基于葉綠體DNA片段的槭屬雞爪槭組系統(tǒng)進化和生物地理學研究
2020-11-13 01:09:44李俊清
生態(tài)學報 2020年17期
高 健,于 濤,李俊清,*
1 內(nèi)蒙古科技大學包頭師范學院資源與環(huán)境學院,包頭 014030 2 北京林業(yè)大學森林資源生態(tài)系統(tǒng)過程北京市重點實驗室,北京 100083
槭屬(AcerL.)是北半球地區(qū)僅次于櫟屬(QuercusL.)的第二大樹木屬,屬內(nèi)物種豐富(約124—200種),且廣泛分布于亞洲、歐洲及北美洲等地[1- 2]。東亞,特別是中國長江流域及其以南地區(qū),是槭屬植物現(xiàn)代的多樣性中心[3]。雞爪槭組(sectionPalmataPax)是槭屬中物種多樣性最為豐富的分類組,組內(nèi)物種為多年生喬木或灌木,落葉或常綠[1]。雞爪槭組內(nèi)物種形態(tài)特征變化豐富,具有掌狀葉片從不裂(例如羅浮槭、濱海槭等)到3—13裂、葉邊緣從全緣到鋸齒、葉裂片從淺裂到深裂乃至全裂、花序類型多樣(圓錐花序、傘房花序等)及果實長度變化較大(1—5 cm)等特征[2]。此外,雞爪槭組物種集中分布于東亞地區(qū)(除了AcercircinatumPursh.分布于北美),是東亞槭屬植物的代表類群[1- 2]。
雞爪槭組物種具有重要的生態(tài)價值和科研價值。例如雞爪槭(A.palmatumThunberg ex Murray)作為北溫帶廣泛分布的園藝觀賞樹種,其種內(nèi)人工選育出多種不同葉形及葉片顏色的雞爪槭變種、變型等,在園林造景、植被恢復等生態(tài)建設中發(fā)揮著重要的作用[4]。東亞地區(qū)作為雞爪槭組物種主要的分布區(qū),其地貌類型復雜多樣,且在末次冰期時未被大面積冰川所覆蓋,其生物地理歷史相較歐美地區(qū)來說更為復雜[5]。所以,雞爪槭組物種是了解東亞地區(qū)植物進化歷史的一個代表性材料。因此,本文對雞爪槭組物種的起源、組內(nèi)物種的分化時間、物種的擴散路徑等進行了分析。該研究有助于了解雞爪槭組的進化歷史,并為東亞地區(qū)植物的生物地理歷史提供了一個重要的范例。
1 材料與方法
1.1 試驗材料
選取雞爪槭組內(nèi)18個物種(包含亞種),共27個個體作為研究材料(表1)。物種的鑒定與分類依據(jù)《Maples of the world》中的定義[2]。部分物種的序列于Genbank中下載得到(表1)。每個分類群選取1—3株個體,采集到的植物幼嫩葉片放入置有變色硅膠的取樣袋中干燥、備用。同時,壓制憑證標本,存放于內(nèi)蒙古科技大學包頭師范學院資源與環(huán)境學院。
1.2 DNA提取、PCR及測序
