不同母巖發(fā)育馬尾松土壤固氮菌群落結(jié)構(gòu)和豐度特征
2020-11-13 01:09:58周運(yùn)超
生態(tài)學(xué)報(bào) 2020年17期
趙 輝,周運(yùn)超
1 貴州大學(xué)貴州省森林資源與環(huán)境研究中心/貴州省高原山地林木培育重點(diǎn)實(shí)驗(yàn)室/林學(xué)院,貴陽(yáng) 550025 2 銅仁學(xué)院,銅仁 554300
氮(N)在植物生長(zhǎng)發(fā)育過(guò)程中具有不可替代的作用,它是合成植物體內(nèi)蛋白質(zhì)和核酸的必需元素,同時(shí)其自身也參與植物信號(hào)的傳遞[1]。生物固氮是指將大氣中的N2轉(zhuǎn)化為可被植物吸收利用的含氮化合物的過(guò)程,每年大約有50%的大氣氮通過(guò)該途徑進(jìn)入生態(tài)系統(tǒng)[2]。生物固氮與具有生理功能的需氧和厭氧微生物密切相關(guān)[3]。目前,已經(jīng)發(fā)現(xiàn)60多個(gè)屬的固氮微生物,主要包括固氮菌屬(Aztobacer)、固氮螺菌屬(Azospirillum)、假單孢菌屬(Pseudomonas)、克雷伯氏菌屬(Klebsiella)和脫硫弧菌屬(Desulfovibrio)等[4]。固氮菌數(shù)量和群落結(jié)構(gòu)對(duì)土壤固氮速率和氮循環(huán)有直接的影響,可作為衡量土壤質(zhì)量的重要指標(biāo)[5]。生物固氮是由固氮酶催化,而固氮酶是一種能夠?qū)⒎肿拥€原成氨的酶,有兩個(gè)多亞基的金屬蛋白酶組成,分別為nifD,nifK基因編碼的鉬鐵蛋白和nifH基因編碼的鐵蛋白[6]。其中,nifH基因是系統(tǒng)進(jìn)化過(guò)程最古老的功能基因之一,在進(jìn)化過(guò)程中與16SrDNA保持一致[7]。因此nifH作為生物標(biāo)記基因被廣泛用于研究固氮微生物群落結(jié)構(gòu)[8]。
馬尾松(PinusmassonianaLamb.)是我國(guó)特有的針葉樹(shù)種之一,由于其適應(yīng)性強(qiáng)和耐干旱瘠薄等特點(diǎn),廣泛分布于我國(guó)南方地區(qū),在我國(guó)生態(tài)環(huán)境建設(shè)和林業(yè)產(chǎn)業(yè)發(fā)展中發(fā)揮著重要作用[9]。因馬尾松屬于速生林,在生長(zhǎng)過(guò)程中對(duì)土壤氮素的需求量較大。而目前在馬尾松生長(zhǎng)的土壤環(huán)境中,有限的氮素利用率成為限制其林木速成豐產(chǎn)的關(guān)鍵因素[10]。對(duì)于很少有氮素施入的馬尾松林而言,生物固氮是維持馬尾松林生長(zhǎng)的主要氮源,對(duì)馬尾松人工林速生豐產(chǎn)及地力維持具有重要作用。然而,由于馬尾松人工林主產(chǎn)區(qū)土壤母巖類型多樣,并且在造林過(guò)程中沒(méi)有做到因地經(jīng)營(yíng),因而導(dǎo)致大量低產(chǎn)林存在。因此,通過(guò)對(duì)不同母巖發(fā)育馬尾松土壤固氮菌群落結(jié)構(gòu)和豐度及其影響因素進(jìn)行研究,對(duì)了解母巖發(fā)育馬尾松土壤生物固氮及調(diào)節(jié)土壤氮素供應(yīng)具有極其重要的意義。
土壤母質(zhì)(巖)是巖石及礦物的風(fēng)化產(chǎn)物,決定著土壤的形成及發(fā)育,而不同母巖發(fā)育土壤自身養(yǎng)分的差異會(huì)引起土壤微生物群落結(jié)構(gòu)的變化[11],并對(duì)植物生長(zhǎng)發(fā)育有重要影響。相關(guān)研究認(rèn)為,土壤固氮微生物群落受到非生物和生物因素影響,非生物因子有土壤pH[12]、有效性氮[13]、土壤質(zhì)地[14]和土壤類型等[15];生物因子有植被類型等[16],這些因素造成固氮微生物群落結(jié)構(gòu)和多樣性在不同母巖和植被下差異較大。目前,不同母巖發(fā)育土壤馬尾松人工林的研究主要集中在林木生長(zhǎng)[17]、酶活性[18]和土壤養(yǎng)分[19]等方面。關(guān)于亞熱帶地區(qū)不同母巖發(fā)育馬尾松土壤固氮菌群落結(jié)構(gòu)、多樣性和豐度的研究未見(jiàn)報(bào)道。為此,本文運(yùn)用Illumina MiSeq高通量測(cè)序技術(shù)和熒光定量PCR技術(shù)對(duì)亞熱帶地區(qū)主要母巖發(fā)育馬尾松土壤固氮菌群落結(jié)構(gòu)及豐度進(jìn)行了研究,探討土壤固氮菌群落結(jié)構(gòu)和豐度在不同母巖發(fā)育土壤中的分布特征及其與土壤環(huán)境因子的關(guān)系,以揭示不同母巖發(fā)育土壤固氮能力及固氮菌生態(tài)功能,從固氮微生物角度來(lái)分析不同母巖土壤質(zhì)量狀況,為馬尾松人工林適地造林及速生豐產(chǎn)提供科學(xué)依據(jù)。
