氣相環境下丙氨酸Ca2+配合物的手性轉變機理及水分子的催化作用
2020-10-15 09:36:58徐銳英劉芳馬宏源張雪嬌潘宇楊曉翠王佐成
浙江大學學報(理學版) 2020年5期
徐銳英,劉芳,馬宏源,張雪嬌,潘宇,楊曉翠*,王佐成*
(1.白城師范學院 理論計算中心,吉林 白城137000; 2.白城師范學院 傳媒學院,吉林 白城137000;3.白城師范學院 物理學院,吉林 白城137000)
0 引 言
丙氨酸(alanine, Ala)是結構最簡單的手性氨基酸。根據旋光性可分為左-丙氨酸(L-Ala)和右-丙氨酸(D-Ala);根據構型可分為S-丙氨酸(S-Ala)和R-丙氨酸(R-Ala)。不同旋光性的Ala 作用不同,L-Ala 是蛋白質的成分之一,D-Ala 在生命體內微量存在,使氨基酸氧化酶(DAAO)在腫瘤細胞質中異位表達,阻止體內脂質氧化損傷;在細菌孢子代謝過程中起重要作用;但過量的D-Ala 會誘發某些疾病或導致衰老[1-4]。D-Ala 是生產維生素B6 的原料,可作為合成二肽甜味劑阿力甜的原料[5-6]。
研究Ala 的手性轉變具有重要意義,目前該領域已有很多研究成果。WANG 等[7]發現,光學純的Ala 分子可以消旋。文獻[8-11]采用密度泛函的B3LYP 方法研究了氣相單體Ala 的手性轉變及水分子的作用,結果表明,氣相下α-氫以氨基氮為橋遷移是其消旋的優勢通道,總包能壘約為266.1 kJ·mol-1;α-氫以羰基氧為橋遷移時,其消旋的總包能壘約為381.4 kJ·mol-1;水分子簇對其消旋反應有較好的催化作用。文獻[12]采用量子力學與分子力學組合的方法研究了SWBNNT 對Ala 手性轉變的限域影響,結果表明,SWBNNT(5,5)對Ala的消旋具有明顯的限域催化作用。文獻[13]采用色散校正泛函的?B97X-D 方法,研究了氣相下Cu2+與α-Ala 配合物的手性轉變,結果表明,Cu2+對α-Ala 的消旋反應具有催化作用。
