許氏平鲉幼魚對飼料中維生素B6 需求量的研究*
2020-11-03 06:08:52王際英李寶山王成強王世信孫永智馬長興
漁業科學進展 2020年6期
周 瑩 王際英 李寶山 王成強 王世信 孫永智 馬長興
(1. 上海海洋大學 水產科學國家級實驗教學示范中心 農業農村部魚類營養與環境生態研究中心水產動物遺傳育種中心上海市協同創新中心 上海 201306;2. 山東省海洋資源與環境研究院 山東省海洋生態修復重點實驗室 煙臺 264006)
維生素B6是一種水溶性維生素,以吡哆醇、吡哆醛和吡多胺3種形式存在(Halsted, 2003)。維生素B6與生物體內營養物質的代謝密切相關。首先,維生素B6作為生物體內轉氨酶和脫羧酶的輔酶,參與催化氨基酸轉氨、脫羧、消旋、脫硫和裂解等反應,在氨基酸代謝中起重要作用(Shiau et al, 2003);其次,作為糖原磷酸化酶的輔酶參與碳水化合物代謝,為糖異生提供碳架,維持血糖濃度;同時,其與必需脂肪酸的代謝、mRNA的合成,糖皮質激素及白蛋白的基因表達有關(Zehra et al, 2018; Natori et al, 1997);此外,它還參與神經物質如γ-氨基丁酸(GABA)、多巴胺、組胺、5-羥色胺(5-HT)等的生物合成,從而影響神經系統的發育和功能(Huang et al, 2005; Akhtar et al,2010)。水產動物缺乏維生素B6會產生相應的缺乏癥,但維生素B6過量并不會對水產動物產生明顯的毒副作用。通過生長指標、肝臟維生素B6的含量、轉氨酶活性等評價指標,確定了水產動物維生素B6的需求量約為1~12 mg/kg(He et al, 2009; Zehra et al, 2018;Hosokawa et al, 1992),其中,斜帶石斑魚(Epinephelus coioides)維 生 素B6需 求 量 為1.75~2.22 mg/kg(Huang et al, 2005),缺乏時表現為厭食,生長不良,驚厥,螺旋狀游泳等;大菱鲆(Scophthalmus maximus)維生素B6需求量為1~2.5 mg/kg(Adron et al, 1978),缺乏時表現為生長不良,厭食,運動失調,水腫,神經紊亂等;金頭鯛(Spatrus aurata) 維生素B6需求量為1.97 mg/kg (Kissil et al, 1981),缺乏時表現為生長遲緩,死亡率高,游泳行為不穩定和周圍神經退行性改變等。
許氏平鲉(Sebastes schlegeli),隸屬于鲉形目(Scorpaeniformes)、鲉科(Scorpaenidae)、平鲉屬(Sebastes),又稱黑鲪、黑頭等,是一種冷溫性卵胎生的魚類,分布于我國黃海、渤海和東海,國外分布于朝鮮、日本和鄂霍次克海南部(劉蟬馨, 1988; 朱龍等, 1999)。許氏平鲉肉質鮮美,在沿海有黑石斑的美譽,是人工魚礁資源增殖和網箱養殖的理想種類(冒樹泉等, 2014)。目前,關于許氏平鲉營養需求研究主要涉及蛋白質、脂肪、糖類、礦物質及飼料添加劑等方面(Lee et al,2002、2009; Cho et al, 2015; Katya et al, 2017),維生素營養需求的報道僅見維生素C (Lee et al, 1998)和維生素E (Bai et al, 1998),B 族維生素研究方面尚處于空白。本研究通過在飼料中添加不同梯度的維生素B6,研究其對生長、肝臟酶活和轉氨酶基因表達等指標的影響,確定許氏平鲉幼魚對維生素B6的最適需求量,為其配合飼料的開發提供參考。
1 材料與方法
1.1 實驗飼料
