻區遷入頻率最高的六個站點(西南稻區:景洪JH、金平JP、屏邊PB,華南稻區:雷州LZ、武鳴WM、防城FC)2000—2017年3月份盛行遷飛層的月均溫,首先分析18年來月均溫的年度間變化得出:近年來西南稻區的站點溫度有升高的趨勢(圖6),而華南稻區的站點溫度幾乎沒有變化(圖6)。其次,分析月均溫與始見期出現時間的相關性得出:西南稻區3月份的站點溫度與始見期早遲具有較弱的負相關性,即盛行遷飛層溫度越高,始見期越早。這說明西南稻區褐飛虱始見期提前可能與其盛行遷飛層上大氣溫度升高有關,而華南稻區的始見期提前與其盛行遷飛層上的大氣溫度變化無關。
在華南稻區初始遷入期,特別是始見期(圖5)的遷入地盛行遷飛層溫度低于16℃的頻次逐年增多,初始遷入期卻仍有提前,甚至83.3%遷入日的日均溫都低于16℃,個別年份遠低于16℃(最低溫9.75℃)。但眾所周知,褐飛虱在低于16℃的環境下是無法繼續飛行的。由此推測,華南稻區始見期蟲源的飛行高度一定低于1500 m(850 hPa),同時回推蟲源地與降蟲地的距離相對縮短,故可能為越南北部稻區或海南、華南南部稻區存在的少量越冬蟲源隨偏南風短途低空遷飛后降落本站的,而境內越冬蟲源的活動期提前導致華南稻區褐飛虱燈下始見期提前與近年來氣候變暖使褐飛虱越冬蟲源分布北界北移有關。
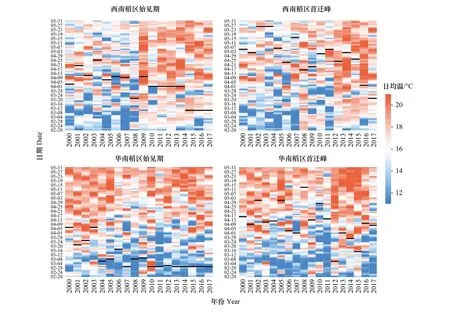
圖5 2000—2017年西南、華南稻區初始遷入期降蟲地盛行遷飛高度層(西南稻區800 hPa/華南稻區850 hPa)日均溫(℃)Fig.5 Daily mean temperature on the prevailing migratory layers (on 800 hPa in the southwest China or 850 hPa in the south China) of N. lugens′ landing areas during the initial immigration dates in the rice-growing regions of the southwest China and the south China during 2000 to 2017

圖6 2000—2017年西南稻區站點(JH景洪、JP金平、PB屏邊),華南稻區站點(LZ雷州、WM武鳴、FC防城)盛行遷飛高度層(西南稻區800 hPa/華南稻區850 hPa)3月份月均溫(℃)與始見期Fig.6 Monthly mean temperature in March of the prevalent migration layers (on 800 hPa in the southwest China or 850 hPa in the south China) of the stations in the rice-growing regions of the southwest China (JH, JP, and PB) and the south China (LZ, WM, and FC), and N. lugens′ immigration beginning dates during 2000 to 2017
3 結論與討論
3.1 褐飛虱初始遷入我國時間提前,華南稻區始見期提早可能與越冬北界的北移有關
