生鮮畜禽肉中沙門氏菌全基因組分析與分子溯源
2020-10-19 00:32:30郭焜鵬張海韻王芳斌
食品與機械 2020年9期
宋 晟 高 晗 嚴 禮 郭焜鵬 張海韻 王芳斌
(1. 食品安全監測與預警湖南省重點實驗室,湖南 長沙 410117;2. 湖南省食品質量監督檢驗研究院,湖南 長沙 410117)
沙門氏菌廣泛分布于自然界,是一種屬于腸桿菌科的革蘭氏陰性桿菌,就邦戈爾沙門氏菌和腸道沙門氏菌這兩個物種而言,迄今就已發現2 500余種血清型。作為一種人畜共患的主要病原菌,沙門氏菌通常會引起發熱、嘔吐等不良反應,是腹瀉病全球四大病因之一[1-2]。某種情況下,沙門氏菌甚至可以進入血液,引發腦膜炎。為治療沙門氏菌病,通常采用抗微生物藥物,但大量抗生素的濫用,導致部分菌株產生耐藥性,使藥物失效[3-4]。人類患沙門氏菌病有可能是食用了被沙門氏菌污染的動物源性食品,如蛋、肉、禽和奶等。對于沙門氏菌以及其他食源性致病菌的溯源研究,主要有傳統的沙門氏菌分離鑒定技術、多位點序列分型(Multi locus sequence typing,MLST)技術以及脈沖場凝膠電泳(Pulsed field gel electrophoresis,PFGE),近年來,隨著新一代基因組測序技術的發展,基于全基因組測序分子分型技術廣泛應用于食源性致病菌溯源中,具有快速精準等技術優勢[5-7]。
實驗室對長沙市銷售的生鮮雞肉、生鮮豬肉進行了隨機取樣調查,依據GB 4789.4—2016分離、鑒定樣品中的沙門氏菌,以充分了解長沙市銷售的生鮮雞肉、生鮮豬肉的污染狀況,為防治沙門氏菌所引起的食源性疾病提供參考依據,并對分離鑒定出的40株沙門氏菌,進行全基因測序與分子溯源分析,研究其溯源關系、致病機理、耐藥機理,為沙門氏菌基因層面的深入研究提供一定的試驗依據。
1 材料與方法
1.1 材料
鼠傷寒沙門氏菌標準菌株ATCC14028:實驗室保存;
生鮮豬肉(24份,每份2 kg)、生鮮雞肉(27份,每份2 kg):湖南省長沙市各大農貿市場;
緩沖蛋白胨水(BPW)、四硫酸鈉煌綠增菌液(TTB)、亞硒酸鹽胱氨酸增菌液(SC)、HE瓊脂:青島海博生物技術有限公司;
亞硫酸鉍瓊脂(BS)、木糖賴氨酸脫氧膽鹽瓊脂(XLD)、革蘭氏染色液試劑盒(Gram Stain)、三糖鐵瓊脂(TSI)、營養瓊脂(NA):北京陸橋技術股份有限公司;
革蘭氏陰性菌鑒定試劑卡(GN):法國Chromagar公司。
1.2 儀器
拍擊式均質器:Masticator Silver型,西班牙IUL公司;
低溫培養箱:MIR-254L-PC型,日本松下電器產業株式會社;
全自動微生物鑒定系統:VITEK 2 compact30型,法國生物梅里埃股份有限公司。
1.3 樣品的采集與處理
將采集的新鮮豬肉24份與整雞27份放入無菌均質袋中,用BPW沖淋后,拍擊式均質器拍擊豬肉2 min,用手搓揉整雞2 min,BPW淋洗液培養18 h。
1.4 增菌與分離培養