每份樣品取約0.2 g干燥葉片,全基因組DNA的提取使用植物基因組DNA提取試劑盒(TIANGEN Beijing China)。依據(jù)本課題組前期對于雞爪槭高分辨率DNA條形碼的篩選結果,選取葉綠體基因片段psbA-trnH、rpl16和trnL-trnF作為本研究的遺傳片段[4]。葉綠體基因片段的PCR擴增均在LabnetMultiGeneTM96-well Gradient Thermal Cycler上完成。PCR反應體系為20 μL,包含DNA模板1 μL、Taq PCR Master Mix(萊楓,上海)10 μL、正反引物各1 μL(表2)、二次水(ddH2O)7 μL。PCR反應程序為:94 ℃預變性3 min;94 ℃變性30 s,52—58 ℃退火30 s,72 ℃延伸60 s,共30個循環(huán);最后72 ℃延伸10 min。PCR擴增產(chǎn)物經(jīng)1%瓊脂糖凝膠電泳檢測并純化后,送往北京生工生物工程股份有限公司在ABI3730XL遺傳分析儀上進行雙向測序。
1.3 系統(tǒng)發(fā)育分析
序列通過ClustalX[11]進行比對,并使用BioEdit 7.1.11[12]進行人工矯正。此外,對3個DNA片段的相容性進行ILD檢測[13],1000次重復啟發(fā)式搜索后顯示片段間沒有顯著的沖突(P= 0.16)。故將3個片段合并后進行后續(xù)的聯(lián)合分析。
金錢槭(DipteroniasinensisOliver)與云南金錢槭(DipteroniadyerianaHenry)在系統(tǒng)發(fā)育分析中作為外類群[14- 15]。最大似然法(ML)、最大簡約法(MP)以及貝葉斯推理(BI)等分析分別使用PhyML 3.0[16]、PAUP*4.0b10[17]、MrBayes 3.2[18]進行計算。最佳的取代模型通過Modeltest 3.7[19]進行分析獲取。

表1 實驗物種基本信息

表2 基因片段PCR擴增的引物信息
1.4 分子定年及多樣性速率分析
采用BEAST v.1.8.2[20]構建系統(tǒng)發(fā)育樹,并推測雞爪槭組物種間的分化時間。采用槭屬與金錢槭屬的分化時間(62 Mya)[21]和雞爪槭組的分化時間(27.99 Mya)[22]作為化石校準點進行分子定年。進行1000000000次的MCMC模擬運算,每100000筆運算進行一次取樣,總取樣結果的前2500次取樣不計入后續(xù)分析當中。
通過隨機分枝過程(stochastic branching process,SBP)[23]繪制支系隨時間累積趨勢圖(lineages through time,LTT),采用reversible-jump馬爾可夫鏈蒙特卡爾(rjMCMC)模擬進行貝葉斯非參數(shù)估計(Bayesian non-parametric estimation)[24]來評估多樣性速率。利用R語言的package APE[25]繪制LTT,并以APE中的mltt.plot功能,重新繪制以貝葉斯進化分析軟件構建的1000次取樣,以此來評估支系隨時間累積的偏差。rjMCMC分析進行1000000次模擬,其中每1000次模擬提取1筆數(shù)據(jù),對前10%的不穩(wěn)定數(shù)據(jù)剔除后進行后續(xù)分析。γ統(tǒng)計被用來通過時間評估多樣性速率的變異規(guī)律[26]。
