靈武長棗不同冠層光質組成對光合特性及果實品質的影響
2020-10-21 01:40:24陳麗華宋麗華姜文倩
新疆農業科學 2020年10期
陳麗華,宋麗華,高 露,謝 云,姜文倩
(寧夏大學農學院,銀川 750021)
0 引 言
【研究意義】光是影響植物生長發育的基本環境因子之一,是植物進行光合作用的能量來源[1]。光包括光質、光照強度和光周期。光質是由不同波長的太陽光譜組成,其中可見光對植物的光合特性及果實品質的形成尤為重要,不同波長的可見光對植物形態建成、光合作用、果實品質及著色具有重要調節作用[2-3]。由于太陽光輻射到植物體過程中會受到大氣及顆粒物反射、吸收和散射,致使植物最終吸收的太陽光的能量和分布發生改變[4]。樹冠內光照分布及光合作用強弱是果實品質形成的直接因素[5],而光照分布中光照強度外,光質組成研究報道較少。靈武長棗(ZizyphusjujubaMill.cv.Lingwuchangzao)是寧夏地方特色經濟林樹種,近年來靈武長棗果實品質下降,研究靈武長棗不同冠層光質組成對光合特性及果實品質的影響,對寧夏靈武長棗適宜的光環境選擇和從樹冠結構與光質調控方面提高果實品質提供參考。【前人研究進展】目前,利用人為遮光過濾的方法調控光質對植物生長發育、光合特性、果實品質形成過程的影響及調控機制研究較多,而對于自然光照下研究不同冠層光質組成對光合能力及果實品質的相關研究較少,且前人的研究中,把光質、光合特性與果實品質及著色聯系起來做較全面研究的報道很少。相關研究在蘋果[5]、梨[6]、葡萄[7]、臍橙[8]等果樹已有研究,但未見對棗的研究相關報道。研究寧夏靈武長棗不同冠層光質組成對光合特性及果實品質的影響。【本研究切入點】以田間靈武長棗為試驗對象,在自然光照條件下,研究光質、光合特性與果實品質及著色關系及樹冠不同冠層部位光質組成對樹體光合特性及果實品質的影響規律。【擬解決的關鍵問題】運用多元統計分析方法,建立不同波長可見光絕對輻射能與光合特性及果實品質的多元關系,分析影響光合特性及果實品質的主要可見光波長組成,為靈武長棗高效栽培實踐光質調控方面提供理論和技術支撐。
1 材料與方法
1.1 材 料