以魚粉、酪蛋白為主要蛋白源,魚油為主要脂肪源,設計粗蛋白約為49.56%、粗脂肪約為9.12%的基礎飼料配方。在基礎飼料中分別添加0、1、2、4、8和32 mg/kg 的鹽酸吡哆醇(純度為98.6%,購自上海麥克林生化科技有限公司),配制維生素B6實際含量為 2.08、3.25、4.16、6.32、10.17 和 31.14 mg/kg 6 組等氮等能的實驗飼料,命名為D1、D2、D3、D4、D5 和D6 組,其中,D1 為對照組。表1 為實驗飼料的配方及營養組成。
各原料經粉碎過80 目標準篩,按配比稱重后,逐級擴大法混合均勻,添加魚油和適宜的蒸餾水,再次混合混勻,用小型飼料擠壓機制成粒徑為2 和4 mm的顆粒飼料,60℃烘干,用塑料袋密封后于–20℃冰箱保存備用。
1.2 實驗用魚及飼養管理
實驗所用許氏平鲉幼魚購自山東日照傳真養殖場,養殖實驗在山東日照片聯綠色養殖示范基地的水泥養殖池內掛網箱(60 cm×60 cm×50 cm)中進行。正式實驗前采用對照組飼料馴化,14 d 后挑選體質健壯、大小均一、初始體重為(36.35±0.06) g 的實驗魚540 尾。實驗隨機分為6 個組,每組3 個重復,每個網箱30 尾魚,共18 個網箱。養殖期間持續充氣,養殖用水為深井海水,采用流水養殖。水溫為16℃~18℃,溶解氧(DO)>5.0 mg/L,pH 為7.0 左右,氨氮和亞硝態氮<0.05 mg/L。每天08:00 和16:00 投喂2 次,初始投喂量為魚體重的1%,并按其攝食情況調整,每日記錄死魚數量和重量。實驗進行63 d。
1.3 樣品采集及分析
養殖實驗結束后,禁食24 h,記錄每個網箱魚的數量并稱重,每個網箱隨機取4 尾魚,用于魚體常規成分測定,另取8 尾魚測量體重及體長后,尾靜脈取血,解剖分離內臟,分別稱量內臟團、肝胰臟、腸道的重量。全魚、背肌、肝臟樣品置于–80℃條件下保存,待測。每桶中隨機取2 尾魚,無菌環境下解剖,取肝尖置于無RNase 離心管中并迅速置于液氮中速凍,后于–80℃超低溫冰箱中保存,用于組織中相關基因表達量的分析。
實驗飼料及樣品中水分采用105℃恒重法測定(GB/T6435-2014),粗蛋白采用凱氏定氮法測定(GB/T6432-2018),粗脂肪采用索氏抽提法測定(GB/T6433-2006),粗灰分采用550℃失重法測定(GB/T 6438-2007),能量采用燃燒法測定(IKA, C6000, 德國)。
肝臟中維生素B6的含量采用上海酶聯生物科技有限公司試劑盒測定。超氧化物歧化酶(Superoxide dismutase, SOD)、丙二醛(Malondialdehyde, MDA)、谷草轉氨酶(Glutamic oxalacetic transaminase, GOT)和谷丙轉氨酶(Glutamic-pyruvic transaminase, GPT)均采用南京建成生物工程的試劑盒進行測定,肝臟組織蛋白含量采用考馬斯亮蘭法測定,酶活力單位定義參照試劑盒說明書。
使用Trizol 法提取總RNA,提取完成后使用核酸蛋白儀(Nanoprop 2000,美國)檢驗RNA 的濃度,并使用瓊脂糖凝膠電泳檢測RNA 的完整性。采用PrimeScriptTMRT reagent Kit with gDNA Eraser 反轉錄試劑盒(Takara,日本)去除基因組DNA 并反轉錄成cDNA 后于–80℃保存待用。根據本實驗室所得的許氏平鲉轉錄組數據,找到谷草轉氨酶(GOT1 和GOT2)、谷丙轉氨酶(GPT1 和GPT2)、酪氨酸轉氨酶(TAT)、絲氨酸轉氨酶(SerC)的基因序列,NCBI Nucleotide BLAST 進行同源性比對分析,確定同源性較高的基因序列,利用Primer 5.0 軟件設計引物,選用核糖體蛋白L17(RPL17)作為內參基因(Ma et al,2013),引物由上海生工生物技術有限公司合成,引物序列如表2 所示。