1.5 祖先分布區(qū)重建
參考香茶菜屬(Isodon)[27]、芭蕉科(Musaceae)[28]等祖先分布區(qū)重建研究中,依據(jù)地形對亞洲地理區(qū)塊進行劃分的方法,再結合本研究中雞爪槭組物種主要分布于東亞、東南亞的特點,把雞爪槭組物種的分布區(qū)范圍主要劃分為4個區(qū)塊:(A)中國華東、華中、華南、華北、東北部分地區(qū)及朝鮮半島;(B)中國西南及東南亞北沿地區(qū)(云貴高原、青藏高原等地);(C)日本地區(qū);(D)中國臺灣島。采用R語言中的BioGeoBEARS軟件包進行雞爪槭組生物地理歷史的推測[29]。
2 結果與分析
2.1 DNA擴增片段分析
序列比對后rpl16片段為909bp,含114個(12.5%)變異位點和74個(8.1%)有效信息位點;psbA-trnH片段為421bp,含72個(17.1%)變異位點和48個(11.4%)有效信息位點;trnL-trnF片段為902bp,含155個(17.1%)變異位點和109個(12.1%)有效信息位點。3個葉綠體DNA片段聯(lián)合后,矩陣長度為2232bp,具有341個(15.2%)變異位點和231個(10.3%)有效信息位點。
2.2 系統(tǒng)發(fā)育關系
三種方法(ML,MP和BI)所構建的物種系統(tǒng)發(fā)育關系基本一致。18個雞爪槭組物種(包含亞種)的27個個體形成單系群(99/95/1.0)(節(jié)點A)(圖1)。稀花槭(A.pauciflorum)與濱海槭(A.sino-oblongum)的親緣關系較近(99/57/0.64)。在節(jié)點B中,藏南槭(A.campbellii)種下的4個亞種之間沒有歸并為單系群。此外,羽扇槭(A.japonicum)與雞爪槭(A.palmatum)被高度支持為單系群(96/94/1.0);秀麗槭(A.elegantulum)與羅浮槭(A.fabri)間的親緣關系比較近(76/70/0.96)(圖1)。
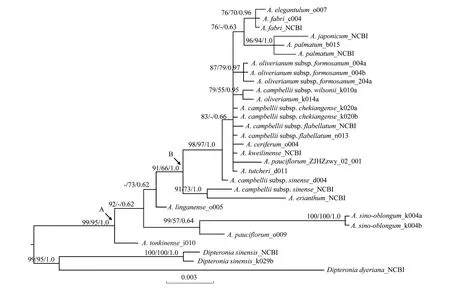
圖1 基于rpl16,trnH-psbA以及trnL-F片段合并序列構建的系統(tǒng)發(fā)育樹Fig.1 The tree combined chloroplast rpl16,trnH-psbA and trnL-F regionsML、MP分析中自展值高于50%及BI分析中后驗概率高于50%被依次標于分枝上
2.3 分化時間及多樣性速率估計