試驗于2019年5月至10月在寧夏大學科研教學實習基地進行(N38°47′07″,E106°04′00″,海拔高度1 116.8 m),屬于半干旱氣候區,年平均日照時數3 000 h,日溫差13℃,年均≥10 ℃積溫3 300 ℃,無霜期140~160 d,年均氣溫8.5 ℃,年均降水量180~200 mm,土壤為灌淤土壤。
試驗于 2019年4月中旬至10月初進行。試驗前,在田間靈武長棗種植園采用五點取樣法隨機選取5 株生長健壯且樹勢均一的7年生靈武長棗嫁接苗,掛牌標記設定為試驗樣樹,該種植園為南北行向種植,株行距 2 m×3 m,平均樹高為2.7 m,平均冠幅(SN×EW)為2.12 m×1.89 m,樹形為改良紡錘形,常規栽培管理。利用水肥一體化裝置采用滴灌形式保證水肥供應的一致性。在樹冠半徑的剖面上,按從上到下將樹冠均勻劃分為上層、中層和下層三層,將已分出的樹冠中層從內到外均勻劃分為內部、中部和外部三個部分。
1.2 方 法
1.2.1 樹冠不同冠層光質
采用ATP2000光纖光譜儀于09:00~11:00分別在選定的5個樣株劃分的不同冠層內連續隨機選取10個代表點,測定其350~770 nm光質成分的絕對輻射能分布。于7月中旬和8月中旬各測定1次。
1.2.2 樹冠不同冠層光合特性的測定
于7和8月中旬各測定1次光合指標,選取晴朗的天氣于09:00~11:00用TARGAS-1便攜式光合測定儀測定葉片凈光合速率(Pn)、氣孔導度(Gs)、蒸騰速率(Tr)、胞間CO2濃度(Ci)、有效水分利用率(WUE),5個樣株各冠層內隨機選擇10片成熟功能葉片進行測定,每片葉的測定中儀器自動重復5組數據。
1.2.3 樹冠不同冠層葉綠素熒光特性
采用OS5P型便攜式脈沖調制葉綠素熒光儀(Opti-science,USA)在Yield和Kinetic模式下,分別對靈武長棗成熟葉片的葉綠素熒光參數進行測定。Kinetic模式下測定前,葉片暗處理15~20 min。5個樣株各冠層內隨機選擇10片成熟功能葉片測定,每片葉的測定中儀器自動重復5組數據。
1.2.4 樹冠不同冠層果實品質
于果實成熟期(9月28日)對每樣株分別從每個冠層的東、西、南、北及上、中、下、里、外9個方向采取棗吊基部第2節處同一批開花結果的無病蟲害果實(約20個)。各冠層果實混合后放入4℃冰箱保存用來測定果實品質各指標。
果肉總糖含量:參照鄒琦[9]的方法,采用蒽酮-硫酸比色法測定。
