嬰兒源雙歧桿菌對人胎結腸上皮細胞的增殖作用及機制研究
2020-10-23 11:25:10李欣芮范小飄高文文尚佳萃孟祥晨
食品工業科技 2020年20期
趙 桉,李欣芮,范小飄,高文文,尚佳萃,萬 峰,孟祥晨
(東北農業大學,乳品科學教育部重點實驗室,黑龍江哈爾濱 150030)
雙歧桿菌是生命早期最為重要的腸道優勢菌群之一[1],具有促進腸道發育[2]、調節免疫系統[3]、維持組織穩態[4]、促進新陳代謝等生理學功能[5]。由于嬰兒腸道發育完善程度影響了嬰兒食物消化、營養吸收、免疫系統增強等生理學功能[2],因此雙歧桿菌促進新生兒腸道發育成為近年來的研究熱點。
Che等[6]用人體內的雙歧桿菌處理無菌豬,建立了HFA(Human flora-associated)豬模型,在菌株定植腸道后,空腸上皮絨毛長度增長、隱窩加深,影響腸道發育,但對其中起作用的菌體成分及作用機制很大程度上未知。目前已發現雙歧桿菌的短鏈脂肪酸(SCFAs)[7]、乳酸[8]以及短雙歧桿菌的菌毛[9]可促進新生兒腸道發育。其他菌體成分如脂磷壁酸(LTA)[10]、胞外多糖(EPS)[11]、蛋白質[12]等物質在完善嬰兒其他的腸道功能中起到了積極作用。其中的蛋白質主要包括分布于菌體上的表面蛋白和菌體外的分泌蛋白,是雙歧桿菌與宿主相互作用和分子間交流的潛在介質,直接參與感知環境因素、促進細菌定植、介導與腸粘膜相關免疫細胞的產生[13]。但是表面蛋白和分泌蛋白是否參與了腸上皮細胞的增殖,尚無研究報道。因此,本實驗室篩選出一株對腸上皮細胞增殖促進作用較好的菌株,探究其中的蛋白成分對細胞增殖的影響。
研究發現Wnt/β-catenin信號通路的關鍵基因與哺乳動物腸上皮細胞增殖密切相關[14-15]。β-catenin、Cyclin D1、c-Myc 是細胞增殖正調控基因;Axin2、GSK-3β是細胞增殖的負調控基因[16]。因此選取Wnt/β-catenin 信號通路正負調控基因,檢測其表達變化情況,可在mRNA水平上檢測雙歧桿菌分泌蛋白和表面蛋白對腸上皮細胞增殖的影響[17]。
與以往研究新生兒腸道發育問題時,常選用動物腸細胞系或人腸癌細胞系不同,本試驗直接選用妊娠23周人胎結腸上皮CCD841 CoN細胞作為受試細胞,從12株雙歧桿菌(3~6月齡健康嬰兒糞便分離鑒定[18])中篩選出1株對CCD841 CoN細胞增殖促進作用最好的菌株,分析該菌株的表面蛋白和分泌蛋白的作用,初步探討雙歧桿菌對腸上皮細胞增殖影響的理論機制,為新生兒腸道發育以及腸道健康研究奠定基礎。
1 材料與方法
1.1 材料與儀器