吸取1 mL培養18 h的BPW淋洗液,轉種至10 mL TTB中,(42±1) ℃培養24 h,同時吸取1 mL培養18 h的BPW淋洗液,轉種至10 mL SC中,(36±1) ℃培養24 h。從TTB與SC中各挑1環劃線至XLD、BS、HE平板。BS平板與XLD平板(36±1) ℃培養48 h,HE平板(36±1) ℃培養24 h。
1.5 生化試驗
自選擇性瓊脂平板上挑取沙門氏菌典型菌落,接種三糖鐵,(36±1) ℃培養24 h。
1.6 革蘭氏染色
自選擇性瓊脂平板上挑取沙門氏菌典型菌落,經生理鹽水稀釋,涂片固定后,革蘭氏染色,顯微鏡觀察細菌形態。
1.7 鑒定與菌株保存
自選擇性瓊脂平板上挑取典型菌落,接種營養瓊脂(36±1) ℃培養24 h,得到純化單菌落,采用全自動微生物生化鑒定系統進行菌種鑒定。單菌落經營養瓊脂純化后用菌種保存管于-70 ℃保存。
1.8 全基因組測序
對40株沙門氏菌進行基因組DNA的提取、測序、功能基因預測注釋等工作由廣州研科生物科技有限公司完成。
2 結果與分析
2.1 菌株形態、生化鑒定結果
分離獲得的40株典型菌株,在BS瓊脂培養基上,形態為棕褐色或黑色且有金屬光澤,菌落周圍呈黑色;在HE瓊脂培養基上,形態為藍綠色,帶黑色中心;在XLD瓊脂培養基上,形態為粉紅色,帶黑色中心。接種三糖鐵瓊脂,斜面紅色產堿,底層黃色產酸,產H2S瓊脂呈黑色,產氣,瓊脂中有氣泡;經革蘭氏染色,油鏡可觀察,兩端鈍圓的紅色桿菌,革蘭氏陰性,經全自動微生物生化鑒定系統鑒定為沙門菌屬。
24份生鮮豬肉中有20份樣品檢出沙門氏菌,檢出率為83.3%;27份生鮮整雞中有20份樣品檢出沙門氏菌,檢出率為74.1%,說明零售生鮮豬肉與生鮮整雞沙門氏菌的污染率非常高。
2.2 40株沙門氏菌基因組基本信息
采用二代測序技術對40株沙門氏菌進行全基因組測序,并進行序列組裝。由表1可知,豬源性沙門氏菌的scaffold數目在24~71,基因組總長度分布為3 842 688~5 019 323 bp,GC含量平均為54.08%。由表2可知,基因數在4 012~4 382,基因平均長度在932~958 bp。雞源性沙門氏菌的scaffold數目在23~73,基因組總長度分布為4 638 420~5 201 462 bp,GC含量平均為51.93%。基因數在4 016~4 476,基因平均長度在930~959 bp。20株豬源性沙門氏菌基因組總長度的平均值比20株雞源性沙門氏菌基因組總長度的平均值小425 799 bp,而GC平均含量卻比20株雞源性沙門氏菌多2.15%,說明豬源性沙門氏菌與雞源性沙門氏菌在基因組層面存在差異,也暗示兩者來源不同,在運輸與銷售環節的交叉污染可能性小,而飼養、屠宰運輸環節豬與豬之間、雞與雞之間污染可能性大。
2.3 豬源性、雞源性沙門氏菌單拷貝直系同源基因分析