果肉可滴定酸含量:采用酸堿中和法測定。
果皮花青素含量: 采用pH值示差法測定。
果皮葉綠素、類胡蘿卜素含量: 采用分光光度法測定。
1.3 數據處理
采用Excel 2016對原始數據進行整理,采用SPSS 25.0進行方差分析與各處理間差異顯著檢驗(Duncan新復極差法),采用Origin 2018進行相關性(Pearson)及多元統計分析。
2 結果與分析
2.1 靈武長棗不同冠層的光質分布
研究表明,靈武長棗不同冠層可見光絕對輻射能呈極顯著差異(P紫光=0.000<0.01,P藍光=0.000<0.01,P綠光=0.000<0.01,P黃光=0.000<0.01,P橙光=0.000<0.01,P紅光=0.000<0.01),可見光絕對輻射能均在樹冠上層表現最大,靈武長棗樹冠上層可截獲更多的可見光絕對輻射能。不同冠層間紫光、藍光和綠光的絕對輻射能均表現為:上層>中層>下層,下層比上層分別減少88.13%、85.78%和27.58%,波長較短的可見光絕對輻射能在不同冠層間由上到下逐漸減小;黃光、橙光和紅光的絕對輻射能均表現為上層>下層>中層,中層比上層分別減少73.06%、75.44%和70.15,表明波長較長的可見光絕對輻射能在樹冠下層比中層截獲更多。不同部位紫光、藍光和紅光的絕對輻射能均在外部最大,且藍光和紅光的絕對輻射能均表現為外部>中部>內部,內部比外部分別減少79.22%和79.15%;黃光和橙光的絕對輻射能在中部最大,綠光的絕對輻射能在內部最大,表明波長較短的藍光和波長較長的紅光的絕對輻射能在不同部位由外到內逐漸減小,而綠光的絕對輻射能恰恰相反。表1
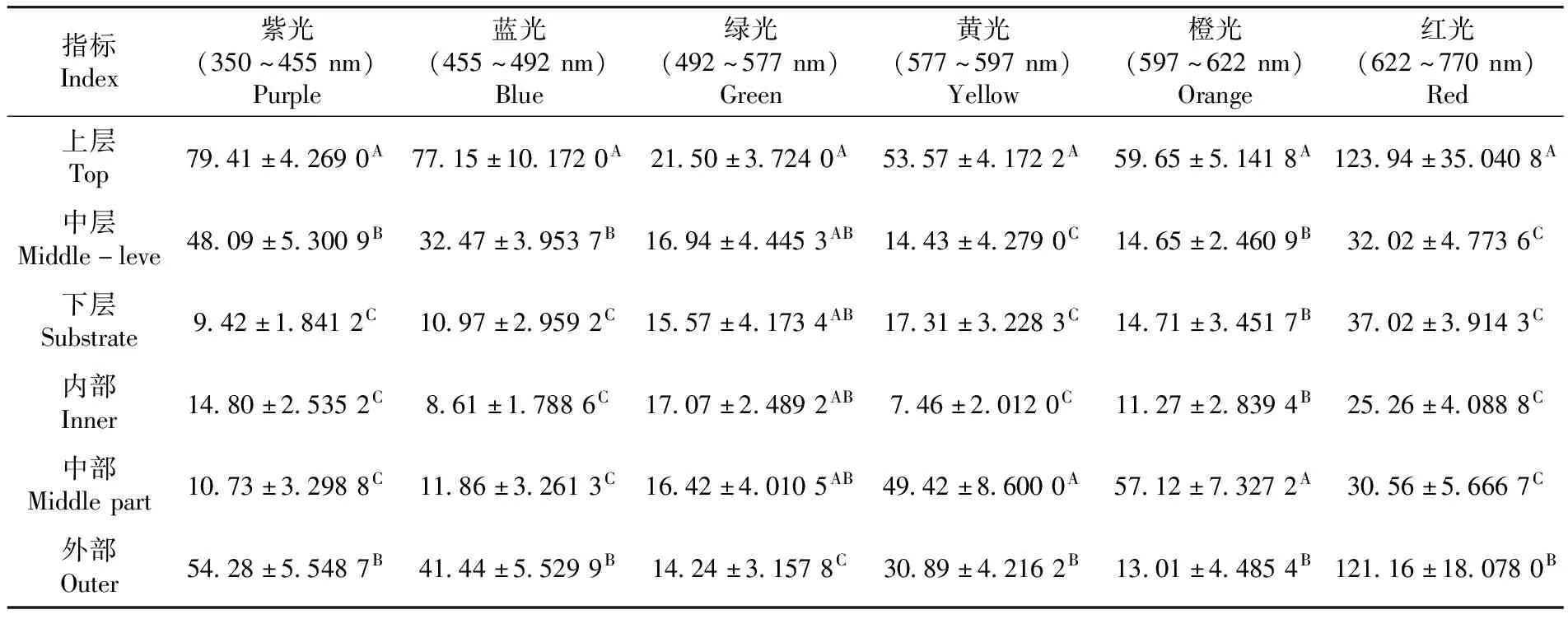
表1 靈武長棗不同冠層可見光(350~770 nm)絕對輻射能比較
2.2 靈武長棗不同冠層葉片光合參數差異性比較
研究表明,靈武長棗不同冠層葉片光合參數均呈顯著差異(PTr=0.044<0.05,PGs=0.036<0.05,PPn=0.025<0.05,PCi=0.017<0.05,PWUE=0.039<0.05),其中葉片Tr、Gs和Pn均在上層表現最大,內部表現最小,上層是內部的1.9~3.3倍,而葉片Ci和WUE在外部表現最大。