實時熒光定量PCR 試劑盒為TaKaRa 的TB GreenTMPremix Ex TaqTMII (TliRNaseH Plus)。PCR 程序:95℃預變性30 s,1 個循環;95℃變性5 s,相應基因退火溫度下退火30 s,72℃延伸60 s,共40 個循環后增長至95℃。通過熒光定量PCR 儀得出目的基因及內參基因的Ct 值,按照2–??Ct計算目的基因的相對表達量。本研究中,RPL17、GOT1、GOT2、GPT1、GPT2、TAT 和SerC 的擴增效率分別為0.992、0.962、0.979、0.965、0.989、1.055 和1.027。
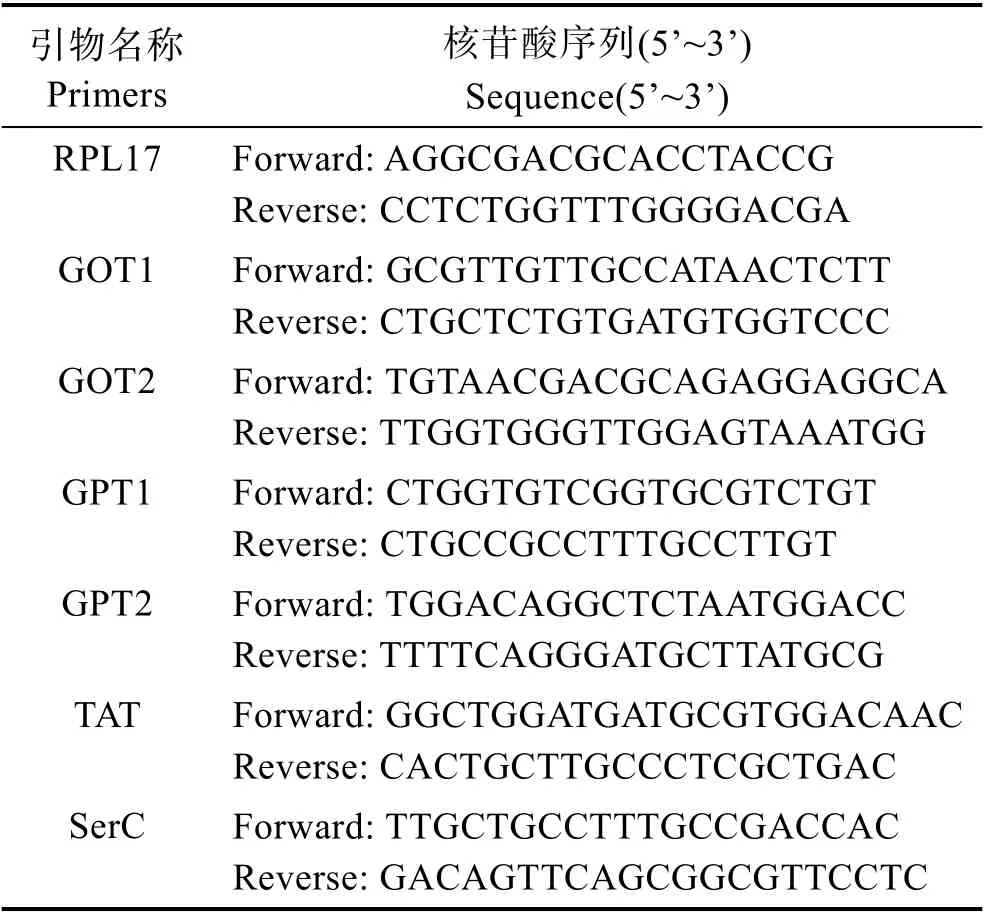
表2 基因表達的引物序列Tab.2 Primer sequences for gene expression
1.4 指標計算及數據統計
增重率(Weight gain rate, WGR, %)=(魚體末重–魚體初重)/魚體初重×100;
特定生長率(Specific growth rate, SGR, %/d) =(ln魚體末重–ln 魚體初重)/養殖周期×100;
攝食率(Feed intake, FI, %/d)=攝食飼料量/[(魚體初重+魚體末重)/2×養殖天數] ×100;
飼料系數(Feed conversion ratio, FCR)=攝食飼料量/(魚體末重–魚體初重);
蛋白質效率(Protein efficiency ratio, PER, %)=(魚體末重–魚體初重)/(攝食飼料量×飼料中粗蛋白的含量)×100;
臟體比(Viscerosomtic index, VSI, %)=內臟團重/魚體末重×100;
肝體比(Hepatosomatic index, HSI, %)=肝臟重/魚體末重×100;
腸體比(Intestine-somatic index, ISI,%)=腸道重/魚體末重×100;
肥滿度(Condition factor, CF)=魚體末重/體長3×100;
存活率(Survival rate, SR, %)=終末魚尾數/初始魚尾數×100。
數據統計使用SPSS 18.0 統計軟件進行單因素方差分析(One-way ANOVA)。每組數據采用平均值±標準誤(Mean±SE)表示,差異顯著(P<0.05)時用Duncan’s檢驗進行多重比較分析。采用折線模型擬合數據,確定許氏平鲉維生素B6的最適需求量。
2 結果
2.1 飼料中維生素B6對許氏平 鲉幼魚生長和形體指標的影響
由表3 可知,隨著維生素B6含量的升高,實驗魚增重率和特定生長率呈先上升后下降的趨勢,D3組顯著高于對照組(P<0.05)。D2~D6 組的攝食率無顯著差異(P>0.05),均顯著高于對照組(P<0.05)。D3 組飼料系數最低,D3~D6 組顯著低于對照組(P<0.05)。蛋白質效率呈現先上升后下降的趨勢,D3 組最高。D3 組的臟體比顯著高于對照組(P<0.05);D2~D5 組的肝體比顯著高于對照組(P<0.05);D2~D6 組的腸體比顯著高于對照組(P<0.05),呈先升后降的趨勢,在D3 組達到最高值;各組間的肥滿度和存活率無顯著差異(P>0.05)。
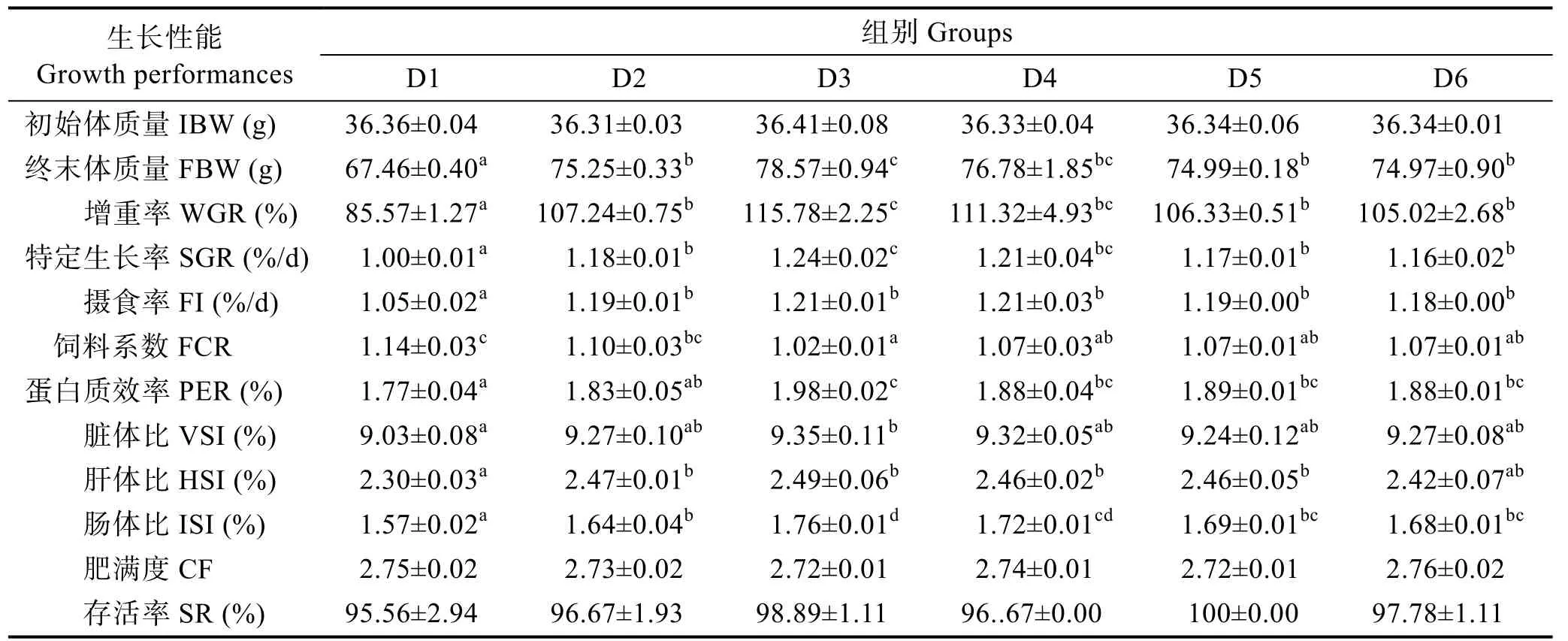
表3 飼料中維生素B6 對許氏平鲉幼魚生長和形體指標的影響Tab.3 Effects of dietary vitamin B6 on growth performance and body indices of juvenile S. schlegeli
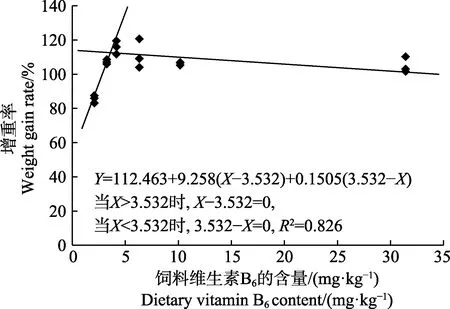