近年來,我國褐飛虱年內初始遷入期有所提前。有研究發現在氣候變暖背景下,廣西、云南等地稻飛虱的遷入始見期、首次遷入高峰期均呈逐年提早的趨勢[25-26, 9]。稻飛虱遷入始見期的出現早遲在我國東、西部間存在著差異,在相同緯度上我國東部始見期要早于西部地區[26]。本文統計多年褐飛虱遷入蟲情得到,褐飛虱初始遷入華南稻區時間較西南稻區要早9—14 d。褐飛虱初始遷入我國東、西部稻區上的時間差異不僅與降蟲區的地形、溫濕度條件、水稻種植等情況有關,蟲源地的蟲源情況以及距離降蟲地的遠近也與之密切相關。華南稻區西連越南,南臨南海,盡管地貌類型多樣,但整體海拔較低,與老撾、越南等蟲源地距離近且無阻擋,能為兩廣稻區常年提供穩定蟲源,因此,褐飛虱年內初始遷入期更早。而西南地區地處青藏高原東南側,不僅受到東亞季風和印度季風及中高緯度天氣系統的交叉影響[27],而且西南地區海拔高、地形地勢多變,褐飛虱遷入受地理因素阻礙較大,褐飛虱遷入的降蟲機制相比東部其他省份要更加復雜,多種因素綜合導致褐飛虱年內遷入我國西部始見期要晚于東部地區。
本研究中發現,華南稻區在盛行遷飛層上的大氣溫度無升高的趨勢,但初始遷入期仍然提前了。特別是在華南稻區始見期,約有83.3%褐飛虱遷入過程的降蟲區最適遷飛層的大氣溫度低于16℃,在這種情況下褐飛虱是無法飛行的,但實際情況卻是有蟲遷入。這說明褐飛虱在3月份遷入華南稻區時的遷飛高度遠低于1500 m,同時,近年來研究表明氣候變暖使褐飛虱越冬北界北移,在華南南部稻區存在少量的越冬蟲源[28],在華南北部稻區則存在低空短途遷飛的現象[29]。Hu等[30]在研究2008年廣西北部灣稻區初始遷入的過程中模擬500 m高度下的蟲源,結果顯示低空短途遷入華南的蟲源可能來自于海南島。上述情況可以說明存在少量越冬蟲源在年內早期低空短途遷入華南稻區的可能性。它的遷飛機制為褐飛虱主動起飛升空后,一般如無直達16℃低溫層的強上升氣流,就會逐漸主動適應環境溫度在適溫層內飛行;但若遇到直達16℃以下低溫層的強上升氣流或在隨氣流水平飛行時突遇高空大氣低溫層(即低于16℃的水平氣流層)時,如其不能及時反應并主動調節翅振頻率和飛行高度,并快速進入適溫區,則會致死或驟停振翅,掉下地面。褐飛虱屬小型風載遷飛性昆蟲,其在高空強氣流影響下對適宜溫度層主動選擇的能力有限,而對流層卻有“邊界層頂效應”[31],因此,在空中飛行時,三維空間的氣流速度絕大多數情況下是遠超其主動遷飛的速度的,褐飛虱在這樣的環境流場和溫場條件下是被動的。由此可以推測,這可能是分布在越南北部稻區或海南、華南南部稻區的越冬蟲源在遠低于1500 m的飛行高度下短途遷入華南中部或北部稻區后而導致其他地區測報站在燈下見蟲了。然而,盡管僅有10%左右的概率,仍不排除有蟲源自越南中部跨海遷入的可能性[32]。
3.2 緬甸是西南稻區多年稻飛虱初始遷入過程的穩定蟲源區,越南、老撾、海南是華南稻區多年稻飛虱初始遷入過程的穩定蟲源區。
云南、兩廣是地處我國南端邊陲的省份,也是稻飛虱遷入我國的第一站。關于云南以及兩廣地區的蟲源問題,許多科研工作者已經做出了較為完整的研究。如沈慧梅[15]對2009年云南百背飛虱的蟲源地分析得出云南早期蟲源主要來自于緬甸和金三角地區,而我國華南稻區的早期遷入蟲源則主要來自越南中部。包云軒[13]等對云南省4、5月份早期褐飛虱遷入蟲源地研究發現,褐飛虱遷入的主要蟲源區是緬甸,部分來自老撾和泰國,少量來自越南,還有極少數來自孟加拉國。Hu等[30]等對2008年廣西早期遷入稻飛虱的蟲源進行分析后得出:廣西早期遷入蟲源主要來自越南中部及老撾南部。王政等[33]對廣東白背飛虱多年早期遷入的蟲源地進行分析得出:蟲源主要分布在越南中北部、老撾中南部、海南以及兩廣南部稻區。