直系同源是指不同物種之間的某一部分序列具有同源性,例如蛋白質的同源性,DNA序列的同源性,而單拷貝直系同源基因是指在不同物種中具有相同序列,相同功能,但基因組中拷貝數為1的基因。單拷貝直系同源基因,大部分為管家基因,在分子系統學中起重要的分子標記作用,通常用于構建生命樹的主干及主干和末梢之間的分枝[8]。分別對20株雞源性沙門氏菌與20株豬源性沙門氏菌的基因組信息進行分析,統計基因家族、單拷貝直系同源基因、多拷貝直系同源基因、獨立旁系同源基因、其他直系同源基因。由表3、4可知,20株豬源性沙門氏菌共有3 988個基因家族,單拷貝直系同源基因數在2 170~2 191,雞源性沙門氏菌共有3 919個基因家族,單拷貝直系同源基因數在2 159~2 183。單拷貝直系同源基因主要包括內肽酶活性基因、水解酶基因、蛋白質分泌調節基因、核糖體結構基因、谷氨酰-tRNA還原酶基因等。

表1 20株豬源性沙門氏菌基因組信息
2.4 基于單拷貝直系同源基因分別構建豬源性、雞源性沙門氏菌系統進化樹
基于單拷貝直系同源基因分別構建豬源性沙門氏菌系統進化樹(見圖1),結果顯示20株豬源性沙門氏菌明顯聚集為5簇,FC10728、FC11983、FC10743、FC11961、FC11980 5株聚集為1簇,FC10733、FC12045、FC10701、FC10726 4株聚集為1簇,FC10738、FC10718、FC10717 3株聚集為1簇,FC10758、FC11967 2株聚集為1簇,FC10739、FC10745 2株聚集為1簇,另有4株FC10755、FC11958、FC10730、FC10702無明顯聚集。
FC10728、FC11983、FC10743、FC11961、FC11980 5株豬源性沙門氏菌聚集為1簇,但采樣點各不相同,說明該菌種在不同銷售點均存在污染,而污染源可能是來源于同一個屠宰基地或同一個養殖生產基地。FC10733與FC10701聚集為1簇,且來源于同一銷售點,說明銷售點存在交叉污染,或來源于同一供貨渠道。另有FC10707與FC10743,FC11983與FC119582分別來源于2個銷售點,且并不聚集,說明該銷售點所銷售的豬肉存在多株豬源性沙門氏菌污染的情況,可能是不同的供貨渠道所致。從同源性的程度來看,FC10701、FC10726與FC10733同源性極高,分別來源于2個銷售點,FC10717與FC10738同源性極高,分別來源于2個銷售點,FC10728與FC10743同源性極高,分別來源于2個銷售點,FC11961、FC11980與FC11983同源性極高,分別來源于3個銷售點,說明其銷售的豬肉分別來源于同一供貨渠道。
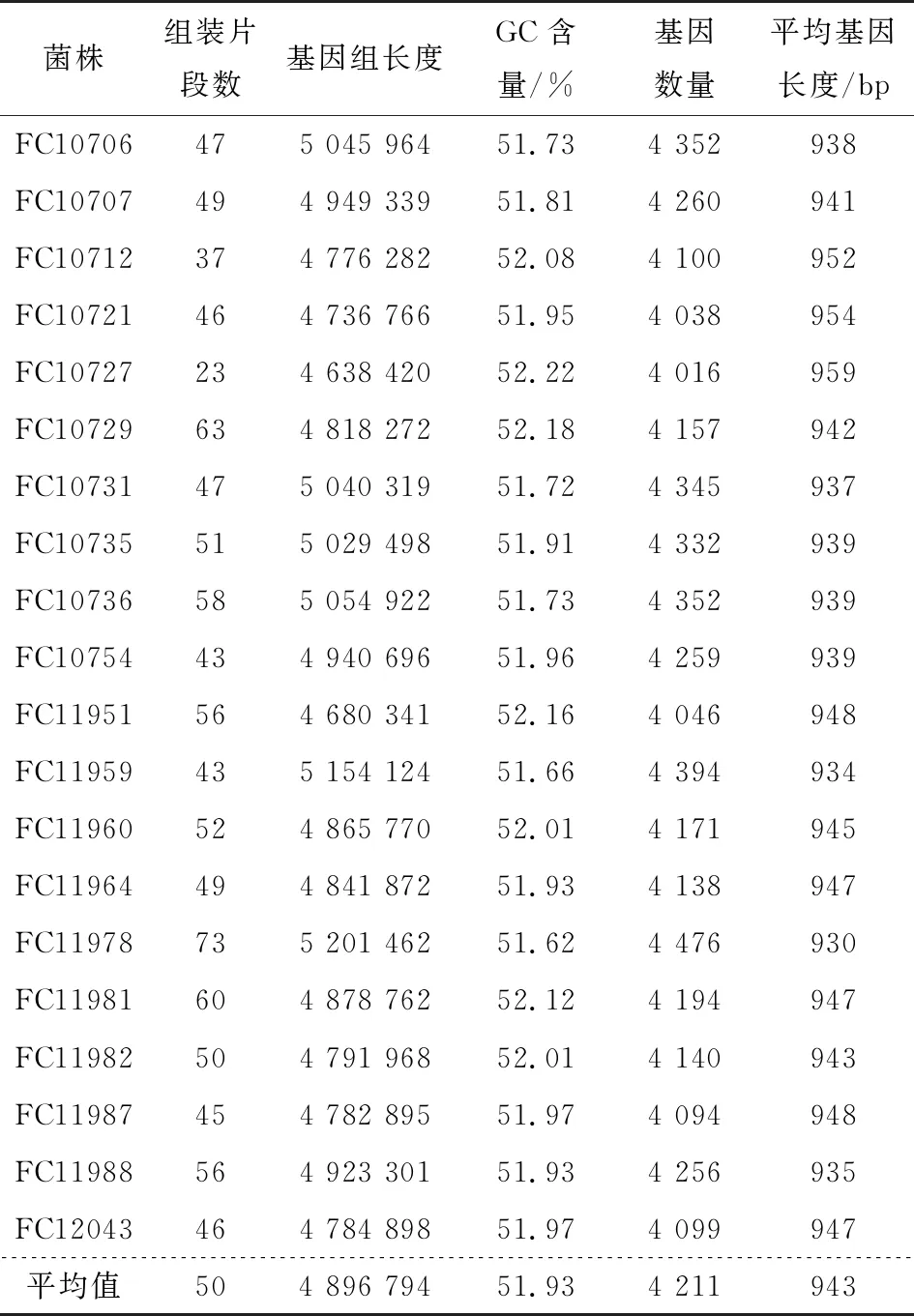
表2 20株雞源性沙門氏菌基因組信息

表3 20株豬源性沙門氏菌直系基因與旁系基因分析
基于單拷貝直系同源基因分別構建雞源性沙門氏菌系統進化樹(見圖2),結果顯示20株雞源性沙門氏菌明顯聚集為5簇,FC10706、FC10707、FC10731、FC10736、FC11978、FC11982 6株聚集為1簇,FC10721、FC11960、FC11964、FC11987、FC12043 5株聚集為1簇,FC10754、FC11959、FC11988 3株聚集為1簇,FC10729、FC11981 2株聚集為1簇,另有4株FC10712、FC10727、FC10735、FC11951無明顯聚集。