1 材料與方法
1.1 研究區(qū)概況及試驗(yàn)設(shè)計(jì)
研究區(qū)位于貴州大學(xué)造林生態(tài)園(106°40′42″E,26°25′41″N),海拔1100 m,屬于典型中亞熱帶季風(fēng)氣候,年相對(duì)濕度81%,年平均氣溫14.9℃,無(wú)霜期285 d左右,年平均日照總數(shù)為1274.2 h。
為盡量減少氣候和群落植被等外界環(huán)境因子差異對(duì)試驗(yàn)造成的影響,采取了池栽試驗(yàn)。選取貴州省馬尾松造林面積分布較廣泛的土體為研究對(duì)象,主要發(fā)育于四類母巖,分別為變余砂巖(Blastopsammite)、長(zhǎng)石石英砂巖(Feldspathic quartz sandstone)、石英砂巖(Quartz sandstone)和玄武巖(Basalt)。四類母巖發(fā)育形成的土壤過(guò)1 cm篩后,分別裝入長(zhǎng)×寬×深=5.0 m×1.2 m×0.5 m石框內(nèi)作為1個(gè)試驗(yàn)小區(qū),每類母巖土壤重復(fù)3次,共計(jì)12個(gè)試驗(yàn)小區(qū),每小區(qū)面積為6 m2。土壤基本情況見(jiàn)表1。于2014年4月向小區(qū)內(nèi)移栽生長(zhǎng)發(fā)育良好且大小均一的1年生馬尾松實(shí)生苗木,每小區(qū)36株。試驗(yàn)期間管理方式一致,均未施肥。

表1 不同土壤母巖土壤基本情況
1.2 樣品采集
2017年7月進(jìn)行土壤樣品采集。在每個(gè)試驗(yàn)小區(qū)內(nèi)沿S形樣線用直徑5 cm土鉆均勻采取5個(gè)0—10 cm土壤樣品,混合成1個(gè)土樣,用低溫冰盒保存并迅速帶回實(shí)驗(yàn)室。土樣在室內(nèi)去除石塊和植物根系等雜物并過(guò)2 mm篩后,最終分為三份,一份保存于-80℃冰箱,用于nifH基因群落結(jié)構(gòu)和豐度分析;一份4℃冰箱保存,用于土壤微生物生物量碳和氮測(cè)定;一份用于土壤銨態(tài)氮和硝態(tài)氮測(cè)定,余下部分室內(nèi)風(fēng)干,過(guò)篩和研磨用于土壤化學(xué)指標(biāo)測(cè)定。
1.3 測(cè)定方法
1.3.1馬尾松株高和地徑測(cè)定
2017年7月對(duì)馬尾松株高和地徑進(jìn)行測(cè)定。株高采用鋼卷尺測(cè)定,地徑采用游標(biāo)卡尺測(cè)定。
1.3.2土壤化學(xué)性質(zhì)及微生物量碳氮測(cè)定
土壤化學(xué)性質(zhì)采用魯如坤方法測(cè)定[20]:土壤pH采用電位法;有機(jī)碳采用重鉻酸鉀氧化法;全氮采用凱氏定氮法;堿解氮采用堿解擴(kuò)散法;銨態(tài)氮采用靛酚藍(lán)比色法;硝態(tài)氮采用酚二磺酸比色法;速效磷采用碳酸氫鈉浸提-鉬銻抗比色法;速效鉀采用乙酸銨提取-火焰光度法。土壤微生物量碳和氮采用吳金水方法測(cè)定[21]:微生物量碳采用氯仿熏蒸-容量分析法,微生物量氮采取氯仿熏蒸-茚三酮比色法。
1.3.3土壤DNA提取及高通量測(cè)定
稱取0.5 g土壤,按照E.Z.N.A.? Soil DNAKit(Omega,GA,USA)試劑盒操作步驟提取土壤DNA。用1%的瓊脂糖凝膠電泳檢測(cè)DNA完整性,用核酸定量?jī)x(Nanodrop-NC2000)檢測(cè)DNA濃度和純度。
采用引物nifH-F(5′-AAAGGYGGWATCGGYAARTCCACCAC- 3′)與nifH-R(5′-TTGTTSGCSGCRTACATS GCCATCAT- 3′)擴(kuò)增固氮菌nifH基因[22]。PCR體系(25 μL):5×ExTaq緩沖液5.0 μL,dNPT(2.5 μmol/L)2.0 μL,上下游引物(10 μmol/L)各1.0 μL,DNA模板2.0 μL(1—10 ng),ExTaq(5 U/μL)0.25 μL,最后用超純水(ddH2O)補(bǔ)至25 μL。PCR反應(yīng)條件為:95℃ 3 min;94℃ 30 s,55℃ 30 s,72℃ 30 s,30個(gè)循環(huán);72℃ 5 min。以2%瓊脂糖凝膠電泳檢測(cè)PCR產(chǎn)物。樣品送至上海派森諾測(cè)序公司(Personalbio,China),運(yùn)用Illumina MiSeq測(cè)序平臺(tái)進(jìn)行測(cè)序。