圖1 許氏平鲉幼魚增重率與飼料中維生素B6 含量的折線回歸關系Fig.1 The broken-line regression analysis based on weight gain rate and dietary vitamin B6 of juvenile S. schlegeli
以增重率為評價指標,經折線模型擬合后得許氏平鲉幼魚對飼料維生素B6的需求量為3.53 mg/kg(圖1)。
2.2 飼料中維生素B6對許氏平 鲉幼魚魚體常規成分的影響
由表4 可知,維生素B6含量對許氏平鲉幼魚全魚水分和粗灰分含量無顯著影響(P>0.05),全魚粗蛋白呈先上升后下降的趨勢,D3~D6 組的全魚粗蛋白顯著高于對照組(P<0.05),且在D4 組達到最高值;全魚粗脂肪呈先上升后下降的趨勢,對照組有最低值,D5組有最高值,D3~D6 組之間無顯著差異(P>0.05)。
許氏平鲉幼魚D3~D6 組肌肉水分含量顯著低于對照組(P<0.05);肌肉粗蛋白隨著維生素B6含量的增加呈先上升后下降的趨勢,對照組含量最低,D3~D6組無顯著差異(P>0.05);對照組肌肉粗脂肪呈先上升后下降的趨勢,D4 組最高;肌肉粗灰分含量無顯著變化(P>0.05)。
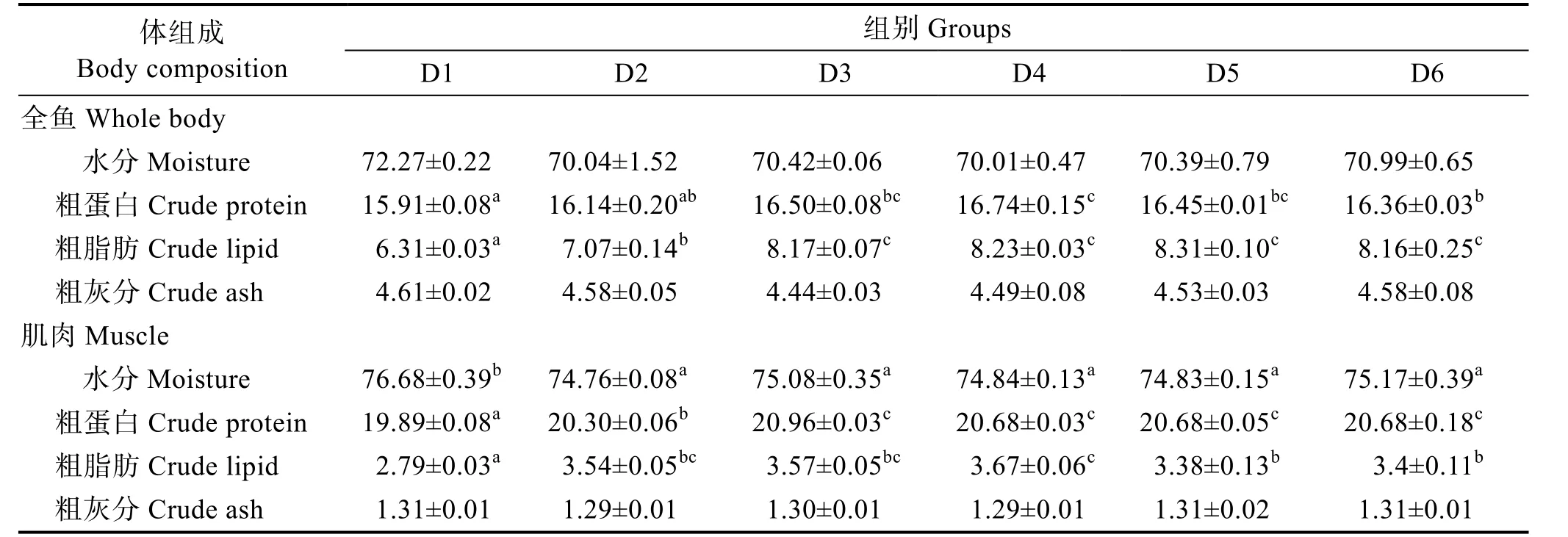
表4 飼料中維生素B6 對許氏平鲉幼魚魚體常規成分的影響(%濕重)Tab.4 Effects of dietary vitamin B6 on proximate composition of juvenile S. schlegeli (% wet weight)
2.3 飼料中維生素B6 對許氏平鲉幼魚肝臟酶活及維生素B6 含量的影響
由表5 可知,隨著維生素B6含量的增加,肝臟超氧化物歧化酶活力呈先上升后下降的趨勢,D3~D5組顯著高于對照組(P<0.05),D3 組最高。丙二醛含量呈現先下降后上升的趨勢,D2~D6 組顯著低于對照組(P<0.05),D3 組最低。D2~D6 組的肝臟谷草轉氨酶活力顯著高于對照組(P<0.05),D4~D6 組之間無顯著差異(P>0.05)。肝臟谷丙轉氨酶活力呈先上升后下降的趨勢,D3~D6 組顯著高于對照組(P<0.05)。肝臟維生素B6含量隨著飼料維生素B6含量的增加,呈先升高后平穩的趨勢,D2~D6 組顯著高于對照組(P<0.05),D3~D6 組間無顯著差異(P>0.05)。
以肝臟維生素B6含量為評價指標,經折線模型擬合后得許氏平鲉幼魚對飼料維生素B6的需求量為4.96 mg/kg(圖2);以肝臟谷丙轉氨酶活力為評價指標,經折線模型擬合后得許氏平鲉對飼料維生素B6的需求量為6.32 mg/kg(圖3)。
2.4 飼料中維生素B6 對許氏平鲉幼魚轉氨酶基因表達量的影響
隨著維生素B6含量的增加,谷草轉氨酶1(GOT1)基因相對表達量呈先上升后下降的趨勢,D4 組顯著高于對照組(P<0.05),D3~D6 組之間無顯著差異(P>0.05)(圖4);D2~D6 組谷草轉氨酶2(GOT2)基因相對表達量顯著高于對照組(P>0.05),D3 組為最大值,D4~D6 組間差異不顯著(P>0.05)(圖5);谷丙轉氨酶1(GPT1)和谷丙轉氨酶2(GPT2)基因相對表達量呈先上升后下降的趨勢,均在D4 組有最大值,且顯著高于對照組和D2 組(P<0.05),D3~D6 組差異不顯著(P>0.05)(圖6 和圖7);酪氨酸轉氨酶(TAT)呈先上升后下降的趨勢,D4 組有最大值,D3~D5 組顯著高于其他組(P<0.05)(圖8);絲氨酸轉氨酶(SerC)呈下降趨勢,D3~D6 組顯著低于對照組和D2 組(P<0.05),D6 組為最低值(圖9)。
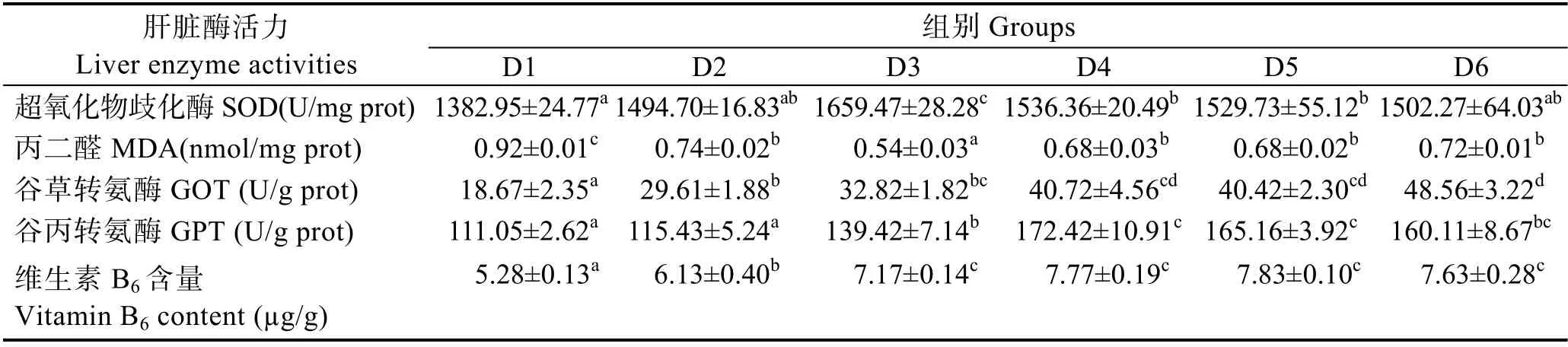