12株雙歧桿菌(2株動物雙歧桿菌乳酸亞種:H6-7、H15-7;2株短雙歧桿菌:H9-3、H4-2;2株雙歧桿菌:H3-R2、H4-3;2株長雙歧桿菌嬰兒亞種:H10-1、H11-7;4株長雙歧桿菌長亞種:H4-1、H11-2、H16-3、H21-7) 分離于東北地區一月齡健康嬰兒(男嬰11人、女嬰10人)糞便,保藏于實驗室;人胎結腸上皮CCD841 CoN細胞 北納生物;改良MRS(mMRS)培養基 MRS培養基中添加0.05% L-半胱氨酸鹽酸鹽;高糖培養基DMEM 美國HyClone;胰蛋白酶 美國Sigma公司;胎牛血清 加拿大Wisent公司;0.25% Trypsin-EDTA 美國Gibco公司;與細胞培養相關的產品 美國Corning公司;CCK-8細胞增殖-毒性檢測試劑盒 美國AbMole公司;細胞周期染色試劑盒 聯科生物技術股份有限公司;細菌總 RNA 提取試劑盒、逆轉錄試劑盒、RT-PCR反應試劑盒(SYBR Green) 天根公司;引物 吉林省庫美生物科技有限公司;其他試劑 國產生化分析純產品。
厭氧培養箱 美國賽默飛公司;HF90型二氧化碳培養箱 力康發展有限公司;BA300型倒置顯微鏡 麥克奧迪實業集團有限公司;Model 680型酶標儀 美國Bio-Rad公司;Cytomics FC 500流式細胞儀 美國Beckman Coulter公司;Step One Plus實時熒光定量PCR儀 美國Applied Biosystem公司;核酸蛋白測定儀 美國Molecular Devices公司。
1.2 實驗方法
1.2.1 細胞培養 CCD841 CoN細胞培養于添加10%胎牛血清的DMEM混合培養基中,置于37 ℃、5%CO2條件下培養,胰酶消化傳代。
1.2.2 雙歧桿菌活化及培養 12株供試菌株在試驗前需進行復蘇活化,3%接種于mMRS 液體培養基中,37 ℃厭氧培養16 h,連續傳2~3代以恢復菌種活力。平板菌落計數出活菌數量后,調整菌體濃度為3×107CFU/mL以備用。
1.2.3 雙歧桿菌的篩選
1.2.3.1 雙歧桿菌發酵上清液和破碎物的制備 發酵上清液的制備:取厭氧培養16 h后的菌液10 mL,8000 r/min 4 ℃離心5 min,收集上清液,調節pH7.2,過0.22 μm濾器除菌,-20 ℃保存[19]。
破碎物的制備:取厭氧培養16 h后的菌液10 mL,8000 r/min 4 ℃離心5 min收集菌體。用細胞培養洗滌2遍并將菌體懸于10 mL細胞培養基中。用超聲波細胞粉碎儀在500 W條件下,每處理10 s、間隔10 s,超聲15 min。在顯微鏡下觀察沒有完整菌體后,過0.22 μm濾器除菌,-20 ℃保存[20]。
