社群性別結構對遂昌牛頭山白鷴活動節律的影響
2020-10-28 09:00:12龔笑飛程曉云潘江炎樊曉麗林植華
麗水學院學報 2020年5期
龔笑飛,程曉云,曹 華,潘江炎,樊曉麗,林植華*
(1.遂昌縣林業發展中心,浙江遂昌323300;2.麗水學院生態學院,浙江麗水323000)
活動節律是動物應對周圍環境的長期適應的結果[1]。在特定條件下,動物可依據非生物因素和生物因素的變化對自身行為進行調節以形成特定的活動節律[2-5]。非生物因素,例如溫度、降雨量和光照水平,可能會影響食物供應、熱調節和內源性節律,從而導致活動節律的變化[6-10]。生物因素,如物種內和物種間的相互作用、動物集群活動及性別等會對活動節律產生影響[11-13]。社群活動是鳥類的一種普遍且重要的行為適應,該行為可減小自身被捕食幾率及增加覓食率[14-15]。因此,通過比較不同的社群性別結構下純雌鳥群、純雄鳥群與雌雄混群的活動節律差異,有助于了解社群性別結構與活動節律之間的關系、分析物種的生存狀況及適應策略、探討動物生態行為策略和適應策略。
白鷴(Lophura nycthemera)隸屬于雞形目(Galliformes)雉科(Phasianidae),屬地棲性鳥類,為國家二級重點保護野生動物[16],主要分布于我國南方各省[16-17]。該物種具有多樣的社群性別組成方式,常集群活動[15,18]。
紅外相機技術作為一種非損傷性的調查方法,因具有不受調查時間和人力的限制、可獲取全天候的數據等優點,而被廣泛應用于野生動物活動模式和活動節律的研究[19-20]。近些年,國內學者成功利用紅外相機技術對某些珍稀瀕危物種的野外行為生態進行了深入的研究[21-22]。目前,基于紅外相機技術,已在戴云山山系[23]、弄崗國家級自然保護區[15,24]、古田山國家級自然保護區[18]與鳳陽山-百山祖國家級自然保護區[25]等地開展了關于白鷴的活動節律研究。但是,關于白鷴社群結構與其活動節律之間關系的研究則相對較少。為此,本文通過在遂昌牛頭山進行的運用紅外相機的調查,旨在:(1)掌握該區域中不同的社群性別結構白鷴的活動節律;(2)分析在不同的社群性別結構下白鷴的活動節律差異;(3)探討不同社群結構下白鷴的行為生態策略。
1 研究區域與方法
1.1 研究區域
牛頭山屬中亞熱帶濕潤季風區,地處武夷山系仙霞嶺山脈分支。年總降水量為1 800~2 400 mm,年平均氣溫在10~16℃之間[26]。林場全境植物群落區系組成多樣,珍稀野生動物種類繁多,包括黑麂(Muntiacus crinifrons)、鬣羚(Capricornis milneedwardsii)、獼猴(Macaca mulatta)、黃腹角雉(Tragop ancaboti)、白鷴(數據未發表)等。
1.2 紅外相機數據獲取
2018年1月—2019年1月,在遂昌牛頭山林場選擇57個研究位點,涵蓋6個林區。每個研究位點布設1臺Ltlacorn紅外相機。采用樣線法進行紅外相機布設,相機間的距離不小于200 m。在監測期間,每3個月對相機的SD卡和電池進行檢查與更新。紅外相機工作模式設置為中等靈敏度,連拍3張后錄制10 s視頻,每張照片間隔1 s等。選擇具有明顯動物活動痕跡的大型喬灌木進行相機的固定,固定高度為0.5~0.75 m,記錄每臺相機的經緯度、海拔、植被類型等信息。
1.3 紅外相機數據讀取與分析
參照薛亞東等[27]的方法提取獨立有效照片,以降低同一物種照片的自相關性;參考《浙江動物志:鳥類》[16]進行白鷴照片的物種及性別的鑒定,利用錄制視頻來輔助鑒定。每張獨立有效照片視為一次有效探測,統計每張獨立有效照片中的雄鳥與雌鳥的個體數量,并匯總該照片的拍攝時間、環境溫度和相機位點等信息,用于白鷴日活動節律和季節活動節律的分析。
1.4 數據分析