1.3.4nifH基因豐度測(cè)定
利用熒光定量PCR(Real-time PCR)技術(shù)檢測(cè)nifH基因拷貝數(shù),反應(yīng)在ABI7500熒光定量PCR儀(ABI,CA,USA)上進(jìn)行,反應(yīng)引物同1.3.3所示。反應(yīng)體系為:2×SYBR real-time PCR premixture(Bioteke,Beijing)10 μL,上下游引物(10 μmol/L)各0.4 μL,DNA模板1.0 μL(1—10 ng),最后用超純水(ddH2O)補(bǔ)至20 μL。熒光定量PCR反應(yīng)條件為:95℃ 5 min;95℃ 15 s,60℃ 30 s,40個(gè)循環(huán)。按照Poly方法[23],獲得含有nifH基因的重組質(zhì)粒,分別以10倍梯度稀釋基因重組質(zhì)粒得到標(biāo)準(zhǔn)曲線。并根據(jù)標(biāo)準(zhǔn)曲線計(jì)算基因豐度,nifH豐度最終被計(jì)算為每克干土的拷貝數(shù)。
1.4 數(shù)據(jù)分析
利用FLASH軟件對(duì)通過(guò)質(zhì)量初篩的雙端序列根據(jù)重疊堿基進(jìn)行配對(duì)連接,從而獲得每個(gè)樣本的有效序列。通過(guò)QIIME軟件調(diào)用USEARCH檢查并剔除嵌合體序列。將優(yōu)質(zhì)序列聚類成操作分類單元(Operational Taxonomic Unit,OTU),閾值設(shè)置為97%,并選取每個(gè)OTU中豐度最高的序列作為該OTU的代表序列。應(yīng)用RDP-classifier在GenBank中對(duì)OTU進(jìn)行分類注釋,統(tǒng)計(jì)各樣品門(mén)和屬水平上群落組成;使用QIIME軟件,對(duì)Chao1、ACE、Shannon和Simpson指數(shù)進(jìn)行計(jì)算。
采用SPSS21.0統(tǒng)計(jì)軟件,通過(guò)單因素方差分析(one-way ANOVA,n=3,P<0.05)和多重比較檢驗(yàn)四類母巖發(fā)育土壤化學(xué)性質(zhì)、馬尾松株高、地徑、固氮菌α多樣性指數(shù)、豐度和群落組成相對(duì)豐度的差異顯著性(P<0.05);相關(guān)性分析用SPSS21.0和R軟件完成;固氮菌群落結(jié)構(gòu)聚類分析、非度量多維尺度分析(Non-metric multidimensional scaling,NMDS)和冗余分析(Redundancy analysis,RDA)用R軟件完成。
2 結(jié)果與分析
2.1 馬尾松株高和地徑
四類母巖馬尾松株高和地徑之間差異顯著(P<0.05)(圖1)。BL、FQS、QS和BA發(fā)育土壤下的馬尾松株高(地徑)分別為282.30 cm(2.78 cm)、292.93 cm(2.84 cm)、305.17 cm(3.05 cm)和340.83 cm(3.36 cm),其中BA土壤下馬尾松株高和地徑顯著高于BL和FQS(P<0.05)。由此可知,BA發(fā)育的土壤有利于馬尾松生長(zhǎng)。
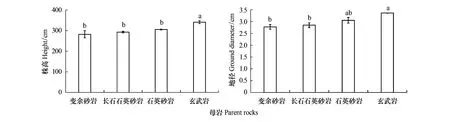
圖1 不同母巖發(fā)育土壤馬尾松株高和地徑Fig.1 Height and ground diameter of Pinus massoniana soils developed from different parent rocksBL:變余砂巖 Blastopsammite;FQS:長(zhǎng)石石英砂巖 Feldspathic quartz sandstone;QS:石英砂巖 Quartz sandstone;BA:玄武巖 Basalt
2.2 土壤化學(xué)性質(zhì)和微生物量碳氮
不同母巖馬尾松土壤化學(xué)性質(zhì)如表2所示。四類母巖發(fā)育馬尾松土壤均呈弱酸性(pH<6.5),QS土壤pH最低為4.71,顯著低于其他母巖(P<0.05)。土壤有機(jī)碳、全氮和堿解氮含量均是BA最高,分別為28.34 g/kg、1.32 g/kg和168.54 mg/kg,并與其他母巖差異顯著(P<0.05)。BA土壤銨態(tài)氮含量最低,硝態(tài)氮含量最高,分別與其他母巖差異不顯著。BL土壤速效磷含量最高,速效鉀含量最低。