分子定年得出的物種樹(圖2)與基因樹(圖1)系統(tǒng)發(fā)育關系基本一致。結果顯示:粗柄槭(A.tonkinense)較早從雞爪槭組分化而出;臨安槭(A.linganense)(浙江、安徽南)和飛蛾槭(A.sino-oblongum)(廣東沿海)兩個現(xiàn)代的局限分布種,分別約在19.92 Mya與15.58 Mya分化而來;12.42 Mya分化出雞爪槭與羽扇槭的共同祖先,進而在4.18Mya分化出這兩個物種;五裂槭(A.oliverianum)與其亞種臺灣五裂槭(A.oliverianumsubsp.formosanum)的分化時間約為2.91 Mya;藏南槭在4.71 Mya開始其種內(nèi)亞種的分化(圖2b)。LTT結果顯示,雞爪槭組內(nèi)物種的多樣性速率保持相對穩(wěn)定(圖2a)。
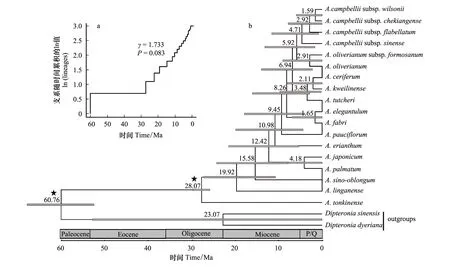
圖2 雞爪槭組物種的支系隨時間累積趨勢(a)及基于3個葉綠體DNA片段構建的雞爪槭組植物分子定年結果圖(b)Fig.2 The lineage-through-time (LTT) of Sect. Palmata (a) and Chronogram for Sect. Palmata reconstructed by three cpDNA sequences (b)
2.4 生物地理分析
通過BioGeoBears分析得出,進行雞爪槭組生物地理分析的最佳模型是DEC模型(AIC=96.54,lnL=-45.92)。根據(jù)DEC模型的分析得出,A加B區(qū)域可能是雞爪槭組物種的祖先分布區(qū)(圖3)。此外,可能曾發(fā)生物種從亞洲大陸擴散至周邊島嶼的情況,從而形成了日本特有種羽扇槭,我國臺灣特有種臺灣五裂槭以及目前分布于我國華南及臺灣地區(qū)的嶺南槭(A.tutcheri)(圖3)。
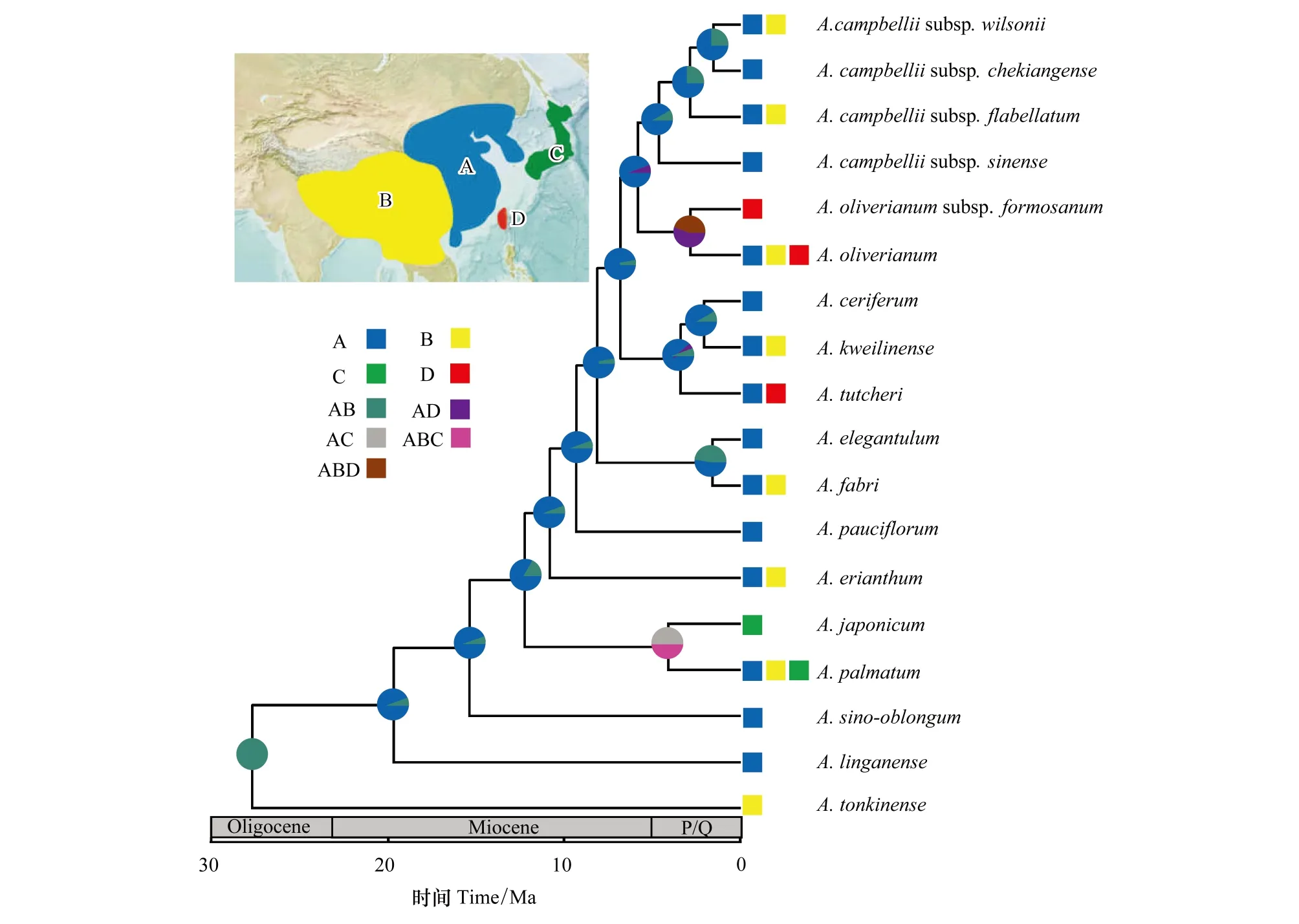
圖3 雞爪槭組植物祖先分布區(qū)重建Fig.3 The reconstruction of ancestral area for Sect. Palmata
3 討論與結論
3.1 組內(nèi)種間的系統(tǒng)位置
雞爪槭組內(nèi)物種(包含亞種)被高度支持為單系群(99/95/1.0)(圖1)[21,30]。在槭屬分類書籍《Maples of the world》中記載A.campbellii中有4個亞種,理論上應當形成單系群,但結果顯示這4個亞種分散在不同的分支中(圖1)。而在相對較新的分類學著作《Flora of China》中,這4個亞種被處理為4個獨立的物種,即:毛脈槭(A.campbelliisubsp.chekiangense)對應A.pubinerve;三峽槭(A.campbelliisubsp.wilsonii)對應A.wilsonii;扇葉槭(A.campbelliisubsp.flabellatum)對應A.flabellatum;中華槭(A.campbelliisubsp.sinense)對應A.sinense。此外,依據(jù)形態(tài)學證據(jù),4個物種也存在較明顯的差別。