表5 飼料中維生素B6 對許氏平鲉幼魚肝臟酶活及維生素B6 含量的影響Tab.5 Effects of dietary vitamin B6 on liver enzyme activities and vitamin B6 content of juvenile S. schlegeli
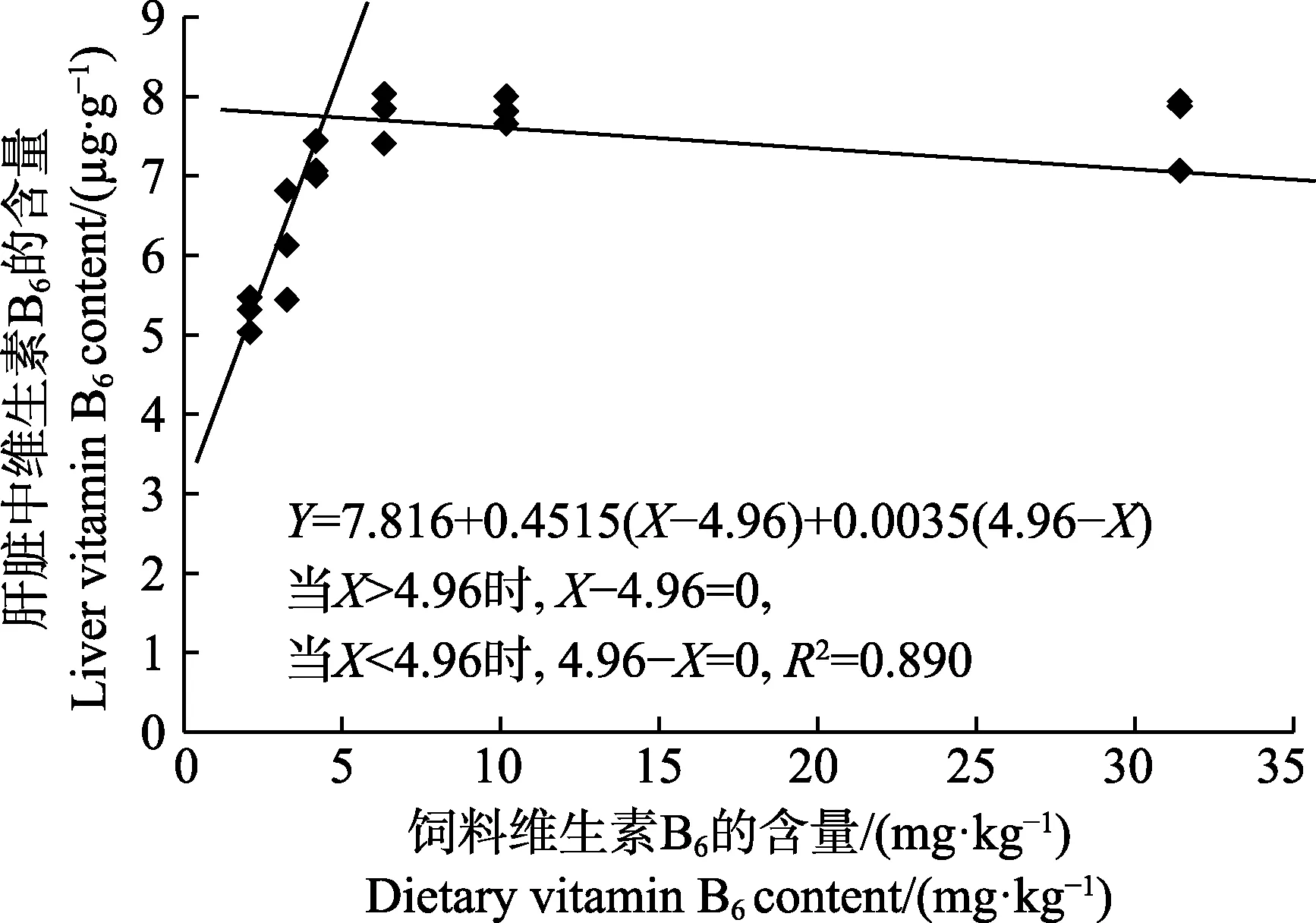
圖2 許氏平鲉幼魚肝臟維生素B6 含量與飼料中維生素B6 含量的折線回歸關系Fig.2 The broken-line regression analysis based on liver vitamin B6 content and dietary vitamin B6 content of juvenile S. schlegeli
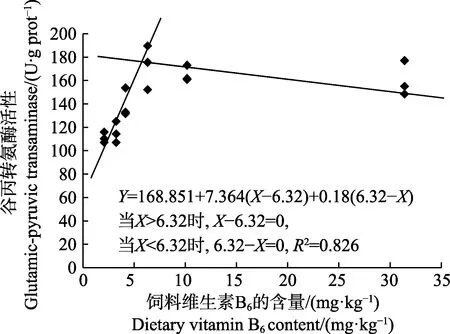
圖3 許氏平鲉幼魚肝臟谷丙轉氨酶活性與飼料中維生素B6 含量的折線回歸關系Fig.3 The broken-line regression analysis based on liver glutamic-pyruvic transaminase and dietary vitamin B6 content of juvenile S. schlegeli
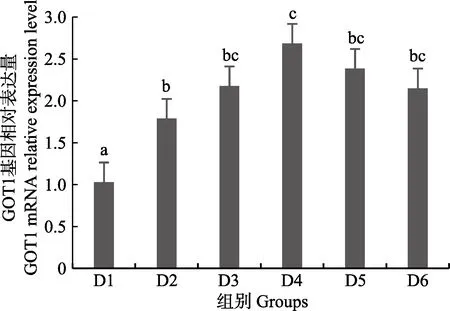
圖4 肝臟谷草轉氨酶1(GOT1)基因相對表達量Fig.4 The relative expression level of the glutamic oxalacetic transaminase1(GOT1) mRNA in liver
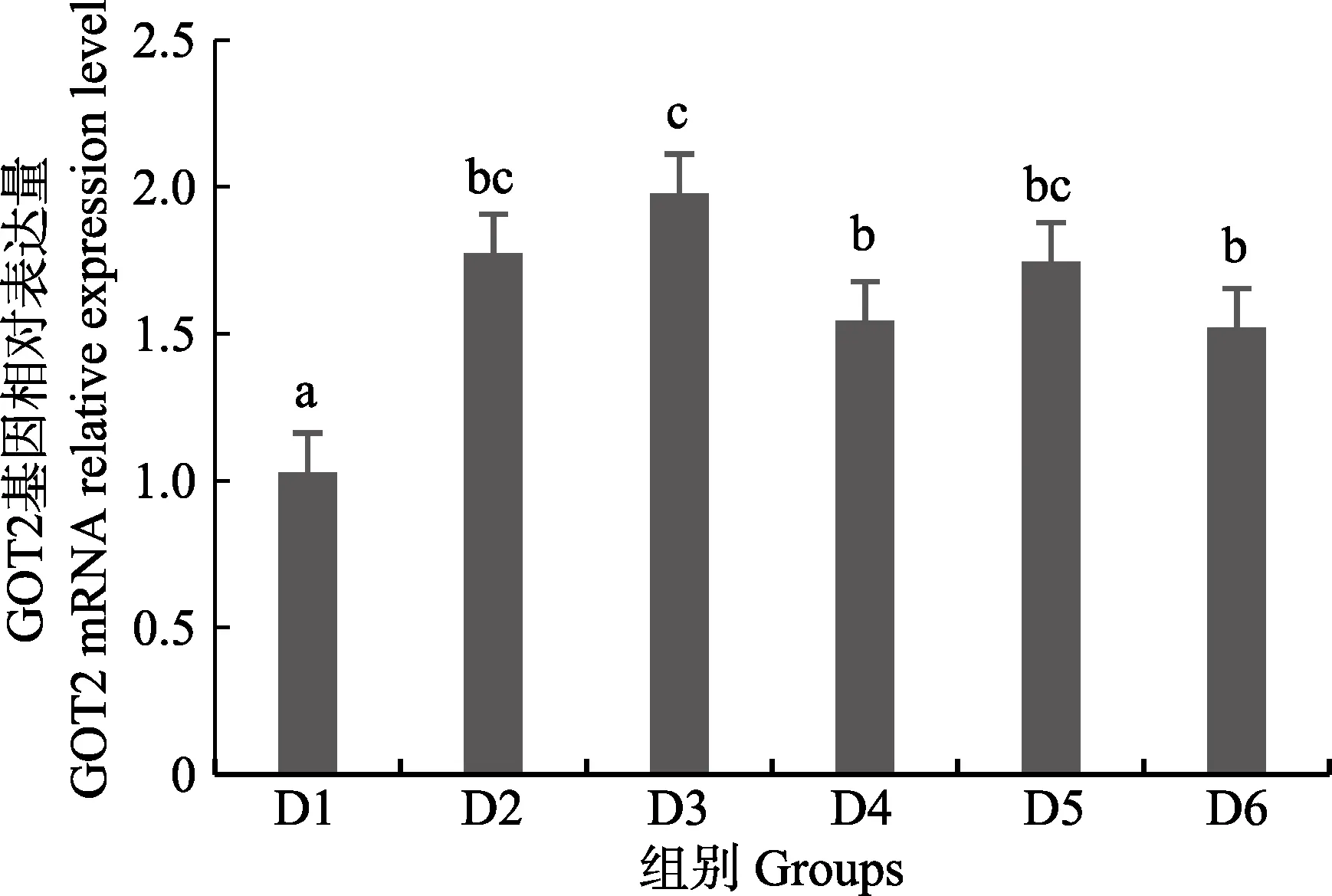
圖5 肝臟谷草轉氨酶2 (GOT2)基因相對表達量Fig.5 The relative expression level of the glutamic oxalacetic transaminase2(GOT2) mRNA in liver
3 討論
3.1 飼料中維生素B6含量對許氏平鲉幼魚生長的影響
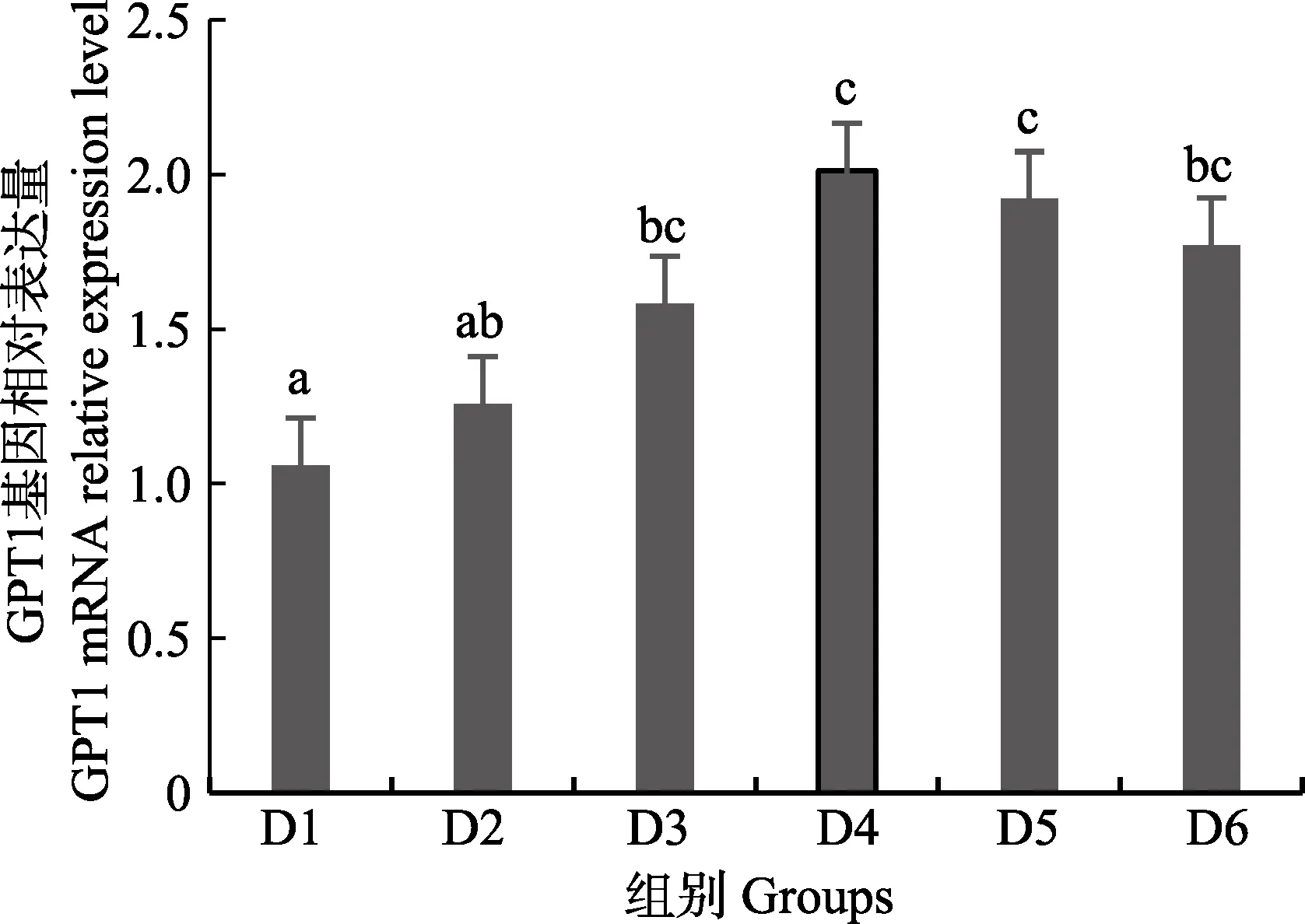
圖6 肝臟谷丙轉氨酶1(GPT1)基因相對表達量Fig.6 The relative expression level of the glutamic-pyruvic transaminase 1(GPT1) mRNA in liver
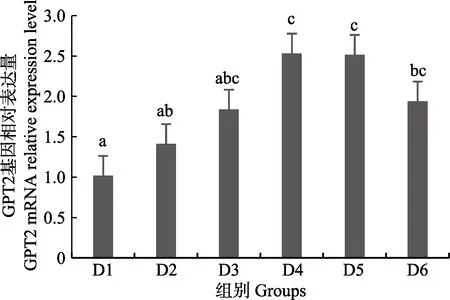
圖7 肝臟谷丙轉氨酶2(GPT2)基因相對表達量Fig.7 The relative expression level of the glutamic-pyruvic transaminase 2(GPT2) mRNA in liver
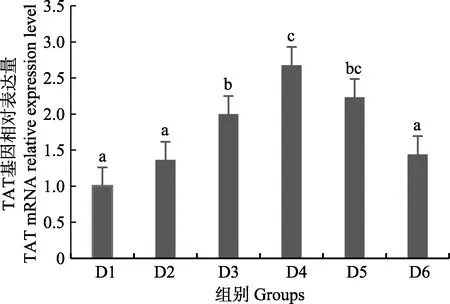
圖8 肝臟酪氨酸轉氨酶(TAT)基因相對表達量Fig.8 The relative expression level of the tyrosine aminotransferase (TAT) mRNA in liver
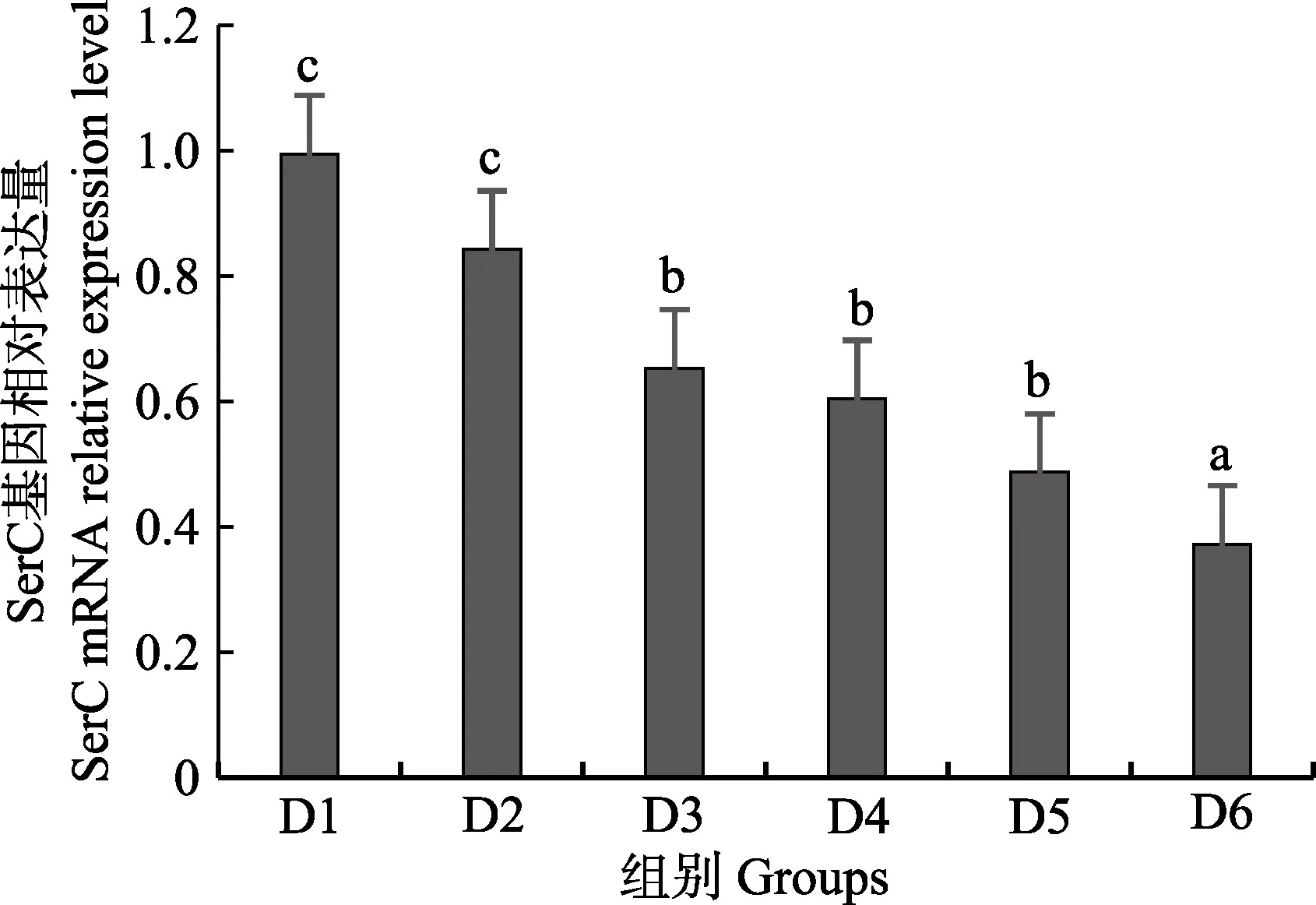
圖9 肝臟絲氨酸轉氨酶(SerC)基因相對表達量Fig.9 The relative expression level of the serine aminotransferase (SerC) mRNA in liver