不同母巖土壤微生物量碳和氮差異顯著(P<0.05)(圖2),且微生物量碳含量高于微生物量氮。BL土壤微生物量碳含量顯著高于其他母巖(P<0.05),BA土壤微生物量碳含量最低,為147.13 mg/kg,但其土壤微生物量氮含量顯著高于其他母巖(P<0.05)。由此可知,不同母巖發(fā)育馬尾松土壤微生物量碳和氮呈現(xiàn)出不同的變化趨勢(shì)。

表2 不同母巖發(fā)育馬尾松土壤化學(xué)性質(zhì)
2.3 固氮微生物nifH基因豐度
不同母巖馬尾松土壤固氮微生物nifH基因豐度差異顯著(P<0.05)。由圖3可見(jiàn),四類母巖土壤nifH基因豐度范圍為0.19×105—1.5×105copies/g干土。BA土壤nifH基因豐度顯著高于其他三類母巖(P<0.05),而B(niǎo)L、FQS和QS之間差異不顯著(P>0.05),大小順序?yàn)锽A>BL>QS>FQS。

圖2 不同母巖發(fā)育馬尾松土壤微生物量碳和氮 Fig.2 Microbial biomass carbon and nitrogen of Pinus massoniana soils developed from different parent rocks不同小寫(xiě)字母表示不同處理間差異達(dá)到顯著水平(P<0.05)

圖3 不同母巖發(fā)育馬尾松土壤樣品nifH基因豐度 Fig.3 Abundance of nifH gene in Pinus massoniana soils developed from different parent rocks不同小寫(xiě)字母表示不同處理間差異達(dá)到顯著水平(P<0.05)
為明確不同母巖土壤固氮微生物nifH基因豐度差異的影響因素,由nifH基因豐度與土壤化學(xué)性質(zhì)、馬尾松株高和地徑進(jìn)行相關(guān)性分析。結(jié)果表明(圖4),nifH基因豐度分別與土壤有機(jī)碳、全氮、堿解氮、微生物量氮含量和株高之間呈極顯著正相關(guān)關(guān)系(P<0.01),與地徑呈顯著正相關(guān)關(guān)系(P<0.05)。可知,固氮菌數(shù)量與土壤氮素供應(yīng)和馬尾松生長(zhǎng)之間關(guān)系密切。

圖4 群落多樣性指數(shù)、nifH基因豐度與土壤化學(xué)性質(zhì)、株高和地徑相關(guān)性Fig.4 Correlation between community diversity index,nifH gene abundance and soil chemical properties,Height,ground diameter*和**分別表示相關(guān)性顯著在P<0.05和P<0.01水平。SMBC:土壤微生物量碳 Soil microbial biomass carbon;SMBN:土壤微生物量氮 Soil microbial biomass nitrogen;H:株高 Height;Gd:地徑 Ground diameter;nifH:nifH基因豐度 Abundance of nifH gene;Chao1:Chao1指數(shù) Chao1 index;ACE:ACE指數(shù) ACE index;Simpson:辛普森指數(shù) Simpson index;Shannon:香農(nóng)指數(shù) Shannon index
2.4 固氮微生物測(cè)序結(jié)果和α多樣性
利用Illumina MiSeq平臺(tái)對(duì)固氮微生物nifH基因測(cè)序分析,BL、FQS、QS和BA土壤中獲得質(zhì)控后序列數(shù)分別為41338、34229、38223和45441條(表3)。把相似度水平≥97%的序列聚為一個(gè)操作分類單元(OTU),其中BA土壤含有的OTUs數(shù)最多(2206個(gè)),FQS土壤OTUs數(shù)最少(1471個(gè))。由單因素方差分析可知(表3),不同母巖發(fā)育馬尾松土壤α多樣性指數(shù)差異顯著(P<0.05)。四類母巖土壤α多樣性指數(shù)變化趨勢(shì)一致,大小順序均為BA>BL>QS>FQS。說(shuō)明BA發(fā)育馬尾松土壤固氮菌多樣性相對(duì)更為豐富。