不同層間葉片Tr和Gs均表現為上層>中層>下層,下層比上層分別減少了30.00%和54.45%;葉片Pn和WUE均表現為上層>下層>中層,中層比上層分別減少了47.83%和42.41%,中層葉片Ci比下層減少18.32,葉片Tr和Gs在不同冠層間由上到下逐漸減小,而葉片Pn和WUE在中層最小。不同部位葉片Tr、Gs、Pn、Ci和WUE均表現為外部>中部>內部,內部比外部分別減少了33.32%、36.87%、62.55%、36.21%、44.12%,葉片Tr、Gs、Pn、Ci和WUE在不同部位由外到內逐漸減小。表2
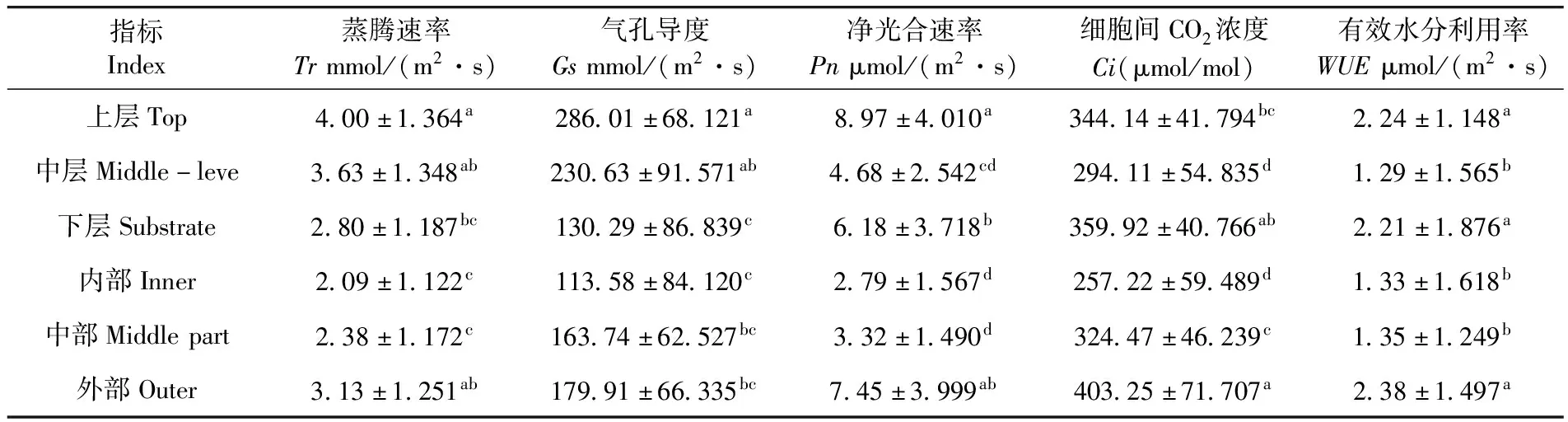
表2 靈武長棗不同冠層葉片光合參數比較
2.3 靈武長棗不同冠層葉片葉綠素熒光參數差異性比較
研究表明,靈武長棗不同冠層葉片葉綠素熒光參數Fo、Fm、Fv/Fm、qP均無差異。不同層間葉片Fm、Fv/Fm、ETR和Yield均表現為上層>中層>下層,下層比上層分別降低了14.42%、11.36%、55.50%和62.50%;葉片qP和NPQ表現為下層>上層>中層,中層比下層降低了6.25%和21.33%,葉片Fm、Fv/Fm、ETR和Yield在不同冠層間由上到下逐漸減小,而葉片qP和NPQ均在中層最小。不同部位葉片Fv/Fm、qP和ETR均表現為內部>中部>外部,外部比中部分別降低了8.98%、14.12%、40.54%;而葉片Fo和Yield表現為中部>外部>內部>,中部比內部分別降低了11.92%、40.91%,葉片Fv/Fm、qP和ETR在不同部位由內到外逐漸減小,而Fo和Yield均在外部最小。表3
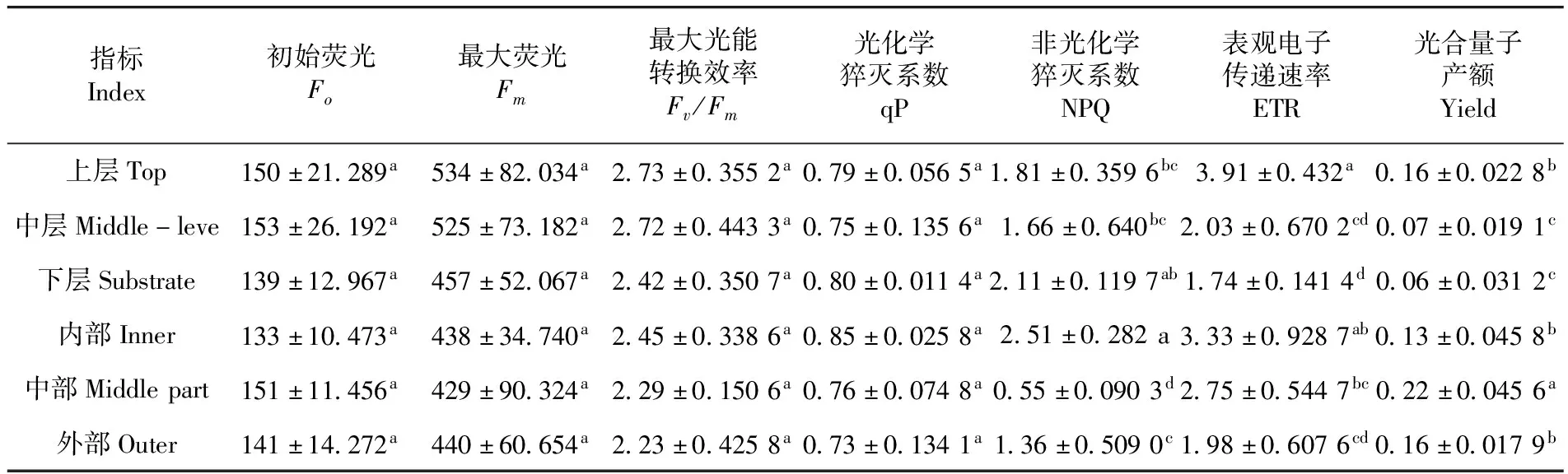
表3 靈武長棗不同冠層葉片的葉綠素熒光參數比較
2.4 靈武長棗不同冠層果實品質差異性比較
研究表明,靈武長棗不同冠層果實品質均存在顯著差異(P可滴定酸=0.048<0.05,P可溶性糖=0.031<0.05,P葉綠素=0.000<0.01,P類胡蘿卜素=0.000<0.01,P花色素苷=0.000<0.01,P類黃酮=0.000<0.01,),其中可滴定酸、可溶性糖、類胡蘿卜素均在外部表現最大,而花色素苷和類黃酮均在中部表現最大,葉綠素在內部表現最大。