研究氣相的金屬離子-生物分子體系,可獲得相關體系最本質的化學及物理性質等信息,同時為深入研究溶劑等復雜環境下金屬與氨基酸的作用奠定基礎。Ca2+是動植物生長過程中必需的營養元素,參與細胞內許多重要生理過程[14-15]。目前,學者們在氣相“鈣離子與氨基酸配合物的結構性質”領域做了大量工作,文獻[16-20]的研究表明,鈣離子與很多金屬離子一樣會誘導甘氨酸、絲氨酸和組氨酸等發生分子內質子遷移,且金屬氨基酸兩性離子配合物構象穩定。目前,Ca2+對Ala 手性轉變影響的研究鮮見報道,通常氣相環境有水分子(簇)存在,基于此并結合以往的研究[13,21],采用密度泛函理論對氣相丙氨酸Ca2+配合物的手性轉變機理及水分子的催化作用進行了研究,對進一步研究水溶劑環境下丙氨酸Ca2+配合物的手性轉變具有指導意義。
1 研究與計算方法
采用對處理含有金屬元素和弱鍵作用體系能量有較高精度的雜化泛函M06 方法[22],在6-31++G(d, p)[23]基組下,通過優化反應過程中單重態勢能面上的駐點結構(本工作體系的自旋態可能是1,3,5和7,計算表明單重態最穩定),計算了體系的自然鍵軌道(natural bond orbital,NBO)電荷。通過對過渡態[24]作的內稟反應坐標(IRC)計算[25],確認過渡態連接期望的穩定點。為得到較高水平的反應過程勢能面,同時兼顧計算成本,在M06/6-311++G(2df, pd)高角動量基組水平上計算單點能。總吉布斯自由能用Gtotal=Gtc+ESP計算(Gtc和ESP分別是吉布斯自由能熱校正和單點能)。S-Ala_1(丙氨酸穩定構象1)與Ca2+的配合物Ala_1·Ca2+記作S-A_1,S-A_1 在a 通道異構的第1 個S-型過渡態配合物記作S-T1_1a,第1 個S-型中間體配合物記作S-I1_1a;a 和b 通道公用的結構X 記作Xa(b),6 個水分子與SAz_1a(b)的Ca2+配位,同時2 個水分子簇(二聚水)與S-Az_1a(b)形成氫鍵,記作S-Az_1a(b)←6H2O·(H2O)2,其他體系用相似方法表示。采用Gaussian 09[26]程序在白城師范學院理論計算中心完成計算工作。
2 結果與討論
將文獻[8-11]的氣相Ala 分子兩種穩定構型作為初始猜測,優化的兩對手性對映體及Ca2+的幾何構型見圖1。計算表明,S-Ala_1 相對于S-Ala_2 的自由能為3.0 kJ·mol-1,S-Ala_1 的穩定性稍差。