相關(guān)性分析結(jié)果表明(圖4),Chao1指數(shù)和ACE指數(shù)與土壤全氮呈顯著正相關(guān)關(guān)系(P<0.05)。Simpson指數(shù)與土壤有機(jī)碳(r=0.651,P<0.05)和堿解氮(r=0.688,P<0.05)均呈顯著正相關(guān)關(guān)系;與全氮(r=0.709,P<0.01)和微生物量氮(r=0.750,P<0.01)呈極顯著正相關(guān)關(guān)系。Shannon指數(shù)與土壤有機(jī)碳(r=0.791,P<0.01)、全氮(r=0.864,P<0.01)、堿解氮(r=0.814,P<0.01)和微生物量氮(r=0.808,P<0.01)均呈極顯著正相關(guān)關(guān)系,與株高(r=0.594,P<0.05)和地徑(r=0.628,P<0.05)呈顯著正相關(guān)關(guān)系。

表3 不同母巖發(fā)育馬尾松土壤固氮菌α多樣性
2.5 固氮微生物群落組成
通過(guò)對(duì)樣品獲得的OTUs進(jìn)行歸類,得到6個(gè)門(mén)、14個(gè)綱、41個(gè)目、69個(gè)科和122個(gè)屬。在門(mén)水平上,分別為變形菌門(mén)(Proteobacteria)、藍(lán)藻門(mén)(Cyanobacteria)、厚壁菌門(mén)(Firmicutes)、放線菌門(mén)(Actinobacteria)、疣微菌門(mén)(Verrucomicrobia)和浮霉菌門(mén)(Planctomycetes)(圖5)。變形菌門(mén)和藍(lán)藻門(mén)為主要優(yōu)勢(shì)類群,相對(duì)豐度分別為84.05%—94.71%和0.57%—3.94%。藍(lán)藻門(mén)和疣微菌門(mén)在四類母巖發(fā)育馬尾松土壤之間差異極顯著(P<0.01),其中藍(lán)藻門(mén)相對(duì)豐度在BL土壤最高。變形菌門(mén)、厚壁菌門(mén)和放線菌門(mén)在四類母巖土壤之間差異顯著(P<0.05)。變形菌門(mén)相對(duì)豐度在BA土壤顯著高于其他母巖(P<0.05);厚壁菌門(mén)相對(duì)豐度在FQS土壤最高(1.19%),QS土壤最低(0.02%);放線菌門(mén)相對(duì)豐度在QS土壤最高(0.17%),FQS土壤最低(0.04%);疣微菌門(mén)在QS土壤不存在;浮霉菌門(mén)僅在QS土壤存在,相對(duì)豐度為0.02%。

圖5 不同母巖發(fā)育馬尾松土壤固氮菌門(mén)和屬水平組成Fig.5 The nitrogen-fixing microbial community composition at the phylum and genus levels in Pinus massoniana soils developed from different parent rocks*P<0.05,**P<0.01
在屬水平上,將平均相對(duì)豐度<0.3%類群歸類為其他,得到20個(gè)類群(圖5)。其中,慢生根瘤菌屬(Bradyrhizobium)、眉藻屬(Calothrix)、根瘤菌屬(Rhizobium)和固氮螺菌屬(Azospirillum)為主要優(yōu)勢(shì)類群,相對(duì)豐度分別為65.13%—77.78%、0.26%—11.24%、1.14%—7.72%和1.71%—4.14%。慢生根瘤菌屬在四類母巖發(fā)育土壤之間差異顯著(P<0.05)。眉藻屬、根瘤菌屬和固氮螺菌屬在四類母巖發(fā)育土壤之間差異極顯著(P<0.01)。慢生根瘤菌屬和固氮螺菌屬相對(duì)豐度均在BA土壤最高;眉藻屬相對(duì)豐度在QS土壤最高,BA土壤最低;根瘤菌屬相對(duì)豐度在BL土壤最高,BA土壤最低。
2.6 固氮菌群落結(jié)構(gòu)及其與土壤化學(xué)性質(zhì)的關(guān)系
基于OTU層次聚類分析結(jié)果顯示(圖6),四類母巖發(fā)育馬尾松土壤固氮菌群落結(jié)構(gòu)差異明顯,相同處理的3個(gè)重復(fù)都聚類在一起。NMDS分析進(jìn)一步證實(shí)了四類母巖發(fā)育馬尾松土壤固氮菌群落結(jié)構(gòu)差異明顯(圖6)。四類母巖土壤固氮菌群落結(jié)構(gòu)在NMDS1和NMDS2發(fā)生明顯分離,分布在不同象限內(nèi)。其中,BA與FQS、BL和QS距離較遠(yuǎn),說(shuō)明BA土壤固氮菌群落結(jié)構(gòu)與其他母巖差異較大;FQS和QS相距較近,說(shuō)明這二類母巖發(fā)育土壤固氮菌群落結(jié)構(gòu)相似度較高。