不同層間葉綠素和類黃酮均表現為下層>中層>上層,上層比下層分別減少了11.04%和11.90%;類胡蘿卜素和花色素苷均表現為中層>下層>上層,上層比中層分別減少了2.25%和10.71%;而可滴定酸和可溶性糖均在上層達到最大;葉綠素、類黃酮、類胡蘿卜素和花色素苷均在上層最小。不同部位可滴定酸和可溶性糖均表現為外部>中部>內部,內部比外部分別減少了20.34%和25.73%;而葉綠素和類胡蘿卜素均表現為內部>外部>中部,中部比內部分別減少了39.04%和4.14%;花色素苷和類黃酮均在中部表現最大。表4
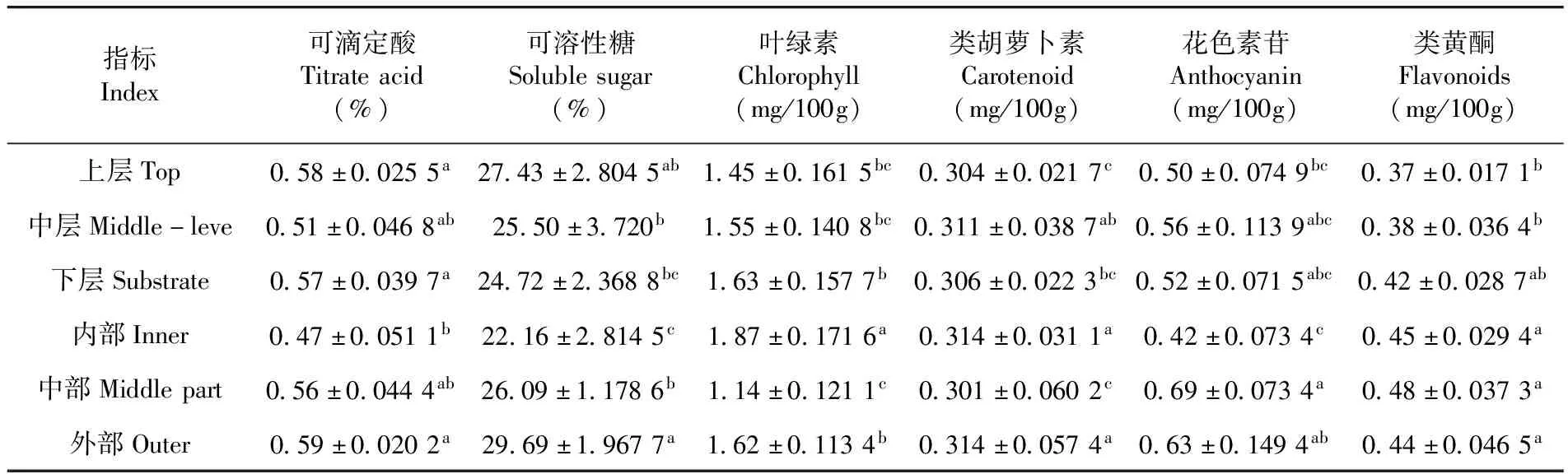
表4 靈武長棗樹不同冠層果實品質比較
2.5 靈武長棗不同冠層光質與光合特性及果實品質的相關性
研究表明,除綠光外,紅光與其他可見光均呈顯著或極顯著正相關,其中相關系數較高的有紫光(相關系數為0.852)、藍光(相關系數為0.936)和黃光(相關系數為0.624);紫光與藍光呈極顯著正相關(相關系數為0.952);藍光和橙光呈顯著正相關(相關系數為0.562);黃光和橙光呈極顯著正相關(相關系數為0.915)。Tr與藍光和紫光均呈極顯著正相關;Gs與紫光、藍光、橙光和紅光呈顯著或極顯著正相關;Pn與紫光、藍光、黃光、橙光和紅光呈顯著或極顯著正相關;Ci與藍光、橙光和紅光呈顯著或極顯著正相關;WUE與紫光、藍光、橙光和紅光呈顯著或極顯著正相關。Fo與藍光和紅光均呈極顯著正相關;Fm與紫光、藍光、橙光和紅光呈顯著或極顯著正相關;Fv/Fm紫光、藍光和紅光呈顯著或極顯著正相關;qP與藍光呈顯著正相關,與紅光呈極顯著正相關;NPQ與紫光、藍光紅光呈顯著或極顯著正相關;ETR與紫光、藍光、黃光、橙光和紅光呈顯著或極顯著正相關;Yield與紫光和紅光均呈顯著正相關。可滴定酸與紫光、藍光、綠光和紅光均呈顯著正相關;可溶性糖與紫光、藍光、橙光和紅光呈顯著或極顯著正相關,與綠光呈極顯著負相關;類胡蘿卜素與紫光、藍光、黃光和紅光呈顯著或極顯著正相關,與綠光呈顯著負相關;葉綠素與藍光、綠光、黃光和紅光呈顯著或極顯著正相關,與橙光呈顯著負相關;花色素苷與藍光呈極顯著正相關,與綠光呈極顯著負相關;類黃酮與紫光、藍光、黃光和橙光呈顯著或極顯著正相關,與綠光呈顯著負相關。表5