例如三峽槭葉片較其他3個物種(多為5裂,少7裂)來說多為3裂(少5裂),中華槭葉片較扇葉槭、毛脈槭(紙質或膜質)來說近革質,扇葉槭子房與花盤較毛脈槭(有毛)來說無毛等等。從地理分布上分析可以得出,4個物種主要集中分布于長江以南地區(qū),但三峽槭、扇葉槭側重分布于華南南部及西南地區(qū)(甚至延伸至越南、緬甸等地),而毛脈槭、中華槭側重分布于華南及華東南部等地。所以,基于本研究結果以及形態(tài)特征、地理分布等證據(jù)支持《Flora of China》的分類結果,即把這4個亞種分別作為獨立的物種進行分類處理。此外,研究結果支持雞爪槭與羽扇槭間親緣關系較近(圖1)。
3.2 雞爪槭組的起源
雞爪槭組的起源時間約為28 Mya左右(漸新世中晚期)[22],且物種多樣性在中新世時出現(xiàn)大幅度的增加(圖2)。在漸新世中晚期至中新世,全球的氣候變暖致使南極冰川的覆蓋范圍縮小[31],且在中新世中期(17—15 Mya)出現(xiàn)了氣溫的高點[32]。Donoghue和Smith對66個物種間斷分布格局的形成進行了總結,得出絕大多數(shù)北溫帶森林物種在亞洲起源及物種多樣化,且中新世是物種多樣化的活躍期之一[33]。此外,在漸新世晚期至中新世初期,昆蟲出現(xiàn)了植食現(xiàn)象(insect herbivory),隨后昆蟲的物種多樣性在中新世得到了大大的提高[34]。總之,氣候變化可能是影響雞爪槭組物種起源及多樣性增加的一個主要驅動力。
通過雞爪槭組的生物地理分析得出,分布于我國廣西、貴州南部、西藏東南部、云南東南部以及馬來西亞、泰國、越南北部等地區(qū)的粗柄槭是組內(nèi)較為早期分化出的物種(圖3),也就是說雞爪槭組中相對較為原始物種的分布區(qū)位于我國西南部及東南亞北部。鄭維艷等指出我國西南部是亞熱帶常綠闊葉林許多科屬的保存中心、現(xiàn)代分布中心和分化中心,且亞熱帶常綠闊葉林植物區(qū)系可能起源于我國的西南部[35]。徐廷志曾提出槭樹科的起源地可能在我國橫斷山區(qū)附近[3,36],而雞爪槭組在槭屬中又被認為是較原始的類群[3,21]。加上本研究結果支持雞爪槭組起源于A區(qū)域加B區(qū)域(圖3),所以有理由推斷雞爪槭組的起源地位于我國西南部地區(qū)。這也從側面支持了槭樹科起源于橫斷山區(qū)[3,36]及槭屬起源于亞洲的結論[22]。
3.3 雞爪槭組的生物地理歷史
雞爪槭組植物在進化的歷程中,存在多次的生物地理事件。例如,雞爪槭與羽扇槭約在4.18 Mya出現(xiàn)分化(圖2)。雞爪槭的現(xiàn)代分布區(qū)域極廣,廣泛分布于中國、朝鮮半島及日本地區(qū),而羽扇槭作為雞爪槭最近緣的物種卻僅僅局限分布于日本。與之相類似的還有五裂槭(廣布種)與臺灣五裂槭(臺灣特有種),兩者的分化時間約在2.91 Mya(臺灣島形成時間約3—2 Mya)。兩組物種的分化可能與第四紀冰期(開始于3—2 Mya)有直接的聯(lián)系。