圖6 不同母巖土壤固氮菌群落結(jié)構(gòu)的層次聚類樹(shù)和NMDS分析Fig.6 Hierarchical clustering tree and nonmetric multidimensional scaling of the nitrogen-fixing microbial community structure in Pinus massoniana soils developed from different parent rocks
為進(jìn)一步分析土壤化學(xué)性質(zhì)對(duì)固氮菌群落結(jié)構(gòu)的影響,對(duì)固氮菌群落結(jié)構(gòu)與土壤化學(xué)性質(zhì)進(jìn)行冗余分析(圖7)。結(jié)果表明,第1軸(A x i s 1)和第2軸(A x i s 2)分別解釋了46.99%和30.56%的變異,兩者累計(jì)解釋變異量為77.55%。由此可知,前兩軸能夠很好地反映土壤固氮菌群落組成與土壤化學(xué)成分之間的關(guān)系。土壤有機(jī)碳、全氮、堿解氮和微生物量氮對(duì)固氮菌群落有極顯著影響。
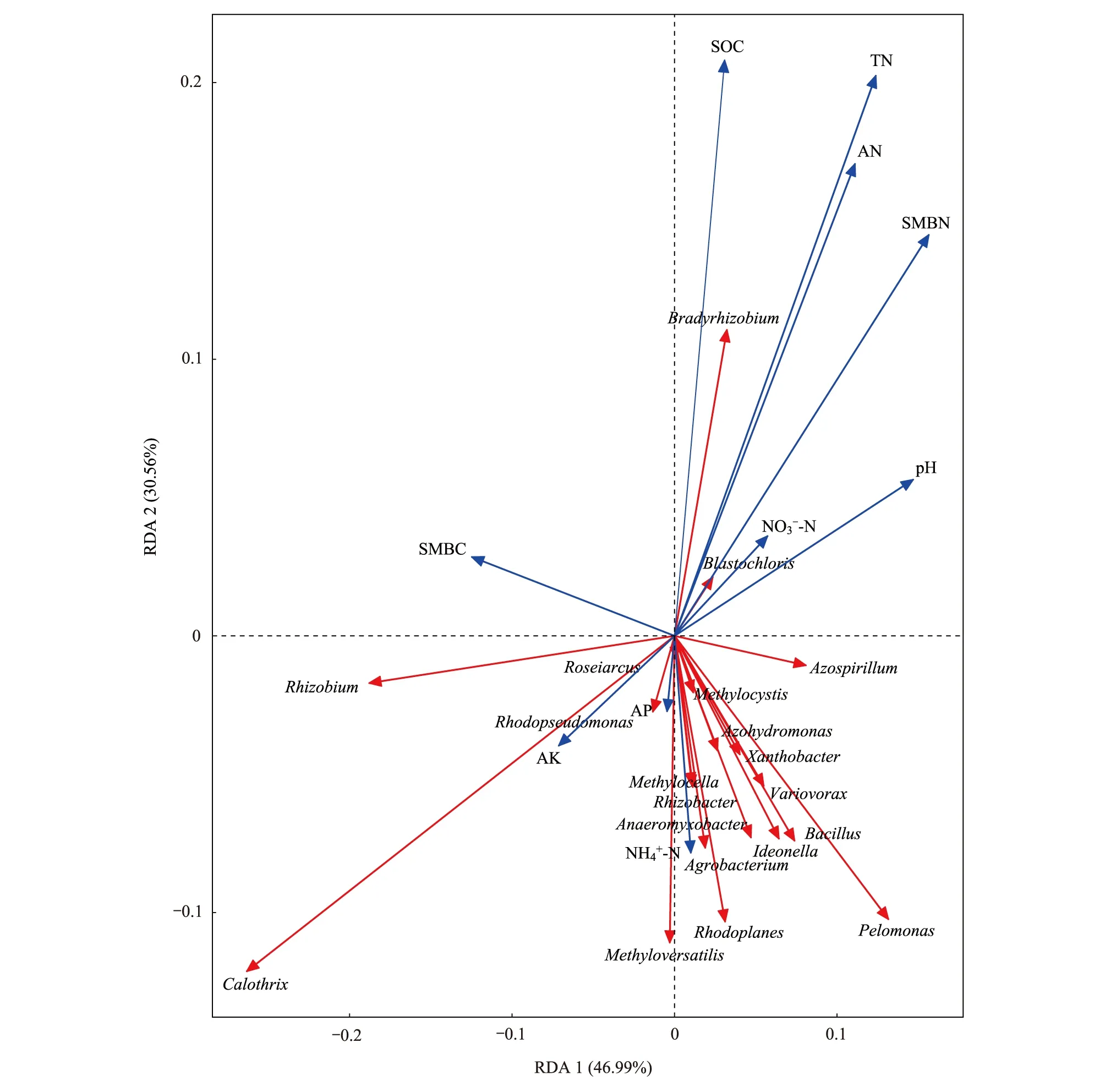
圖7 屬水平固氮菌群落和土壤化學(xué)性質(zhì)的冗余分析Fig.7 Redundancy analysis between the nitrogen-fixing microbial community structureat the genus level and soil chemical properties
3 討論
3.1 不同母巖發(fā)育馬尾松土壤對(duì)固氮菌豐度和多樣性的影響
土壤能夠?yàn)槲⑸锷L(zhǎng)提供所需養(yǎng)分,土壤養(yǎng)分的差異直接導(dǎo)致微生物數(shù)量和群落結(jié)構(gòu)發(fā)生變化[24]。閆小莉和王德?tīng)t[25]研究認(rèn)為,土壤母巖對(duì)其發(fā)育土壤理化性質(zhì)有顯著影響。因此,母巖也將間接對(duì)土壤微生物種群數(shù)量和結(jié)構(gòu)產(chǎn)生重要影響。本研究中四類母巖發(fā)育馬尾松土壤固氮菌豐度與α多樣性指數(shù)變化規(guī)律相同,均是玄武巖土壤顯著高于其他母巖。大量研究表明,土壤有機(jī)碳[26]、氮[27- 28]和根系分泌物[29]是影響nifH基因豐度的主要因素。Herbert[30]研究認(rèn)為,微生物固氮所需能量主要來(lái)源于土壤有機(jī)碳,每固定一分子氮將需要16個(gè)ATP。Coelho等[27]研究認(rèn)為,土壤氮含量增加將導(dǎo)致nifH基因豐度降低;然而Juraeva等[28]研究認(rèn)為,nifH基因豐度隨土壤氮含量增加而增加。楊亞?wèn)|等[29]研究認(rèn)為,植物生長(zhǎng)越旺盛,根系分泌物越多,越有利于固氮菌生長(zhǎng)。Nelson和Mele[31]研究表明,土壤質(zhì)地對(duì)固氮菌數(shù)量影響較大,黏質(zhì)土壤比沙質(zhì)土壤具有更多的nifH基因豐度,與本研究結(jié)果相似。本研究中玄武巖土壤nifH基因豐度顯著高于其他母巖,這可能由于玄武巖土壤的有機(jī)碳和氮養(yǎng)分含量高,馬尾松生長(zhǎng)快,根系分泌物代謝物較多,為固氮菌生長(zhǎng)提供較多的碳源和氮源;而變余砂巖、石英砂巖和長(zhǎng)石石英砂巖土壤養(yǎng)分含量低,不能為固氮菌生長(zhǎng)提供充足的碳源和養(yǎng)分,因此造成固氮微生物數(shù)量較少。Huang等[32]研究認(rèn)為,植物會(huì)對(duì)土壤理化性質(zhì)產(chǎn)生直接或間接的影響。與栽種前相比,本研究中四類母巖馬尾松生長(zhǎng)的土壤pH均有所下降,這可能由于馬尾松生長(zhǎng)過(guò)程中根系分泌物或針葉在分解過(guò)程中產(chǎn)生酸性物質(zhì)所致。Silva等[33]研究認(rèn)為,土壤pH與nifH基因豐度有顯著相關(guān)性。然而,本研究中土壤pH與nifH基因豐度沒(méi)有顯著相關(guān)性,與楊亞?wèn)|等[29]研究結(jié)果一致;但是土壤pH在不同母巖發(fā)育土壤之間差異顯著,因此土壤pH可能通過(guò)改變土壤理化性質(zhì)[34],間接對(duì)固氮菌數(shù)量產(chǎn)生一定影響。