實驗結束后,對照組的增重率和特定生長率均顯著低于其他組,且隨著維生素B6含量的增加呈先上升后下降的趨勢,說明維生素B6是維持許氏平鲉正常生長發育所必需的營養物質。維生素B6在多種代謝過程中起作用,它的缺乏可導致細胞蛋白質合成和轉化反應的錯位,在飼料中添加適量的維生素B6會提高蛋白質的利用率(Baker et al, 1995)。本研究中,添加維生素B6組的蛋白質效率顯著高于對照組,飼料系數呈先降后升的趨勢,飼料利用率有所提高。水產動物缺乏維生素 B6會出現相應的癥狀,如黃尾(Seriolalalandi)出現厭食,生長不良,貧血,神經紊亂和高死亡率的缺乏癥(Hosokawa et al, 1992);條石鯛(Oplegnathus fasciatus)的缺乏癥是生長緩慢,厭食,神經失調,游泳異常,高死亡率和貧血(Ishibashi et al, 2002)。實驗中對照組未出現常見的維生素B6缺乏癥,有可能因為實驗開始前,許氏平鲉幼魚體內積累了一部分維生素B6;養殖實驗在水泥池掛網箱中進行,水體中也有部分維生素B6的存在,而不能及時排出;攝食飼料中也添加了魚粉(含有一些維生素B6),因此,對照組缺乏癥不明顯,只出現了攝食不積極的癥狀,但存活率較其他組無顯著差異,說明2.08 mg/kg 的維生素B6可以防止許氏平鲉幼魚出現嚴重的缺乏癥。
本實驗以增重率、肝臟維生素B6含量和谷丙轉氨酶為評價指標,得出許氏平鲉幼魚維生素B6的需求量為3.53~6.32 mg/kg,與印度鯰(Heteropneustes fossilis)3.21 mg/kg(Mohamed, 2001)、紅羅非魚(Oreochromis mossambicus × O. niloticus) 3 mg/kg(Lim et al, 1995)、大黃魚(Pseudosciaena crocea) 4.49~4.61 mg/kg(張春曉,2006)、鱸魚(Lateolabrax japonicus) 4.38~4.53 mg/kg(張春曉, 2006)、團頭魴(Megalobrama amblycephala)4.17~ 5.02 mg/kg (王瑩等, 2013)的需求量類似;高于大菱鲆1~2.5 mg/kg(Adron et al, 1978)、斜帶石斑魚1.75~2.22 mg/kg(Huang et al, 2005)的需求量;低于翠鱧(Channa punctatus) 7.6~10.4 mg/kg(Zehra et al,2018)、黃尾2.2~11.7 mg/kg(Hosokawa et al, 1992)、吉富羅非魚(Oreochromis niloticus) 9.22~10.11 mg/kg(吳凡等, 2018)的需求量。不同實驗魚有不同需求量,原因可能與實驗魚的種類、大小、實驗環境、實驗飼料、飼喂頻率和方式、實驗方法和評估標準的有關。
魚體營養狀態可以通過形體指標來反映,如臟體比、肝體比和腸體比。研究表明,未添加維生素B6時,吉富羅非魚和異育銀鯽(Carassius auratas gibelio)肝體比顯著低于其他組(吳凡等, 2018; 王錦林等,2011);大西洋鮭(Salmo salar)10 mg/kg 組的肝體比顯著高于其他組(Albrektsen et al, 1994);建鯉(Cyprinus carpio)對照組的腸體比為最低值(He et al, 2009),本實驗的臟體比、肝體比和腸體比均呈先上升后下降的趨勢,對照組最低,說明維生素B6不足可能會影響許氏平鲉幼魚的器官發育能力。
3.2 飼料中維生素B6 對許氏平鲉幼魚魚體常規成分的影響