2.6 影響靈武長棗光合特性及果實品質的不同波長可見光篩選
研究表明,不同波長可見光絕對輻射能和葉片光合特性及果實品質分別為不同的正態總體,靈武長棗樹冠可見光絕對輻射能間的相關系數大于 0.6 的有 5項,不同波長可見光絕對輻射能間存在多重共線性,試驗應用多元相關分析,以紫光(X1)、藍光(X2)、綠光(X3)、橙光(X4)和紅光(X5)為一個總體,蒸騰速率(Y1)、氣孔導度(Y2)、凈光合速率(Y3)、細胞間CO2濃度(Y4)、水分利用效率(Y5)、初始熒光(Y6)、最大熒光(Y7)、最大光能轉換效率(Y8)、光化學猝滅系數(Y9)、非光化學猝滅系數(Y10)、表觀電子傳遞速率(Y11)、光合量子產額(Y12)、可滴定酸(Y13)、可溶性糖(Y14)、葉綠素(Y15)、類胡蘿卜素(Y16)、花色素苷(Y17)和類黃酮(Y18)為一個總體。根據多元相關系數的大小,篩選出有利于靈武長棗葉片光合特性及果實品質的不同波長可見光絕對輻射能,并對篩選出的不同波長可見光絕對輻射能與葉片光合特性及果實品質的指標建立多元線性方程。表5
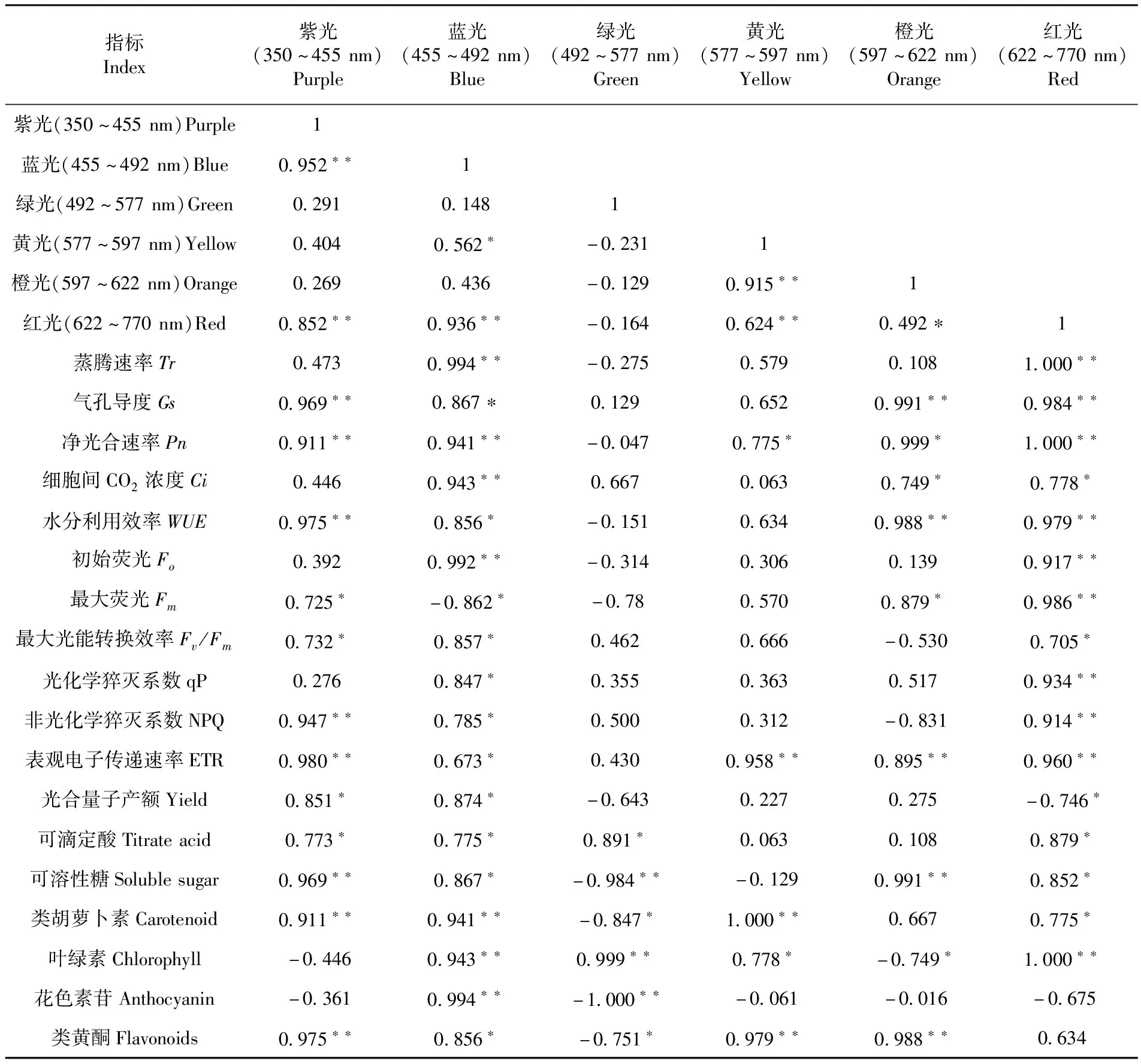
表5 靈武長棗不同波長可見光分布與光合特性及果實品質的相關性
靈武長棗葉片光合特性及果實品質主要受紫光、藍光、綠光、橙光和紅光的影響,葉片光合參數均與藍光呈正相關,與綠光呈負相關;Gs與紫光和橙光均呈正相關,其他葉片光合參數均與紫光和橙光呈負相關;WUE與紅光呈正相關,其他葉片光合參數均與紅光呈負相關。葉綠素熒光參數中Fo、ETR和Yield與紫光呈正相關,與藍光呈負相關,其他葉片葉綠素參數均與紫光呈負相關,與藍光呈正相關;Fo和Fm與綠光呈負相關,其他葉片葉綠素熒光均與綠光呈正相關;Fo、ETR和Yield與橙光呈正相關,其他葉片葉綠素熒光均與橙光呈正相關;Fo、Fm和Fv/Fm與紅光呈正相關,而其他葉片葉綠素熒光均與紅光呈正相關。果實品質中可滴定酸和可溶性糖與紫光呈負相關,其他果實品質均與紫光呈正相關,葉綠素、類胡蘿卜素和類黃酮與藍光呈負相關;其他果實品質均與藍光呈正相關;葉綠素與綠光呈正相關,其他果實品質均與綠光呈負相關;可滴定酸和葉綠素與橙光呈負相關,其他果實品質均與橙光呈正相關;葉綠素、類胡蘿卜素和類黃酮與紅光呈正相關,其他果實品質均與紅光呈負相關。應用典型相關分析篩選出的影響葉片光合特性及果實品質的不同波長可見光輻射能與通過簡單相關性選擇的不同波長可見光輻射能存在較大差異。表6