微生物α多樣性指數(shù)是評(píng)價(jià)土壤微生物群落多樣性的重要指標(biāo),多樣性指數(shù)越高表明微生物群落豐富度和多樣性越高,土壤肥力越高,土壤生態(tài)環(huán)境越穩(wěn)定[35]。Chao1或ACE指數(shù)越大,表明群落豐富度越高。Simpson和Shannon指數(shù)越高表明群落多樣性越高。周燕等[36]研究認(rèn)為,固氮微生物多樣性與植物凋落物和根系分泌物關(guān)系密切。本研究表明,玄武巖土壤固氮菌α多樣性指數(shù)均顯著高于其他母巖,表明玄武巖土壤固氮菌的豐富度和多樣性較高,可能由于馬尾松生長(zhǎng)較快,凋落物和根系分泌物增多,導(dǎo)致養(yǎng)分歸還和轉(zhuǎn)化速率加快[37],土壤可利用養(yǎng)分含量增加,有利于固氮微生物生長(zhǎng)繁殖。本研究結(jié)果進(jìn)一步佐證了土壤肥力(養(yǎng)分)增高可促進(jìn)土壤微生物多樣性升高的論點(diǎn)[38]。石英砂巖和長(zhǎng)石石英砂巖馬尾松由于生長(zhǎng)緩慢,凋落物和根系分泌物較少,限制了固氮菌生長(zhǎng),因而導(dǎo)致固氮微生物豐富度和多樣性指數(shù)較低。
本研究中土壤有機(jī)碳、全氮和堿解氮與nifH基因豐度和多樣性指數(shù)之間呈顯著正相關(guān)性關(guān)系,進(jìn)一步表明不同母巖發(fā)育馬尾松土壤有機(jī)碳、全氮和堿解氮含量差異是影響固氮菌數(shù)量和多樣性的重要因素。有研究表明土壤氮素含量是微生物生長(zhǎng)的限制因子,其與微生物數(shù)量和群落組成關(guān)系密切[39]。然而,本研究中土壤銨態(tài)氮和硝態(tài)氮含量與固氮菌豐度和多樣性指數(shù)并無(wú)顯著相關(guān)性,這和何冬華[40]等研究結(jié)果一致,說(shuō)明土壤環(huán)境因子對(duì)固氮菌的影響較為復(fù)雜。
3.2 固氮菌群落與土壤質(zhì)量之間的關(guān)系
固氮菌群落組成對(duì)土壤地力維持和植物生長(zhǎng)有重要影響[41]。Poly等[23]研究表明,土壤類型和養(yǎng)分狀況對(duì)固氮菌群落結(jié)構(gòu)影響較大。本研究中四類母巖土壤優(yōu)勢(shì)門(mén)和屬類群相似,但不同母巖中優(yōu)勢(shì)類群相對(duì)豐度存在顯著差異(圖5);層次聚類和NMDS分析也表明,四類母巖發(fā)育土壤固氮菌群落結(jié)構(gòu)分離明顯,因此母巖對(duì)土壤固氮菌群落結(jié)構(gòu)有顯著影響。本研究中變形菌門(mén)和藍(lán)藻門(mén)是主要優(yōu)勢(shì)類群,且在不同母巖發(fā)育土壤中差異顯著。相關(guān)研究認(rèn)為,變形菌門(mén)廣泛分布于土壤中且有較高的相對(duì)豐度,屬于異養(yǎng)型微生物,具有嗜營(yíng)養(yǎng)特點(diǎn),與土壤有機(jī)碳含量關(guān)系緊密,可用來(lái)指示土壤肥力狀況[42- 43]。Anderson等[44]研究表明,植物可以增強(qiáng)母巖土壤風(fēng)化速率和促進(jìn)養(yǎng)分釋放。玄武巖土壤在有機(jī)碳含量高和馬尾松生長(zhǎng)旺盛共同作用下促進(jìn)變形菌門(mén)菌群生長(zhǎng),導(dǎo)致變形菌門(mén)相對(duì)豐度顯著高于其他母巖。其中,慢生根瘤菌屬相對(duì)豐度在玄武巖土壤最高;然而眉藻屬相對(duì)豐度在玄武巖土壤最低;可知變形菌門(mén)中不同屬群對(duì)相同環(huán)境因子的響應(yīng)存在差異,具有各自的生態(tài)位。藍(lán)藻門(mén)為水生環(huán)境中優(yōu)勢(shì)固氮類群,在濕潤(rùn)環(huán)境下生長(zhǎng)較好[45- 46]。目前,關(guān)于藍(lán)藻門(mén)對(duì)氮固定的貢獻(xiàn)大小一直未達(dá)成共識(shí)[47]。Beversdorf等[48]研究認(rèn)為藍(lán)藻門(mén)對(duì)氮固定的貢獻(xiàn)較大,然而Ferber等[49]研究認(rèn)為藍(lán)藻門(mén)對(duì)氮的固氮在整個(gè)環(huán)境氮庫(kù)中貢獻(xiàn)較小。