FC10706、FC10707、FC10731、FC10736、FC11978、FC11982 6株雞源性沙門氏菌聚集為1簇,其中FC10706、FC10707與FC10731來源于同一品牌超市的2個分店且同源性極高,說明同一品牌超市的供貨渠道可能存在污染,導致分店銷售的雞肉污染同一沙門氏菌。FC11982與FC11959、FC11988與FC11960分別聚集成不同簇,但來源于同一銷售點,說明該銷售點所銷售的雞肉存在多株雞源性沙門氏菌污染的情況,可能是不同的供貨渠道所致,且存在交叉污染的可能。從同源性的程度來看,FC11960、FC12043、FC11987與FC11964同源性極高,分別來源于4個銷售點,FC10754與FC11959同源性極高,分別來源于2個銷售點,FC10728與FC10743同源性極高,分別來源于2個銷售點,FC10706、FC10707、FC10731、FC10736與FC11978同源性極高,分別來源于4個銷售點,說明其銷售的雞肉分別來源于同一供貨渠道。

表4 20株雞源性沙門氏菌直系基因與旁系基因分析
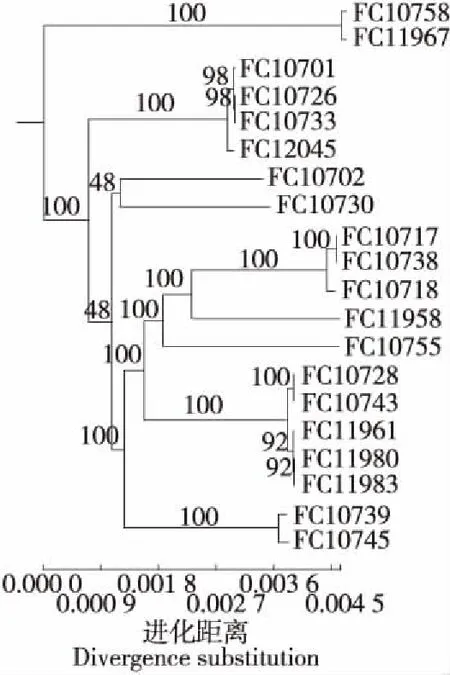
圖1 基于單拷貝直系同源基因構建20株豬源性沙門氏菌系統進化樹

圖2 基于單拷貝直系同源基因構建20株雞源性沙門氏菌系統進化樹
2.5 豬源性、雞源性沙門氏菌致病基因分析
通過數據比對與分析,發現20株豬源性沙門氏菌,20株雞源性沙門氏菌,共涉及15種致病基因,如表5和表6所示。整體比較而言,雞源性沙門氏菌致病基因較豬源性沙門氏菌多,20株雞源性沙門氏菌共有277個致病基因,而20株豬源性沙門氏菌共有271個致病基因。fimA、fimD、ipaH9.8、ipgD&sopB、K08303、ptrB、sipA&ipaA、sipB&ipaB&bipB、sipC&ipaC&bipC、sipD&ipaD&bipD、yeeJ11個基因為20株豬源性沙門氏菌所共有;fimA、fimD、ipaH9.8、ipgD&sopB、K08303、ptrB、sipA&ipaA、sipB&ipaB&bipB、sipC&ipaC&bipC、sipD&ipaD&bipD、sptP、yeeJ12個基因為20株雞源性沙門氏菌所共有。關于sopE基因,豬源性沙門氏菌中只有FC10730擁有,但雞源性沙門氏菌中有FC10706、FC10707、FC10712、FC10731、FC10736、FC11978 6株擁有該基因。
15種致病基因涉及多種致病機理。fimA,基因注釋為菌毛蛋白,特異性介導細菌對于骨髓源樹突狀細胞的粘附和侵襲作用[9]。fimD,基因注釋為外膜引入蛋白,位于菌毛頂端的蛋白,是所有血清型的共有蛋白,介導沙門氏菌直接粘附宿主細胞。yeeJ,基因注釋為黏附素,介導沙門氏菌對宿主細胞的粘附,對沙門氏菌的定植起重要作用,化學性質方面,黏附素可能為沙門氏菌表面某種特定的蛋白質結構或糖脂成分[10]。