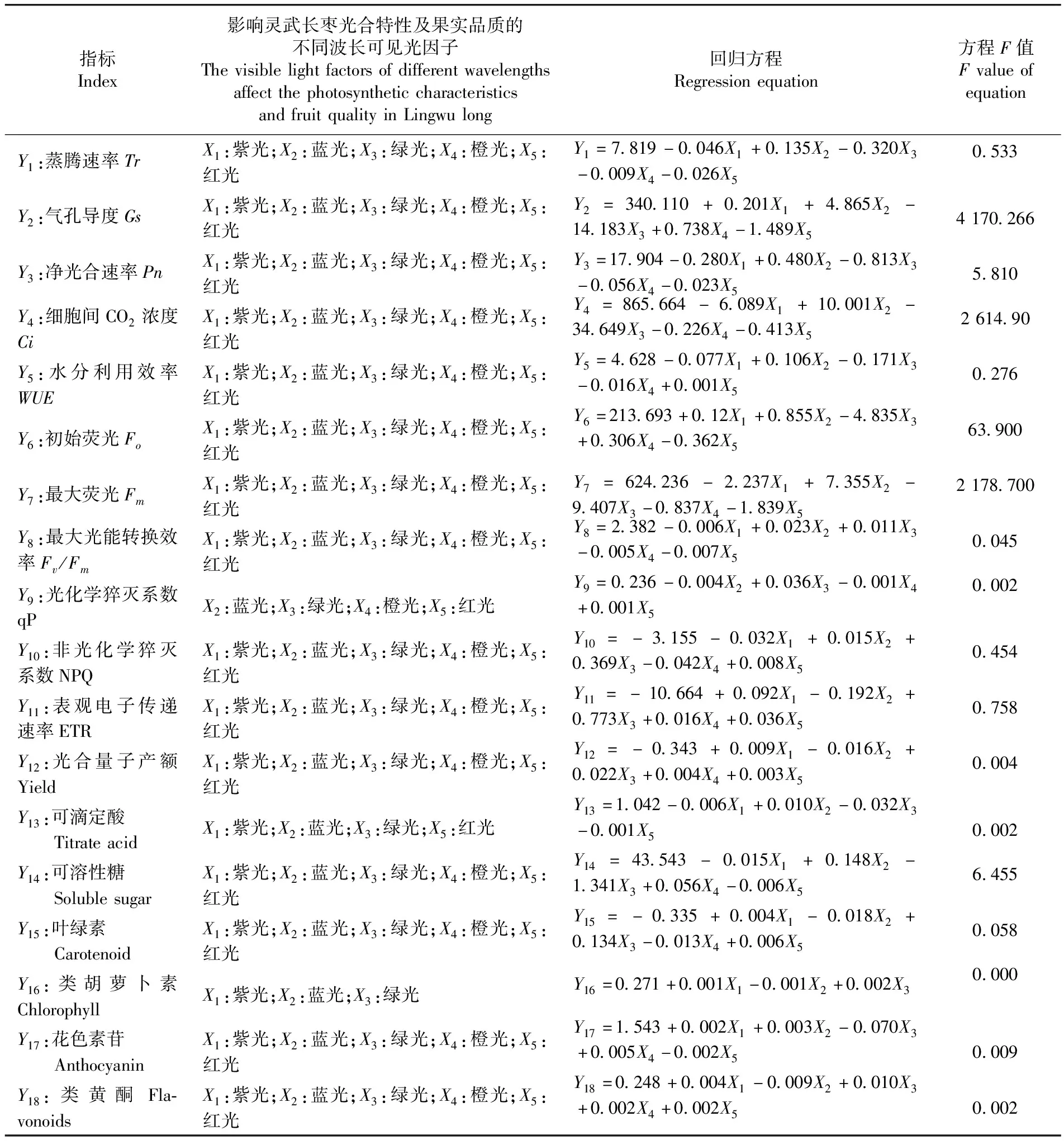
表6 影響靈武長棗光合特性及果實品質的不同波長可見光的篩選及回歸方程建立
3 討 論
太陽輻射透過樹冠過程中,不同波長可見光輻射能被吸收、反射和散射的程度不同,導致不同冠層葉片對可見光的吸收具有差異性。由于冠層的上層和外部受到太陽光的直射,而內部和下層主要受到太陽光的散射,太陽直射光中紫光較多,而太陽散射光中黃光較多[10]。研究表明,靈武長棗樹冠上層受到可見光(350~770 nm)絕對輻射能均比其他冠層多,上層、下層、內部和外部主要受到波長較長的紅光絕對輻射能,中層主要受到波長較短的紫光絕對輻射能,中部主要受到橙光和黃光的絕對輻射能,不同冠層間紫光和藍光的絕對輻射能均由上層到下層依次減小,不同部位藍光和紅光的絕對輻射能均由外部到內部依次減小。
靈武長棗不同波長可見絕對光輻射能與光合特性及果實品質間的關系非常復雜,不同波長可見光之間呈顯著或極顯著正相關關系,表明不同波長可見絕對光輻射能之間存在不同程度的協同作用,這可為靈武長棗補光提供一定的理論依據。本研究中除綠光外,紅光與其他可見光絕對輻射能均呈顯著或極顯著正相關,增加波長較長的紅光可以提高光質質量。
光合作用是植物體內重要的生理過程,而光是光合作用的基礎。植物對可見光的吸收中,紅、橙光主要是被葉綠素吸收,其次藍、紫光也能被葉綠素、胡蘿卜素吸收[11]。光質對葉片氣孔的結構和特性有影響[12]。藍光對植物葉片的氣孔導度具有重要誘導作用[13]。氣孔作為植物光合作用過程中 CO2的輸入和水蒸氣的輸出通道,因此氣孔導度對植物的凈光合速率和蒸騰速率具有重大意義,試驗發現,靈武長棗葉片的蒸騰速率和氣孔導度從樹冠上層到下層及外部到內部依次減小,葉片光合參數均與藍光絕對輻射能呈正相關,與綠光絕對輻射能呈負相關,藍光很大程度上可以改變靈武長棗光合作用,而綠光可以抑制靈武長棗光合作用。