本研究中藍(lán)藻門(mén)相對(duì)豐度在玄武巖土壤最低,變余砂巖土壤最高;可能是由于玄武巖土壤為砂質(zhì)粘壤土[19],土壤透水好,抑制了藍(lán)藻門(mén)固氮菌的生長(zhǎng);變余砂巖土壤為壤質(zhì)粘土[19],土壤透水性差,且7月雨水較多,土壤易于板結(jié)和積水,有利于其生長(zhǎng)繁殖;同時(shí)變余砂巖土壤藍(lán)藻門(mén)相對(duì)豐度高導(dǎo)致銨態(tài)氮含量較高,然而銨態(tài)氮含量與其他母巖差異不顯著,可見(jiàn)藍(lán)藻門(mén)在變余砂巖土壤中可能對(duì)氮固定的貢獻(xiàn)不大。厚壁菌門(mén)具有寡營(yíng)養(yǎng)特點(diǎn),同時(shí)分泌大量有害物質(zhì),干擾植物正常氮素代謝,不利于植物生長(zhǎng)[13]。厚壁菌門(mén)相對(duì)豐度在長(zhǎng)石石英砂巖土壤最高,因此推測(cè)長(zhǎng)石石英砂巖馬尾松生長(zhǎng)緩慢可能與該菌群有關(guān)。疣微菌門(mén)極富生態(tài)多樣性,對(duì)環(huán)境耐受能力強(qiáng),具有降解烴類能力[50]。在馬尾松生長(zhǎng)過(guò)程中,疣微菌門(mén)固氮的同時(shí)所降解的烴類物質(zhì)可能是馬尾松所產(chǎn)生的次級(jí)代謝產(chǎn)物,次級(jí)代謝產(chǎn)物的積累將不利于其生長(zhǎng)[51]。本研究中疣微菌門(mén)相對(duì)豐度在玄武巖和長(zhǎng)石石英砂巖土壤較高,變余砂巖土壤較少,石英砂巖土壤未檢測(cè)到。因此,不同母巖發(fā)育土壤中疣微菌門(mén)的減少或消失,可能也是抑制馬尾松生長(zhǎng)的一個(gè)重要因素。
本研究發(fā)現(xiàn),土壤有機(jī)碳、全氮、堿解氮和微生物量氮是固氮菌群落結(jié)構(gòu)的主要影響因素。Orr等[52]研究也認(rèn)為土壤有機(jī)碳和氮對(duì)固氮菌群落結(jié)構(gòu)有顯著影響,同時(shí)有機(jī)碳、全氮和堿解氮相互之間有顯著相關(guān)性;由此可知,固氮菌群落結(jié)構(gòu)可能由幾個(gè)關(guān)聯(lián)性較強(qiáng)的化學(xué)指標(biāo)共同進(jìn)行調(diào)控。綜上所述,玄武巖土壤有機(jī)碳、全氮、堿解氮和微生物量氮顯著高于其他母巖,土壤肥力較高,有利于馬尾松生長(zhǎng),對(duì)優(yōu)化固氮菌群落結(jié)構(gòu)和促進(jìn)土壤氮固定具有重要作用。今后研究中需要開(kāi)展相應(yīng)的野外試驗(yàn),與池栽試驗(yàn)進(jìn)行相互印證;同時(shí)對(duì)參與馬尾松凋落物分解和拮抗致害真菌等重要微生物功能類群進(jìn)行深入的研究,有利于進(jìn)一步解釋不同母巖發(fā)育馬尾松土壤質(zhì)量差異的微生物機(jī)理,為馬尾松適地造林提供充分的理論依據(jù)。
4 結(jié)論
四類母巖土壤固氮菌(nifH)基因豐度和群落α多樣性指數(shù)差異顯著,表現(xiàn)為玄武巖>變余砂巖>石英砂巖>長(zhǎng)石石英砂巖。變形菌門(mén)和藍(lán)藻門(mén)相對(duì)豐度占到固氮菌總量的87.99%—95.29%,是四類母巖土壤共有的優(yōu)勢(shì)群落,其中玄武巖土壤所占比例最高。厚壁菌門(mén)和疣微菌門(mén)通過(guò)自身代謝產(chǎn)物對(duì)馬尾松生長(zhǎng)具有重要影響。土壤有機(jī)碳、全氮和堿解氮是影響四類母巖土壤固氮菌豐度、群落結(jié)構(gòu)和α多樣性的主要因素。因此,馬尾松在不同母巖發(fā)育土壤栽種時(shí),應(yīng)根據(jù)母巖特性采取有效措施,改善土壤養(yǎng)分狀況,優(yōu)化土壤固氮菌群落結(jié)構(gòu),提高土壤固氮能力,實(shí)現(xiàn)人工林速生豐產(chǎn)及可持續(xù)經(jīng)營(yíng)。
猜你喜歡
音樂(lè)探索(2022年2期)2022-05-30 21:01:37
小讀者(2021年2期)2021-03-29 05:03:48
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國(guó)特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
紅領(lǐng)巾·萌芽(2016年1期)2016-09-10 07:22:44