1.2.3.2 細胞增殖實驗分組 發酵上清組:加入含10%(V/V)發酵上清的細胞培養基;陰性對照組:加入含10%(V/V) mMRS液體培養基的細胞培養基;陽性對照組:加入含100 μg/mL小牛血清蛋白(BSA)[21]的細胞培養基;空白組:空孔加入細胞培養基。
破碎物組:加入含10%(V/V)破碎物的細胞培養基;陰性對照組:加入細胞培養基;陽性對照組:加入含100 μg/mL BSA的細胞培養基;空白組:空孔加入細胞培養基。
1.2.3.3 細胞增殖活性分析 細胞增殖活性檢測采用CCK-8法[22]。調整CCD841 CoN細胞密度為3×104cell/mL,接種于96孔板,每孔100 μL,培養24 h貼壁待用。棄上清,分別加入12株雙歧桿菌的發酵上清組和破碎物組,及其陰性對照組、陽性對照和空白組,每孔體積100 μL,設4個復孔,與細胞共作24 h。作用結束后,每孔加入10 μL CCK-8溶液,37 ℃孵育2 h后振蕩混勻,在酶標儀上檢測各孔光密度(OD)值,檢測波長設置為450 nm。按下列公式計算細胞增殖活性。
細胞增殖活性(%)=(實驗組OD值-空白組OD值)/(陰性組OD值-空白組OD值)×100
1.2.4 菌株H3-R2表面蛋白和分泌蛋白對CCD841 CoN細胞的增殖影響
1.2.4.1 雙歧桿菌H3-R2表面蛋白和分泌蛋白的制備 表面蛋白的制備:參照文獻[23]的方法并略做修改。取1 L于37 ℃厭氧培養16 h的雙歧桿菌H3-R2培養物,9000 r/min 4 ℃離心15 min收集沉淀,用PBS(pH7.2)洗滌2次。
在菌體中加入50 mL 5 mol/L LiCl溶液,在37 ℃于搖床(200 r/min)中處理30 min。9000 r/min 4 ℃離心15 min收集上清,過0.22 μm濾膜,收集濾液裝入8000 Da截留分子量的透析袋,在去離子水中透析48 h(4 ℃)后真空冷凍干燥,-20 ℃保存。
分泌蛋白的制備:參照文獻[24]的方法并略做修改。取1 L于37 ℃厭氧培養16 h的雙歧桿菌H3-R2培養物,9000 r/min 4 ℃ 離心15 min收集上清液,過0.22 μm濾膜除去菌體。向上清中加入三氯乙酸溶液,調整終濃度為10%,4 ℃靜止2 h,9000 r/min離心20 min,收集沉淀,用冷丙酮洗滌2次,真空冷凍干燥,-20 ℃保存。