葉綠素熒光動力學技術在測定葉片光合作用過程中光系統對光能的吸收、傳遞、耗散、分配等方面具有獨特的作用,與光合的氣體交換指標相比,葉綠素熒光參數更具有反映植物光合的內在特征指標,是分析植物光合生理的重要技術手段[14]。有研究報道,紅光照射下Fo、Fm均顯著高于其他光質,表明紅光有利于 PSⅡ電子傳遞,增加草莓的熒光產量[15]。Fv/Fm反應 PSⅡ反應中心的最大光能轉化效率,可作為光抑制的指標。紅藍混合光處理能顯著提高葉用萵苣[16]和葡萄試管苗[17]的Fv/Fm值。試驗發現,靈武長棗葉片熒光參數Fm、Fv/Fm、ETR和Yield從樹冠上層到下層依次減小,這與紅光和和藍光的絕對輻射能變化一致。
不同冠層光質組成與果實品質及著色有著密切的關系,不同光質組成可以調節糖和酸的代謝。陳田甜[18]研究發現,紅光能提高番茄的糖酸含量。宋哲等[19]研究表明,藍紫光對果實著色最為有效,本研究中靈武長棗果實可滴定酸和可溶性糖從樹冠上層到下層依次減小,采用多元相關分析表明紫光絕對輻射能與類胡蘿卜素、葉綠素、花色素苷及類黃酮均呈正相關,而可滴定酸、可溶性糖和花色素苷與藍光絕對輻射能呈正相關,葉綠素與綠光絕對輻射能呈正相關,葉綠素、類胡蘿卜素和類黃酮與紅光絕對輻射能呈正相關。進一步分析表明增加紫光和紅有利于果實著色,減少綠光可減少葉綠素。靈武長棗光合特性及果實品質與不同光質組成之間的關系復雜,試驗進一步表明應用簡單的相關性分析無法解釋靈武長棗光合特性及果實品質與不同光質組成之間的關系,不同光合特性指標和果實品質受土不同光質組成的影響各異,不同光合特性指標和果實品質受不同波長可見光的共同作用。
4 結 論
靈武長棗不同冠層光質分布存在極顯著差異。波長較短的可見光絕對輻射能由上到下逐漸減小,藍光和紅光的絕對輻射能由外到內逐漸減小。葉片光合參數由外到內逐漸減小。葉綠素熒光參數Fm、Fv/Fm、ETR和Yield由上到下逐漸減小,Fv/Fm、qP和ETR由內到外逐漸減小,滴定酸、可溶性糖、類胡蘿卜素均在樹冠外部表現最大,而花色素苷和類黃酮在樹冠中部表現最大,葉綠素在樹冠內部表現最大。不同波長可見光絕對輻射能對靈武長棗葉片光合參數、葉綠素熒光參數、果實品質等方面都有一定的影響,增加波長較長的紅光可以提高光質質量,紅光是靈武長棗光合特性及果實品質的主要影響因子,在生產中補充370~470 nm和600~770 nm波長的光質,以提高葉片光合作用及果實品質。