全部數據用SPSS18.0進行分析。用G檢驗對相應數據進行統計分析。所有統計值用平均值±標準誤表示,顯著性水平設置為0.05。
2 結果
研究期間,遂昌牛頭山的紅外相機有效工作日為13 655個,獲得紅外相機視頻與照片文件約89 176份。其中,共收集到白鷴照片數1 631張,提取有效探測次數為366次。調查記錄白鷴548只次,成體及亞成體雄性218只次,雌性330只次,成體及亞成體的雌雄性比為1.51:1。記錄到白鷴群體組成模式有雌雄混群(44次)、純雌鳥群(161次)、純雄鳥群(161次)。
2.1 白鷴日活動節律的社群性別差異
白鷴在不同時間段的活動頻率存在極顯著差異(G=380.16,df=11,P< 0.01),不同社群性別結構的白鷴種群間無顯著差異(G=14.67,df=22,P>0.05)。白鷴具有3個日活動高峰,分別在06:00—08:00、12:00—14:00及16:00—18:00時間段內(圖1)。
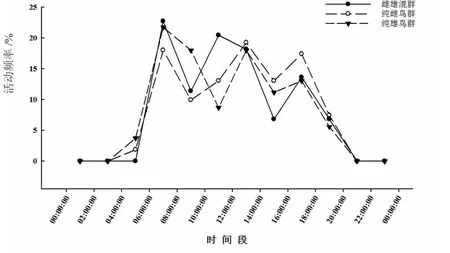
圖1 不同社群結構白鷴的日活動節律
2.2 白鷴季節活動節律的社群性別差異
白鷴在不同季節的活動頻率存在極顯著差異(G=62.55,df=3,P<0.01),主要集中于春季。社群性別結構不同會使白鷴季節活動頻率產生差異(G=20.63,df=6,P<0.05),表現為純雌鳥群與純雄鳥群的季節活動頻率受季節影響(all P<0.01)。純雌鳥群季節活動高峰期為秋季;純雄鳥群季節活動高峰期為春季。雌雄混群的季節活動頻率則不受季節影響(G=5.62,df=3,P>0.05)。如圖 2。
2.3 白鷴溫度范圍活動節律的社群性別差異
白鷴在不同的溫度范圍內的活動頻率存在極顯著差異(G=123.92,df=6,P<0.001)。不同社群性別結構白鷴的溫度活動頻次分布均呈偏峰分布,其活動高峰均在22.5℃左右。不同社群性別結構的白鷴種群間無顯著差異(G=9.35,df=12,P>0.05)。如圖3。

圖2 不同社群結構白鷴的季節活動節律
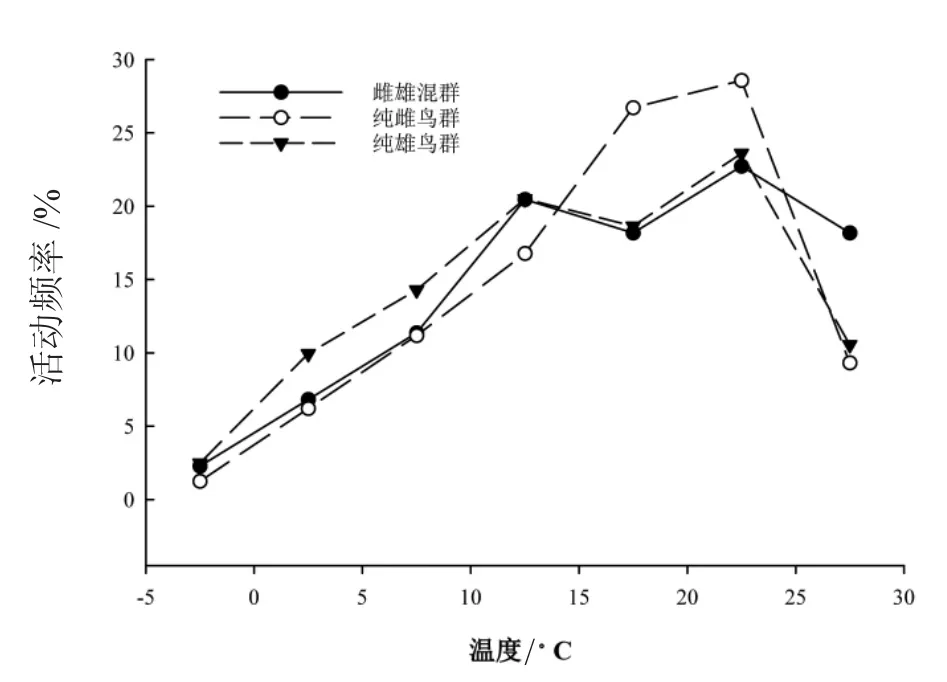
圖3 不同社群性別結構白鷴的溫度活動節律
2.4 白鷴植被類型選擇的社群差異
白鷴在不同植被類型下活動頻次存在顯著差異(G=212.96,df=3,P<0.01),而不同社群性別結構的白鷴種群間無顯著差異(G=8.89,df=6,P>0.05)。當植被類型為常綠闊葉林時,不同社群性別結構的白鷴活動頻率均為最大值(圖4)。

圖4 不同社群性別結構白鷴的植被類型活動節律
3 討論
本研究采用紅外相機技術對遂昌牛頭山林場內白鷴的活動節律進行研究。結果顯示:遂昌牛頭山林場內白鷴為日行性鳥類,純雌鳥群、純雄鳥群與雌雄混群是白鷴的3種集群方式,且單性集群為主要方式,這與白冠長尾雉(Syrmaticus reevesii)的研究結果類似[28]。該物種具有3個活動高峰期,這與廣西弄崗國家級自然保護區內白鷴的日活動模式相似[24]。與直接觀察法相比[29],在06:00—08:00及16:00—18:00均為白鷴活動高峰期外,本研究還發現12:00—14:00為白鷴活動高峰期。這與劉佳等[15]在喀斯特生境中的研究有差異,而導致此差異的原因可能是兩地海拔和氣候不同。不同社群性別結構的白鷴,其活動高峰并不完全一致,具有一定程度的錯峰現象。這一現象可能是為獲得最大的廣義適合度,減小不同社群間的競爭。