1.2.4.2 表面蛋白和分泌蛋白的SDS-PAGE電泳 SDS-PAGE:分別取1 mg表面蛋白和分泌蛋白粉末溶解于100 μL 1% SDS溶液中,加入等量的2×SDS-PAGE緩沖液,沸水浴10 min,取10 μL上樣,進行SDS-PAGE電泳。SDS-PAGE凝膠:12%分離膠、5%濃縮膠。電泳電壓:分離膠120 V,濃縮膠80 V。用考馬斯亮藍R250對凝膠染色4 h后出現明顯條帶,甲醇/醋酸脫色液脫色6 h后背景變淺、條帶清晰。凝膠成像系統拍照分析電泳結果。
1.2.4.3 細胞增殖實驗分組 表面蛋白組和分泌蛋白組:加入含表面/分泌蛋白濃度為100、50、25、10、5、1 μg/mL的細胞培養基;陽性對照組:加入含100 μg/mL BSA的細胞培養基;陰性對照組:加入細胞培養基;空白組:空孔加入細胞培養基。
1.2.4.4 細胞增殖活性分析 調整CCD841 CoN細胞密度為3×104cell/mL,接種于96孔板,每孔100 μL,培養24 h貼壁待用。棄上清,加入表面蛋白組和分泌蛋白組,及其陰性對照組、陽性對照和空白組,每孔體積100 μL,設4個復孔,與細胞共作12、24、48 h。作用結束后,測量細胞增殖活性同1.2.3.3。
1.2.4.5 細胞周期、熒光定量PCR實驗分組 表面蛋白組:加入含表面蛋白濃度為50 μg/mL的細胞培養基;分泌蛋白組:加入含分泌蛋白濃度為10 μg/mL的細胞培養基;對照組:加入細胞培養基。
1.2.4.6 細胞周期檢測 調整CCD841 CoN細胞密度為2×105cell/mL,接種于6 cm培養皿,每孔4 mL,培養24 h后貼壁待用。棄上清,分別加入表面蛋白組、分泌蛋白組以及對照組與細胞共作24 h。用胰蛋白酶消化細胞,1000 r/min離心5 min 收集細胞,棄去上清。用PBS洗滌1次,1000 r/min離心5 min,棄去上清。加入1 mL碘化丙啶染色工作液和10 μL破膜劑,振蕩混勻后室溫避光孵育30 min,避免振動,上流式細胞儀檢測[25]。
1.2.4.7 熒光定量PCR檢測基因表達水平 收集1.2.4.6中表面蛋白組、分泌蛋白組以及對照組與CCD841 CoN細胞共作24 h后的細胞,用試劑盒提取細胞總RNA,核酸蛋白測定儀測定RNA濃度及純度,符合A260/A280=1.8~2.1后用于逆轉錄反應。
用逆轉錄試劑盒合成cDNA,反應體系20 μL,反應條件:37 ℃ 15 min,85 ℃ 5 s。