本文對西南、華南兩稻區多年褐飛虱遷入始見期、首次遷入峰蟲源地分布進行逆推后發現:多年褐飛虱初始遷入過程的蟲源地分布總是穩定在固定區域內。2000—2017年西南稻區褐飛虱始見期蟲源均來自于緬甸,首次遷入高峰期蟲源區大部分在緬甸,少量分布在老撾和越南。這是由于近年來緬甸政府在農業政策上作了的改革,增加了水稻的種植面積和產量。在每年的4月上中旬,緬甸中北部稻區所種植的水稻處于抽穗、灌漿期,5月初水稻收獲。同時4月份緬甸中北部的高空存在明顯的盛行西風,形成了褐飛虱蟲源遠距離遷飛的輸送通道,為我國云南省褐飛虱的早期遷入提供了蟲源[34]。而華南稻區褐飛虱始見期蟲源主要分布在越南中北部及老撾北部,少量蟲源來自緬甸和海南。越南中北部的紅河三角洲平原以盛產水稻著稱,冬春稻在每年2月份播種,同年4月份水稻抽穗楊花,此時有大量褐飛虱種群開始外遷。同樣由于老撾、越南等地在3、4月份盛行的西南風、西南偏南風為褐飛虱種群遷入我國華南稻區提供了可能[32]。由此可見,華南稻區蟲源地分布更為復雜。
3.3 始見期的低溫脅迫在華南稻區影響頻次比西南稻區高、發生時間早、強度大;首次遷入峰的低溫脅迫在西南稻區的影響頻次比華南稻區高、發生時間早、強度大、降溫幅度也大。
褐飛虱完整的遷飛過程并非像模型模擬的那樣,其中還存在著褐飛虱主動選擇的行為。遷飛的褐飛虱個體在遇到降雨、下沉氣流、風向幅合等外力作用時會集中迫降,或在飛行能源物質耗盡或溫度驟變低于其飛行溫度閾值時也會選擇主動降落[35]。在不考慮食料條件的影響下,一次完整的降落過程是降蟲地地形及多種氣象因素交互作用的結果。盡管降水、下沉氣流是影響褐飛虱大規模降落的兩大主要氣象因子[33, 36],但在初始遷入的過程中,時值早春,大氣溫度低,降雨少,低溫對稻飛虱降落的影響更加明顯[7]。如沈慧梅[15]在對2009年云南白背飛虱降落機制分析中指出,白背飛虱早期遷入云南時,風切變與低溫屏障幾乎存在于每一次降落過程中。本文重點研究了低溫對褐飛虱初始遷入我國的脅迫作用,且初始遷入蟲量較少,故不考慮降水以及下沉氣流對褐飛虱降落的影響。徐蘭珍等[37]在研究滇東南白背飛虱早期遷入過程中得到,影響白背飛虱的降落因子中低溫屏障占56.5%,與本文中得到的西南稻區早期遷入過程中影響褐飛虱的降落因子中低溫屏障占50%相近。本文通過統計兩稻區18 a中共68次初始遷入過程發現,約有54.4%的遷入過程受到了低溫影響,且在不同稻區、不同時期(始見期、首次遷入峰期)的發生情況均有差異。正是因為在空間尺度上不同地理位置獨有的地形特征與氣候特點和時間尺度上氣候變化的特點綜合作用,使華南稻區始見期的遷入過程受低溫影響更大。
盡管本文對有關褐飛虱年內初始遷入過程中低溫脅迫的特征進行了相關的統計分析,但研究中還存在一些不足,如兩稻區2005—2017年為每日燈誘蟲情數據,2000—2004年為每候燈誘蟲情數據且開燈日期較晚(開燈日期:2000- 04- 06,2001- 03- 06,2002- 03- 10,2003- 04- 06,2004- 03- 06,2005—2017年均為3月1日),同時西南稻區缺少2000年和2005年的蟲情數據。這些部分蟲情數據不完整的問題會對本文研究結果產生一定的影響,且本文僅對低溫屏障及其特征進行了初步的定量統計分析,而沒有涉及到低溫是如何在飛行過程中脅迫褐飛虱產生降落行為的以及種群對低溫脅迫的響應是否會在不同時空條件下存在差異,這些機理性問題需要結合個例進行更加詳細深入的闡釋與剖析。