sipA&ipaA、sipB&ipaB&bipB、sipC&ipaC&bipC、sipD&ipaD&bipD,基因注釋分別為侵襲素A、侵襲素B、侵襲素C、侵襲素D,4個基因均位于致病島SPI-1,細菌進入宿主上皮細胞的關鍵因子,是沙門氏菌的關鍵毒力因子。ipaH9.8,基因注釋為侵襲質粒抗原,可抑制血小板的正常凝集,促使水腫[11-12]。sopB,基因注釋為磷脂酰肌醇-4,5-雙磷酸4-磷酸酶,是一種肌醇磷酸酶,在發病機理中起重要作用,當沙門氏菌侵入人腸道上皮細胞并分泌sopB蛋白后,將激發3-磷酸肌醇激酶依賴性的信號傳導,從而使腸上皮細胞Cl-水平提高,打亂宿主的多種信號傳導途徑。K08303,基因注釋為蛋白酶,在上皮細胞起信號傳導作用。ptrB,基因注釋為寡肽酶B[13-14]。另有4個致病基因為個別沙門氏菌所有,其中sopE,只有7株沙門氏菌具有,該基因注釋為效應蛋白,可介導肌動蛋白集合,起細菌內化作用;能刺激腸道上皮細胞導致炎癥和腹瀉,促進炎癥細胞的趨化和增殖;破壞宿主細胞的肌動蛋白,進而使得細菌侵入;引起遲發性細胞毒作用,引起黏膜炎癥反應。sopD,基因注釋為效應蛋白,在入侵真核細胞中起協同作用。sptP,基因注釋為效應蛋白,能引起至少兩種蛋白的去磷酸化,抑制肥大細胞的脫顆粒,導致抑制嗜中性粒細胞的聚集,阻止血管內容物流如感染部位,從而引起細菌的全身性感染[15]。TROVE2&SSA2,基因注釋為SS-A / Ro核糖核蛋白,是一種RNA-蛋白質復合體,功能上屬于一種紅斑狼瘡抗原,SS-A / Ro抗體可作為新生兒紅斑狼瘡(NLE)的血清學標志[16]。
2.6 豬源性、雞源性沙門氏菌耐藥基因分析
通過數據比對與分析,發現20株豬源性沙門氏菌,20雞源性沙門氏菌,共涉及15種耐藥基因,如表7和表8所示。整體比較而言,雞源性沙門氏菌與豬源性沙門氏菌的耐藥基因分布情況基本類似,20株豬源性沙門氏菌共有294個耐藥基因,20株雞源性沙門氏菌共有293個耐藥基因。acrA、ampG、ampC&penP、norR、norV、norW、nsrR、ompC、ompF、tolC、YHB1&hmp、lepA、pagP、SIG2&rpoS14個耐藥基因為20株豬源性沙門氏菌所共有;acrA、ampG、ampC&penP、norR、norV、norW、nsrR、ompC、ompF、tolC、YHB1&hmp、lepA、pagP、SIG2&rpoS14個耐藥基因為20株雞源性沙門氏菌所共有,關于vanX基因,分別有14株豬源性沙門氏菌與13株雞源性沙門氏菌有該基因。
15種耐藥基因涉及多種耐藥機制。acrA,基因注釋為膜融合蛋白;TolC,基因注釋為外膜通道蛋白,兩者相互配合,形成主動外排系統,由膜融合蛋白acrA捕獲抗菌藥物后,通過外膜通道蛋白TolC,將抗菌藥物運轉至外界。ompC、ompF,基因注釋為外膜孔蛋白,細菌接觸抗菌藥物后,發生外膜孔蛋白ompC、ompF的表達基因失活,造成孔蛋白丟失或嚴重減少,最終導致β-內酰胺類藥物進入菌體內減少,切斷β-內酰胺類藥物輸入細菌體內的途徑,以產生耐藥性。