用RT-PCR試劑盒進行熒光定量PCR,反應體系25 μL,反應條件:95 ℃預變性30 min,95 ℃變性5 s,60 ℃退火30 s,95 ℃延伸15 s,共40個循環,60~95 ℃記錄熔解曲線,引物序列設計見表1。在熒光定量PCR儀器上進行反應,并記錄樣本循環閾值(Ct值)。以2-ΔΔCt相對定量法計算基因表達量的變化。

表1 熒光定量PCR引物序列
1.3 數據處理
采用Excel 2010軟件處理試驗數據,結果以Means±SD表示。采用Origin 8.0軟件繪圖及擬合處理。運用SPSS 23.0軟件,單因素方差分析以及Duncan多重比較分析,P<0.05有統計學意義。
2 結果與分析
2.1 雙歧桿菌對CCD841 CoN細胞的增殖促進作用
2.1.1 雙歧桿菌發酵上清的作用 細胞增殖活性百分比可以表示細胞增殖作用的強弱,其值大于100% 時表示有增殖促進作用。12株菌中有7株菌(H4-1、H21-7、H6-7、H3-R2、H4-3、H10-1、H11-7)的發酵上清對CCD841 CoN細胞有增殖促進作用(圖1)。與陰性對照組相比,添加菌株H4-1、H21-7、H6-7、H3-R2、H4-3、H10-1、H11-7的發酵上清液與細胞共作24 h的細胞增殖活性分別為:134%、102%、123%、137%、111%、114%、102%。并且添加菌株H4-1、H3-R2的發酵上清液與細胞共作24 h的增殖促進效果均顯著高于陽性對照組(P<0.05)。
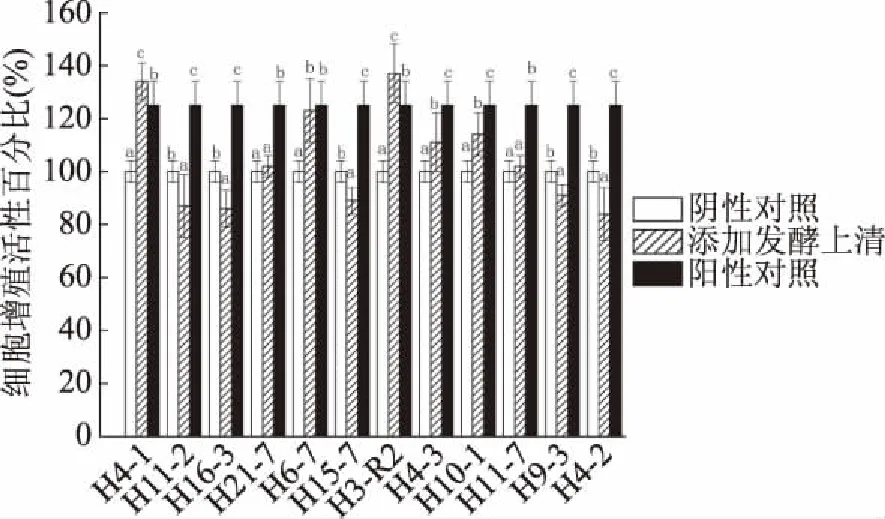
圖1 12株雙歧桿菌的發酵上清液對CCD841 CoN細胞的增殖促進作用
2.1.2 雙歧桿菌破碎物的增殖作用 12株菌中有7株菌(H21-7、H6-7、H15-7、H3-R2、H4-3、H9-3、H4-2)的細胞破碎物對CCD841 CoN細胞有增殖促進作用(圖2)。與陰性對照組相比,添加菌株H21-7、H6-7、H15-7、H3-R2、H9-3、H4-2的破碎物與細胞共作24 h的細胞增殖活性分別為:124%、120%、140%、153%、110%、122%、157%。并且添加菌株H15-7、H3-R2、H4-2的破碎物與細胞共作24 h的增殖促進效果均顯著高于陽性對照組(P<0.05)。
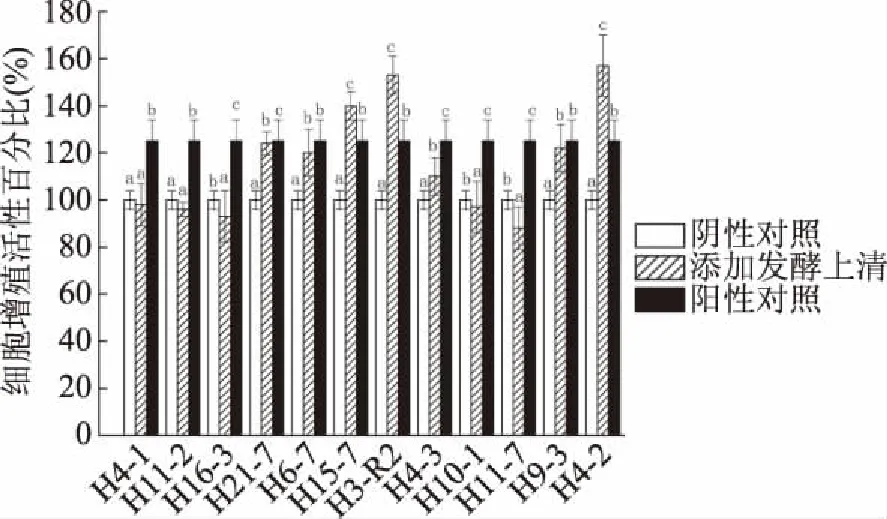
圖2 12株雙歧桿菌的破碎物對CCD841 CoN細胞的增殖促進作用
在12株雙歧桿菌中,只有菌株H3-R2的發酵上清液(圖1)和細胞破碎物(圖2)均對CCD841 CoN細胞有增殖促進作用且顯著高于陽性對照組(P<0.05),具有潛在研究價值。