隨維生素B6的增加,許氏平鲉幼魚全魚粗蛋白、全魚和肌肉的粗脂肪呈先上升后下降的趨勢,肌肉粗蛋白含量先上升后趨于平穩,對照組肌肉水分顯著低于其他各組,這與吉富羅非魚(吳凡等, 2018)、異育銀鯽(王錦林等, 2011)的研究結果類似。維生素B6以5’-磷酸吡哆醛的形式作為轉氨酶的輔酶,參與氨基酸分解代謝的第一步去除α-氨基,使氨基酸轉化為相應的酮酸。維生素B6含量的增加會提高魚體蛋白利用率,增加機體蛋白的含量,促進蛋白的合成(Baker et al,1995; Giri et al, 1997);因此,全魚和肌肉的粗蛋白含量會隨維生素B6含量的增加而有所提高。此外,維生素B6在乙酰輔酶A 和脂肪酸利用等方面的脂質代謝中起重要作用。在草魚(Ctenopharyngodon idellus)和皺紋盤鮑(Haliotis discus hannai)中發現添加維生素B6可以調節和平衡多不飽和脂肪酸的代謝,有助于脂蛋白酯酶等的合成與釋放,促進了脂肪的運輸和利用,避免了脂質的沉積(馮秀妮, 2005; 黃世蕉等, 1992)。本研究中,對照組全魚和肌肉的粗脂肪含量顯著低于其他各組,隨著維生素B6的增加,D5、D6 組肌肉的粗脂肪和D6 組全魚的粗脂肪出現了下降,在印度鯪(Cirrhinus mrigala)的研究上也有類似結果(Zehra et al, 2017)。大西洋鮭未添加維生素B6組的全魚和肝臟脂質含量最低(Albrektsen et al, 1994)。以上結果說明,添加適量的維生素B6,能促進機體蛋白質的合成和脂質代謝反應的正常進行。
3.3 飼料中維生素B6對許氏平鲉幼魚肝臟維生素B6含量和抗氧化酶的影響
維生素B6在氨基酸代謝和含氮化合物的反應中充當氨基轉移酶和脫羧酶的輔酶,參與非必需氨基酸的合成(Baker et al, 1995)。魚類肝臟是維生素代謝和蓄積的重要器官,腸道吸收的水溶性維生素被運送到肝臟,以滿足新陳代謝的需要,肝臟中維生素B6的含量常作為特征性的評價指標(Zehra et al, 2017)。因此,測定肝臟中維生素B6的含量能更好的確定維生素B6的需求量。通過折線回歸模型,以肝臟維生素B6含量為評價指標,許氏平鲉幼魚維生素B6需求量為4.96 mg/kg。
維生素B6具有間接的抗氧化作用,可以清除機體超氧化物自由基并進一步預防氧化應激,而缺乏維生素B6會增加自由基的生成、降低抗氧化酶活性(Kumar et al, 2017)。抗氧化物酶作為魚體抗氧化防御的重要組成部分,其中,SOD 是一類抗氧化酶,從細胞環境中清除具有破壞性的活性氧(ROS),具有清除體內自由基的能力,抑制超氧化物的生成,在機體的防御系統和免疫系統中發揮著重要的作用(胡凱,2008; Cui et al, 2016)。MDA 是脂質過氧化的終產物,含量可反映機體細胞受自由基攻擊的嚴重程度(Mccord, 1969)。在斜帶石斑魚、凡納濱對蝦(Litopenaeus vannamei)的研究中發現,維生素B6缺乏組SOD 活性顯著降低(Huang et al, 2005; 黃曉玲,2014),與本實驗結果相似,隨著飼料維生素B6含量的增加,SOD 活性呈先上升后下降的趨勢,MDA 呈先下降后上升的趨勢,均在D3 組出現極值,這表明添加適當含量的維生素B6可以提高機體的抗氧化能力。
3.4 飼料中維生素B6 對許氏平鲉幼魚肝臟轉氨酶活力及其基因表達量的影響
維生素B6以5’-磷酸吡哆醛的形式,作為轉氨酶參與氨基酸代謝的標志性輔酶(Zehra et al, 2018),肝臟作為氨基酸代謝的主要場所,肝臟的谷草轉氨酶和谷丙轉氨酶常被用作評價魚體營養狀況的敏感指標,其活力間接反映蛋白質合成效率,同時,也是反映肝功能是否正常的重要指標(陳壯等, 2014; 馬俊等,2016; Smith et al, 1974)。故測定肝臟中谷草轉氨酶和谷丙轉氨酶的活力可以更準確得出維生素B6的需求量。本研究表明,對照組肝臟維生素B6含量和轉氨酶活力均顯著低于其他組,隨著維生素B6含量的增加,肝臟維生素B6含量呈先上升后平穩的趨勢;谷草轉氨酶呈上升的趨勢,但在D3~D6 組無顯著差異;谷丙轉氨酶呈先上升后下降的趨勢。這與吉富羅非魚、異育銀鯽和印度鯰的研究結果類似(Mohamed,2001; 吳凡等, 2018; 王錦林等, 2011),說明添加維生素B6可以提高轉氨酶的活力,保證魚體蛋白質代謝的正常進行。通過折線回歸模型,以谷丙轉氨酶為評價指標,許氏平鲉幼魚維生素B6需求量為6.32 mg/kg。