第四紀冰期氣候的變化導致了中國東海海平面的抬升與降低,在海平面下降時,亞洲大陸與日本、中國臺灣等島嶼之間形成了東海陸橋,為分布于該地區(qū)的生物提供了遷徙的走廊[37,38]。而且,在4—3 Mya左右時,青藏高原的持續(xù)隆起[39],太平洋季風增強帶來的豐沛降水[40]均對分布于東亞地區(qū)物種的擴散和多樣化創(chuàng)造了客觀條件。所以,對于雞爪槭和五裂槭來講,由于兩物種較強的適應能力,冰期時可能借由東海陸橋分別擴散至日本島、中國臺灣島,而冰后期兩島嶼與亞洲大陸不再相連,地理隔離導致物種局地適應,而分別形成了現(xiàn)在的羽扇槭和臺灣五裂槭(圖3)。此外,依據(jù)中國植物志中的介紹,嶺南槭種下包含分布于我國華南地區(qū)的嶺南槭(原變種)與僅分布于中國臺灣地區(qū)的小果嶺南槭[1],所以嶺南槭也可能曾借由東海陸橋進行物種的遷徙(圖3)。
值得一提的是,縱觀雞爪槭組生物地理特征的結果,不難發(fā)現(xiàn)B區(qū)域物種是由A區(qū)域多次獨立擴散而來的(圖3)。而且,B區(qū)域物種同一進化分支中的姐妹種,多分布于A區(qū)域南部(亞熱帶地區(qū)),例如:稀花槭,秀麗槭,杈葉槭(A.ceriferum),中華槭等[1]。B區(qū)域多為高原地形,海拔較高,氣溫相對同緯度地區(qū)較低。所以,在周期性環(huán)境壓力的背景下(例如:冰期與間冰期),雞爪槭組物種為尋求合適生境,可存在多次往B區(qū)域擴散的現(xiàn)象(圖3)。Qiu等通過對多種落葉闊葉林植物進行研究發(fā)現(xiàn),亞熱帶地區(qū)闊葉林存在局部地區(qū)高低海拔遷移的現(xiàn)象[38]。由此而言,雞爪槭組物種在進化歷程中,可能存在方向性的擴散過程。
綜上所述,可以推斷得出以下結論:(1)我國西南地區(qū)可能是雞爪槭組的起源地;(2)東海陸橋的隱沒與抬升可能影響著羽扇槭、臺灣五裂槭及嶺南槭等雞爪槭組物種的遷徙與擴散過程;(3)分布于亞熱帶亞洲大陸的雞爪槭組物種,可能存在多次往同緯度高海拔地區(qū)擴散的現(xiàn)象。
致謝:感謝臺灣師范大學生命科學系廖培鈞教授、黃秉宏博士在數(shù)據(jù)處理上的幫助。
猜你喜歡
課堂內(nèi)外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內(nèi)外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
電子制作(2018年18期)2018-11-14 01:48:24
汽車觀察(2018年10期)2018-11-06 07:05:26
山東工業(yè)技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·少年版(2015年1期)2015-07-07 17:15:12
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