菌株H3-R2的發酵上清中存在著蛋白質[26]、多糖[27]等物質,破碎物中存在著蛋白質[28]、多糖[29]、脂磷壁酸[10]等物質。由于這些物質的存在,可能介導了CCD841 CoN細胞的增殖。已有研究發現鼠李糖乳桿菌GG蛋白產物能夠促進新生小鼠腸道的功能性成熟,包括腸道上皮細胞增殖、分化及緊密連接蛋白形成等[30]。為了了解雙歧桿菌H3-R2的表面蛋白和分泌蛋白是否對腸上皮細胞增殖有促進作用,本文提取了這兩種蛋白進行后續研究。
2.2 雙歧桿菌H3-R2表面蛋白和分泌蛋白的鑒定
表面蛋白經SDS-PAGE分析后(圖3),分子量在大約43 kDa的蛋白質得到極大的富集,是提取物中的主要蛋白質種類。已知表面蛋白分子量范圍25~71 kDa[31],并且與嗜酸乳桿菌表面蛋白相對分子質量多在41~49 kDa之間相接近[32]。說明本試驗采取的LiCl提取法可提取到雙歧桿菌H3-R2的表面蛋白。
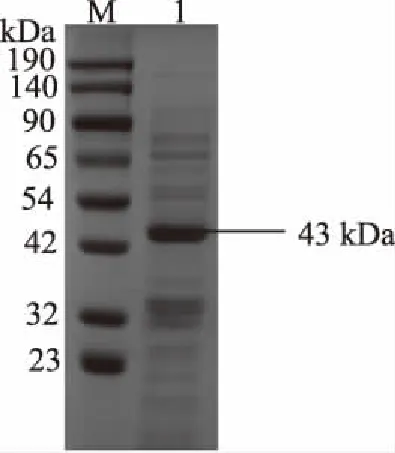
圖3 表層蛋白電泳圖
分泌蛋白經SDS-PAGE分析后(圖4),分子量在大約34 kDa和45 kDa。這與Bifidobacterium longum FNCC2705分泌蛋白相對分子質量在20~60 kDa分布均勻相接近[33]。說明本試驗采取的三氯乙酸沉淀法可提取到雙歧桿菌H3-R2的分泌蛋白。
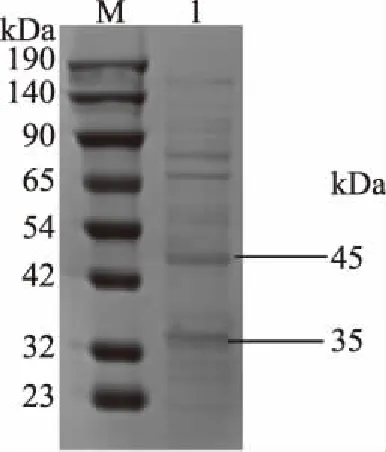
圖4 分泌蛋白電泳圖
2.3 雙歧桿菌H3-R2對CCD841 CoN細胞的增殖促進作用
2.3.1 表面蛋白的作用 菌株H3-R2表面蛋白對CCD841 CoN細胞的增殖促進作用受蛋白添加濃度和培養時間影響(圖5)。添加表面蛋白與CCD841 CoN細胞共作12 h時,與陰性對照組比細胞活性最大增加至114%;蛋白濃度為50 μg/mL組對細胞有最大增殖促進作用,且顯著高于陽性對照組(12 h)(P<0.05)。添加表面蛋白與CCD841 CoN細胞共作24 h時,與陰性對照組比細胞活性增加113%~145%;蛋白濃度為50 μg/mL組對細胞有最大增殖促進作用,且顯著高于陽性對照組(24 h)(P<0.05)。添加表面蛋白與CCD841 CoN細胞共作48 h時,與陰性對照組比細胞活性增加110%~136%;蛋白濃度為25 μg/mL組對細胞有最大增殖促進作用,且顯著高于陽性對照組(48 h)(P<0.05)。
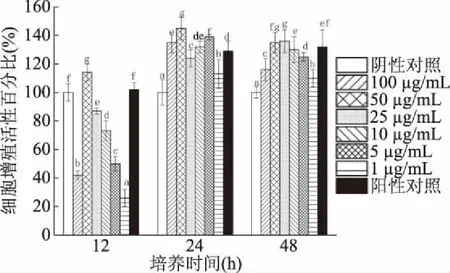
圖5 菌株H3-R2的表面蛋白對CCD841 CoN細胞的增殖促進作用