春季是白鷴的繁殖期[16]。處于繁殖期的雄鳥運動行為會顯著增加(如開始追逐雌鳥,來回走動),以更多地展示自己,尋求配偶,繁殖期過后雄鳥離群;而雌鳥在繁殖期與非繁殖期間的行為頻次沒有差異[30-31]。這種現象與本研究中純雄鳥群季節活動高峰為春季的結果一致。繁殖期后,雌鳥在夏季開始離群,入巢孵卵[32],此時雌鳥活動頻率低且不易被拍攝到,導致純雌鳥群的夏季活動頻次較春季低。秋季,雌雄混群和純雌鳥群的活動頻次最高可能是由于在秋季結群重新形成,雌鳥與雛鳥回到群內[32],加之食物豐富,白鷴增加活動頻次以攝取足量的能量。冬季氣溫低,在低溫環境中活動,會耗費過多的能量[18],同樣,過高的環境溫度也會影響動物的生存活動[33]。3種社群性別結構的白鷴均在一定程度內減少了在夏冬兩季的活動頻次。因此,我們推測,白鷴集群行為的季節性變化可能是導致不同社群性別結構的種群間季節活動節律存在差異的原因之一。
動物一天中的活動高峰發生在最適合其活動的環境溫度時段[34]。紅外相機是根據被拍攝對象的熱輻射與周圍環境的熱量差異而觸發相機拍攝的,其溫度傳感裝置較為敏感[35]。因此,可基于紅外相機所記錄的環境溫度來研究動物活動最適宜的參考溫度范圍[36]。研究結果顯示,20~25℃的溫度范圍為白鷴的活動高峰。因此,20~25℃為白鷴活動的最適宜環境溫度范圍。同時,基于限制相似性和競爭排斥的概念,因為性別間的種內競爭,同一物種的兩性間可能在資源的時間利用上表現出差異[37-39]。研究發現,純雌鳥群與純雄鳥群的次活動高峰的溫度選擇存在一定程度的差異。因此,不同社群間次活動高峰的溫度差異可能是白鷴社群間生態位分化的策略之一。
白鷴偏好棲息在常綠闊葉林。這與熊志斌等[40]研究結果一致。在亞熱帶和溫帶森林中,殼斗科植物常成為重要的建群種和山地常綠闊葉林的主要上層樹種[41]。白鷴雖為雜食性鳥,但其食物中動物性食物含量很少,植物性食物含量占絕大多數,以殼斗科果實的取食頻度與比例最高[42-43]。巢址選擇是鳥類防御天敵、提高繁殖成功率的重要生活史策略[44],其需滿足保證自身和后代的安全性和為孵卵提供適合的微氣候[45]。張娟茹等[46]通過研究白鷴的巢址選擇發現,白鷴偏好在郁閉度高的地方筑巢;邵晨等[47]對白鷴夜棲息所選擇的研究結果顯示,白鷴夜棲息所多選擇高大的常綠喬木,如青岡櫟(Cyclobalanopsis glauca)、苦櫧(Castanopsis sclerophylla)、甜櫧(C.eyrei)、樟樹(Cinnamomum camphora)等。在遂昌牛頭山林場,常綠闊葉林相較于其他植被類型具有更高的郁閉度且殼斗科植物、青岡櫟、樟樹等高大的常綠喬木常存在于常綠闊葉林中。因此,在遂昌牛頭山區域中,白鷴的這種生境類型偏好選擇,可獲得較為豐富的食物、良好的巢址和夜棲息所,從而使自己的生境適合度達到最大[48]。不同社群性別結構的白鷴種群間無顯著差異,可能是因為不同社群性別結構之間對食物、巢址和夜棲息所的選擇無明顯差異。
綜上所述,本研究得到以下4個主要結論:(1)不同社群性別結構的白鷴種群在3個活動高峰期存在一定程度的錯峰現象;(2)白鷴集群行為的季節性變化可能是導致不同社群性別結構的種群間季節活動節律存在差異的原因之一;(3)白鷴活動最適宜環境溫度范圍為20~25℃,純雌鳥群與純雄鳥群的次活動高峰的溫度選擇存在一定程度的差異;(4)白鷴偏好棲息在常綠闊葉林中,偏好在春季活動,不同社群性別結構之間差異不顯著。
猜你喜歡
少先隊活動(2022年5期)2022-06-06 03:45:04
音樂探索(2022年2期)2022-05-30 21:01:37
家庭科學·新健康(2022年3期)2022-05-10 00:32:13
中老年保健(2021年2期)2021-08-22 07:31:10
少先隊活動(2021年1期)2021-03-29 05:26:36
快樂語文(2020年30期)2021-01-14 01:05:38
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
海峽姐妹(2018年3期)2018-05-09 08:20:40