GOT 在真核細胞中存在2 種遺傳上不同的同工酶,細胞質和線粒體蛋白分別由GOT1 和GOT2 基因編碼(Jiang et al, 2015);GPT 基因也常分為2 個亞型,即肝細胞質中的谷丙轉氨酶1(GPT1)和位于肝線粒體中的谷丙轉氨酶2(GPT2)(荊曉麗, 2010)。目前,在水產動物上尚無關于維生素B6對轉氨酶基因表達水平影響的報道,但研究發現,維生素B6缺乏的大鼠(Rattus norvegicus)肝臟細胞質的谷草轉氨酶mRNA水平比對照組高7 倍,施用皮質醇可以誘導肝谷草轉氨酶 mRNA 的表達,而給予維生素B6會抑制由皮質醇誘導的谷草轉氨酶的合成;缺乏維生素B6會增強糖皮質激素受體的DNA 結合活性,提高谷草轉氨酶的基因表達(Kondo et al, 1985; Oka et al, 1995)。研究表明,維生素B6缺乏不但降低了肝臟丙氨酸-乙醛酸轉氨酶活性,而且還下調了肝細胞丙氨酸-乙醛酸轉氨酶的基因表達(Nishijima et al, 2003)。給大鼠喂食缺乏維生素B6的飼料后,丙氨酸-乙醛酸轉氨酶1 活性的降低比丙氨酸-乙醛酸轉氨酶2 活性(在14 d 內降低33%相比86%)更快速和顯著(Takada et al, 1984)。5’-磷酸吡哆醛(PLP)作為糖皮質激素受體的體內調節劑,PLP 水平變化會影響糖皮質激素受體的DNA 結合位點,并最終影響酪氨酸轉氨酶(TAT)的活性(Disorbo et al, 1981)。絲氨酸轉氨酶(SerC)參與氨基酸合成通路的表達,使谷氨酸轉氨基生成3-磷酸絲氨酸,SerC 基因作為調控和關鍵酶參與L-絲氨酸的合成(劉浩等, 2015),維生素B6作為所有轉氨酶的輔酶,也會影響絲氨酸轉氨酶基因的表達水平。
本研究中,隨著飼料中維生素B6含量的升高,GOT1 的mRNA 基因表達呈先上升后下降的趨勢,對照組的基因表達量顯著低于其他各組,D3~D6 組間沒有顯著差異;GOT2 的對照組基因表達水平最低,D3 組高表達,以上結果表明,添加維生素B6會提高谷草轉氨酶的基因表達。GPT1 和GPT2 的mRNA 基因表達水平呈相似的趨勢,均是先升高后降低,且在D4 組表達量最高,對照組表達量最低,由此可見,維生素B6缺乏會降低谷丙轉氨酶的基因表達,適宜的維生素B6添加會使其高表達。同時,TAT 的基因表達水平也是對照組最低,在D3~D5 組表達量顯著高于對照組、D2 和D6 組。SerC 基因表達水平呈下降趨勢,對照組和D2 組表達量顯著高于D3~D6 組。本研究結果與以上結論有差異的原因可能是物種、實驗條件和方法、實驗形式等所致。
隨著維生素B6的添加,谷草轉氨酶活力呈上升趨勢,谷丙轉氨酶活力先升后降,這與mRNA 表達水平趨勢類似,說明適宜含量的維生素B6能促進谷草轉氨酶和谷丙轉氨酶基因的轉錄,提高谷草轉氨酶和谷丙轉氨酶的活力,促進魚體蛋白質合成效率和利用率,更好的調控許氏平鲉幼魚的蛋白質代謝。此外,添加維生素B6會上調酪氨酸轉氨酶基因的表達量,下調絲氨酸轉氨酶基因的表達量,具體作用機制需要進一步的實驗來證實。
4 結論
綜上,維生素B6是許氏平鲉幼魚生長發育和維持正常生理功能的必需營養物質。適宜含量的維生素B6可以提高許氏平鲉幼魚抗氧化能力和代謝酶活性,提高谷草轉氨酶、谷丙轉氨酶和酪氨酸的基因表達水平,降低絲氨酸轉氨酶的基因表達水平。以增重率、肝臟維生素B6含量和谷丙轉氨酶為評價指標,36 g 的許氏平鲉幼魚維生素B6的需求量為3.53~6.32 mg/kg 飼料。