添加表面蛋白濃度50 μg/mL與細胞共作24 h的細胞增殖促進作用,高于蛋白濃度50 μg/mL與細胞共作12 h和蛋白濃度25 μg/mL與細胞共作48 h(P<0.05),是表面蛋白對細胞增殖促進作用的最佳條件。
2.3.2 分泌蛋白的作用 菌株H3-R2分泌蛋白對CCD841 CoN細胞的增殖促進作用受蛋白添加濃度和培養時間影響(圖6)。添加分泌蛋白與CCD841 CoN細胞共作12 h時,與陰性對照組比細胞活性增加102%~110%;蛋白濃度為10 μg/mL組對細胞有最大增殖促進作用,且顯著高于陽性對照組(12 h)(P<0.05)。添加分泌蛋白與CCD841 CoN細胞共作24 h時,與陰性對照組比細胞活性增加111%~128%;蛋白濃度為10 μg/mL組對細胞有最大增殖促進作用,且與陽性對照組(24 h)無顯著性差異(P<0.05)。添加表面蛋白與CCD841 CoN細胞共作48 h時,與陰性對照組比細胞活性增加104%~121%;蛋白濃度為5 μg/mL組對細胞有最大增殖促進作用,且與陽性對照組(48 h)無顯著性差異(P>0.05)。
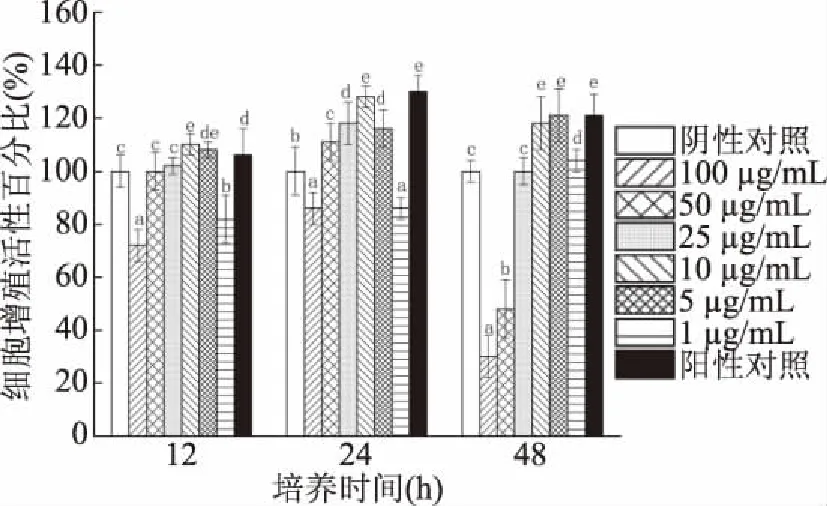
圖6 菌株H3-R2的分泌蛋白對CCD841 CoN細胞的增殖促進作用
添加分泌蛋白濃度10 μg/mL與細胞共作24 h的細胞增殖促進作用,高于蛋白濃度10 μg/mL與細胞共作12 h和蛋白濃度5 μg/mL與細胞共作48 h,是分泌蛋白對細胞增殖促進作用的最佳條件。
綜上所述,選取添加菌株H3-R2表面蛋白50 μg/mL為表面蛋白組,分泌蛋白濃度10 μg/mL為分泌蛋白組,與CCD841 CoN細胞共作24 h后,進行細胞周期檢測以及增殖相關基因表達檢測。
2.4 雙歧桿菌H3-R2表面蛋白和分泌蛋白對CCD841 CoN細胞周期的影響
細胞的周期循環速度決定了細胞的增殖速度[34]。因此可以猜測,細胞添加表面蛋白或分泌蛋白后,細胞周期循環加速進而實現細胞增殖。為證實這一觀點,本文采用流式細胞術檢測表面蛋白和分泌蛋白對CCD841 CoN細胞周期循環的影響。
流式細胞術分析所示,第一個紅色峰為G0/G1期,第二個紅色峰代表G2/M期,兩峰之間的斜線填充部分為S期(圖7)。對照組、表面蛋白組和分泌蛋白組與CCD841 CoN細胞共作24 h后,各個細胞周期發生變化(圖8)。對照組的周期分布是G0/G1期為56.29%,S期為23.37%,G2/M期為20.34%;表面蛋白組的周期分布是G0/G1期為48.24%,S期為30.38%,G2/M期為21.38%;分泌蛋白組的周期分布是G0/G1期為53.10%,S期為25.56%,G2/M期為21.34%。