ampC&penP,基因注釋為β-內酰胺酶,可使使易感抗生素水解而滅活[17]。ampG,基因注釋為β-內酰胺酶感應信號傳感器,編碼一種依賴質子動力勢能的單要素滲透酶,起到向胞漿內傳遞誘導信號的作用。AmpG蛋白是細胞黏肽循環中的關鍵蛋白。若ampG基因缺失,細菌會完全喪失AmpCβ-內酰胺酶的誘導性或只表達低水平誘導[18]。YHB1&hmp、norR、norV、norW、nsrR,基因注釋分別為一氧化氮雙加氧酶、厭氧型一氧化氮還原酶轉錄調節因子、厭氧型一氧化氮還原酶、一氧化氮還原酶FoRd-AND(+)、Rrf2家族轉錄調節因子 & 一氧化氮敏感的轉錄阻遏物,沙門氏菌通過還原一氧化氮,實現對一氧化氮的脫毒,導致吞噬細胞無法殺死沙門氏菌,從而提高沙門氏菌對宿主的抗性[19]。PagP,基因注釋為酰基轉移酶,沙門氏菌細胞外膜的酰基轉移酶PagP,能將磷脂的C16碳脂肪酸鏈轉移到內毒素分子的脂肪酸鏈上,形成次級脂肪酸鏈,PagP產生的內毒素可以干擾TLR4的識別,使沙門氏菌對陽離子抗菌肽產生抗性[20]。SIG2 & rpoS是RNA聚合酶的主要識別因子,當沙門氏菌處于逆境條件下sigma因子啟動調控網,改變其對不同環境壓力的應答,以克服脅迫環境。lepA,基因注釋為GTP結合蛋白,能識別錯誤轉位的核糖體,使其有機會正確轉位。另有vanX為27株沙門氏菌具有,該基因注釋為D-丙氨酰-D-丙氨酸二肽酶,可將細胞胞壁中合成的D-丙氨酰-D-丙氨酸水解,導致細胞壁前質末端的二肽發生結構化變化,從而降低萬古霉素藥物分子跟細菌細胞壁前質的結合力,以獲得對萬古霉素的耐藥性[21]。

表5 20株豬源性沙門氏菌致病基因分析?
3 結論
(1) 長沙市零售的生鮮豬肉與生鮮整雞,沙門氏菌的污染率非常高,如缺乏必要的微生物學知識及防護措施知識,很容易引發交叉污染,發生沙門氏菌食物中毒事件,因此在日常的食品安全監管以及衛生督導工作中,應加強沙門氏菌來源、防護、預防治療的教育工作,消費者也應增強衛生防范意識。
(2) 通過對40株沙門氏菌進行全基因組測序,統計基因家族、單拷貝直系同源基因、多拷貝直系同源基因、獨立旁系同源基因、其他直系同源基因相關信息,并以豬源性沙門氏菌2 170~2 191個單拷貝直系同源基因,雞源性沙門氏菌2 159~2 183個單拷貝直系同源基因,構建系統進化樹,實現了豬源性、雞源性沙門氏菌的全基因溯源。
(3) 通過對豬源性、雞源性沙門氏菌致病基因的預測可知,生鮮畜禽肉中篩選出的沙門氏菌含有多種毒力因子,包括侵襲基因、分泌效應蛋白、黏附素、肌醇磷酸酶、菌毛蛋白、外膜引入蛋白、效應分子,通過協同作用,產生綜合毒力。在基因層面上闡明了沙門氏菌可能存在的致病基因與致病機制,提示了沙門氏菌潛在的致病風險,為藥物研發與疾病治療提供了分子基礎。多個耐藥基因,多種耐藥機制的發現,說明養殖環節可能存在抗生素濫用的情況,加快高耐藥性毒株的產生,因此,農業生產監管部門應加大抗生素使用危害的宣傳力度,加強對養殖戶抗生素使用的監管力度,從源頭減少抗生素的使用,降低對抗生素的依賴。