與對照組相比,表面蛋白組的G0/G1期細胞比例下降了14.30%,S期細胞比例上升了30.00%;與對照組相比,分泌蛋白組的G0/G1期細胞比例下降了5.67%,S期細胞比例上升了9.37%。上述結果表明,菌株H3-R2的表面蛋白和分泌蛋白
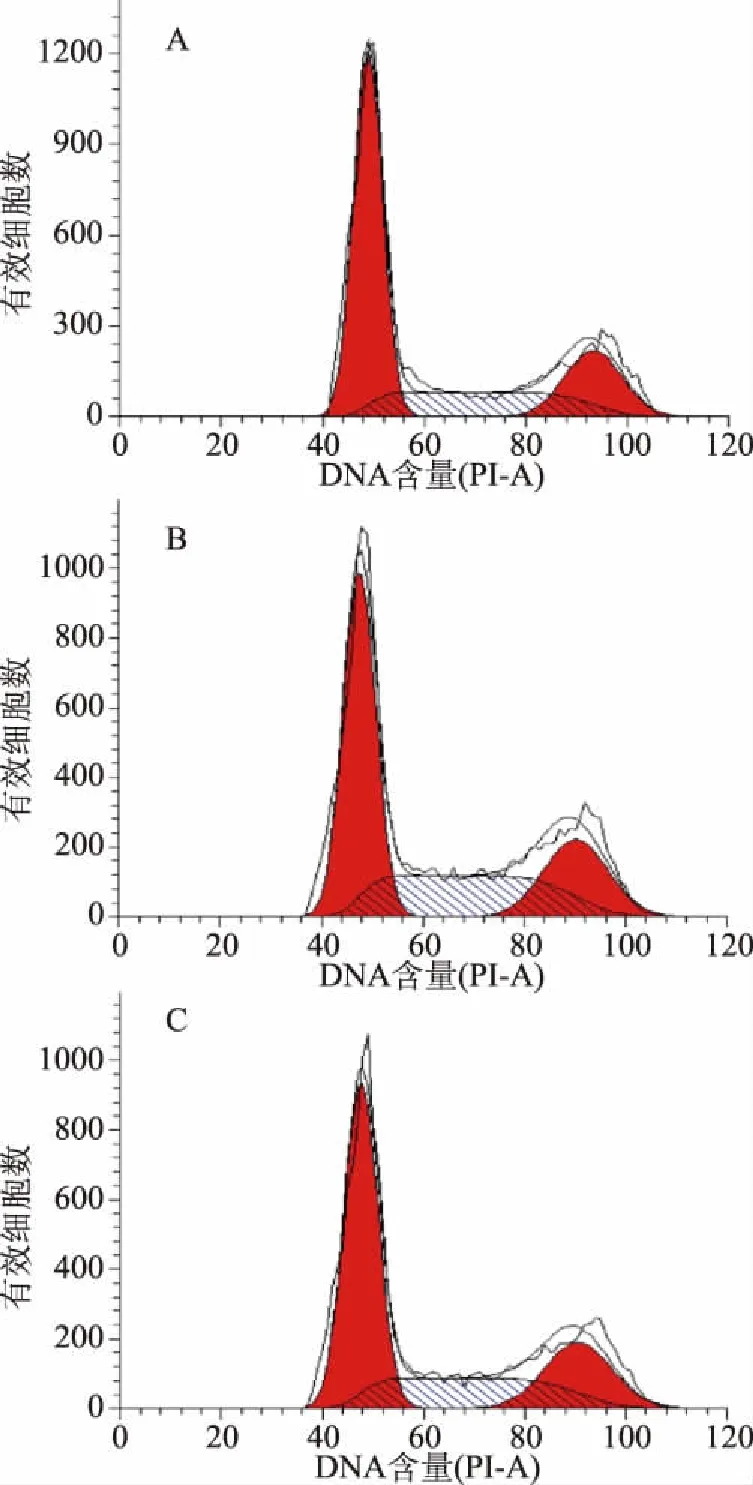
圖7 表面蛋白和分泌蛋白對CCD841 CoN細胞周期分布的影響
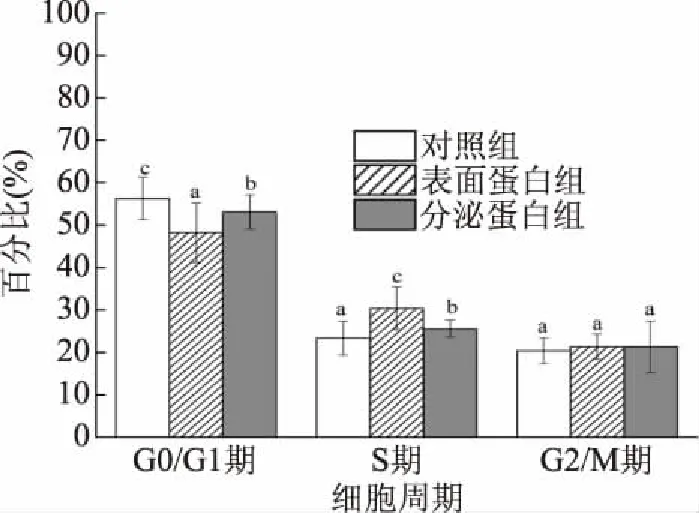
圖8 表面蛋白和分泌蛋白細胞對各個細胞周期所占的百分比的影響
2.5 雙歧桿菌H3-R2表面蛋白和分泌蛋白對增殖相關基因表達的影響
Lee等[8]的研究表明,給小鼠喂食雙歧桿菌和乳桿菌等乳酸產生菌,可顯著促進腸上皮的腸道干細胞、潘氏細胞和杯狀細胞的增殖。乳酸作用于潘氏細胞和腸基質細胞上的G蛋白偶聯受體Gpr81,影響β-catenin、c-Myc、Cyclin D1、Axin2和GSK-3β等相關基因的表達,刺激腸道細胞的增殖。
增殖相關基因進行熒光定量PCR檢測結果見表2,與對照組相比,表面蛋白組和分泌蛋白組與CCD841 CoN細胞共作用24 h后,胞內β-catenin、c-Myc和Cyclin D1基因的表達量均顯著上調(P<0.05),Axin2、GSK-3β基因的表達量顯著下調(P<0.05)。該結果表明,在表面蛋白或分泌蛋白的作用下,抑制了降解β-catenin復合物(Axin1/2、GSK-3β等蛋白形成)的激活,β-catenin 可在細胞內蓄積并向細胞核轉移,起始轉錄c-Myc、CyclinD1等靶基因,促進細胞增殖和遷移。其中CyclinD1與細胞周期直接相關[35],高表達時表明加快細胞從G0/G1期進入S期,從而促進細胞增殖,這與上述細胞周期檢測結果相一致。因此可以從轉錄組水平驗證了細胞增殖結果。

表2 表面蛋白和分泌蛋白的添加對相關基因表達的影響
3 結論
本試驗從12株嬰兒源雙歧桿菌中篩選出雙歧桿菌H3-R2,其發酵上清液和破碎物均對CCD841 CoN細胞有較好的增殖促進作用,表明該菌株具有促進新生兒腸道發育功能的潛在研究價值。菌株H3-R2的表面蛋白和分泌蛋白成分對細胞增殖有一定的積極作用,可加速細胞周期循環,并在轉錄組水平上調節增殖相關基因變化。本文初步探討了雙歧桿菌H3-R2對人胎結腸上皮細胞增殖促進作用及機制,但對蛋白成分的作用程度和具體的蛋白組分仍不清楚,有待進一步研究。