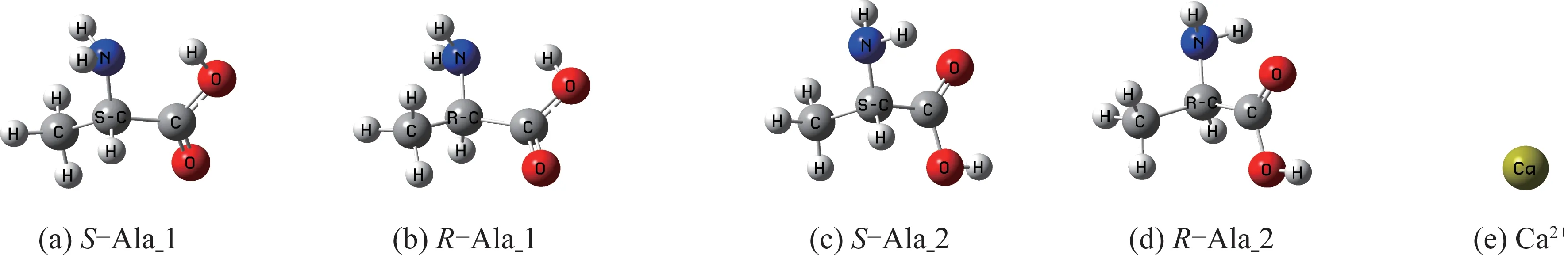
圖1 Ala 分子手性對映體及Ca2+的幾何構型Fig.1 Chiral enantiomer of Ala and geometric conformation of Ca2+
Ca2+可在不同位置與S-Ala_1 和S-Ala_2 的氮和氧配位形成配合物,優化得到的最穩定配合物SA_1 和S-A_2 的幾何構型見圖2。
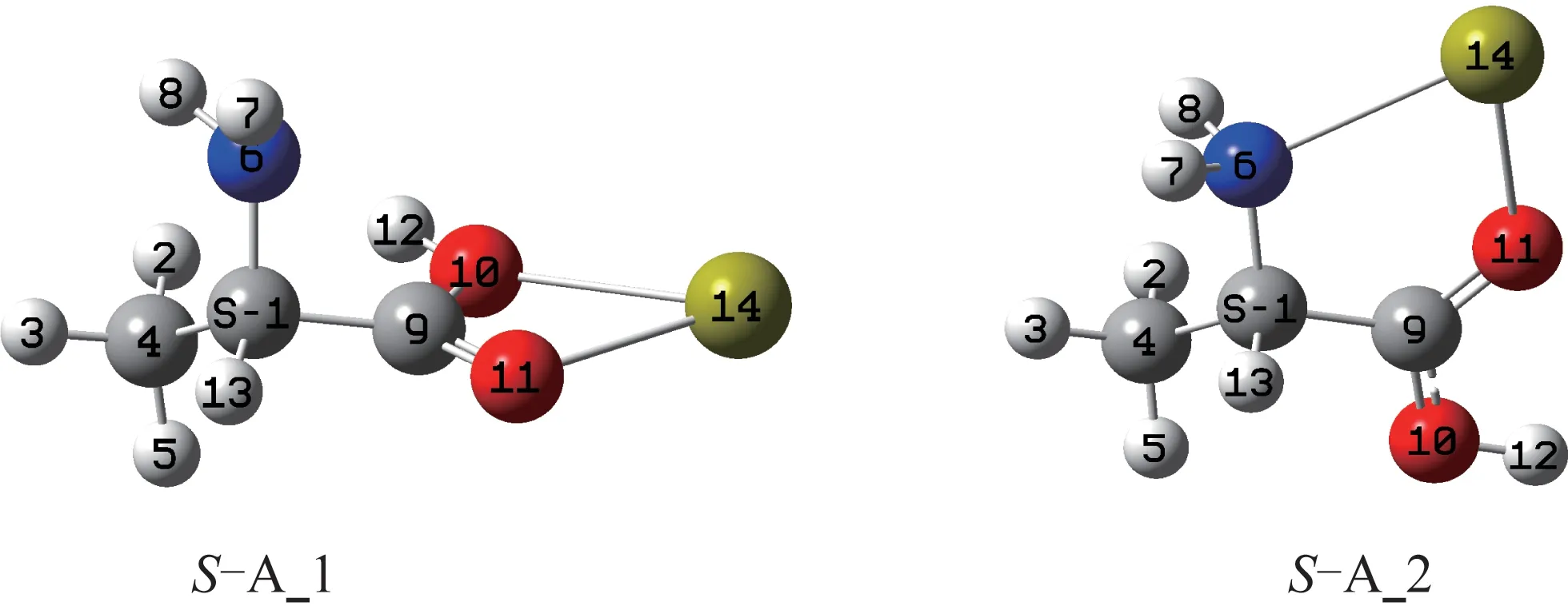
圖2 配合物S-A_1 和S-A_2 的幾何構型Fig.2 Geometric configuration of S-A_1 complex and S-A_2 complex
2.1 S-A_1 的手性轉變
S-A_1 的手性轉變反應有a 和b 2 個通道,反應歷程及過渡態矢量見圖3(第1 和第2 基元反應為共用基元),反應的吉布斯自由能勢能剖面見圖4。
第1 基元:反應物S-A_1,經1C—9C 鍵內旋轉的過渡態S-T1_1a(b),構象異構,形成兩性的中間體S-Az_1a(b)。從S-A_1 到S-T1_1a(b),1C—9C 鍵 沿 過渡態矢量的正方向向內旋轉,同時質子12H 從羧基氧10O 向氨基氮6N 遷移。二面角11O—9C—1C—6N 從94.1°變為97.5°,小角度的 C—C 鍵內旋轉只需很小的能量,S-T1_1a(b)產生的內稟能壘為3.7 kJ·mol-1。S-T1_1a(b)靠 近S-A_1,是 早 期 過 渡 態。該基元反應放熱148.5 kJ·mol-1,所放熱量可被后續的反應利用。兩性Ala 和Ca2+的配合物S-Az_1a(b)處于勢阱最深處,其構象遠比S-A_1 穩定。
第2 基元:S-Az_1a(b)經α-氫13H 向羰基氧11O遷移的過渡態T2_1a(b)構型異構,形成兩性的中間體配合物I2_1a(b)。從S-Az_1a(b)到T2_1a(b),13H 沿過渡態矢量正向遷移,1C—13H 鍵的鍵長從0.109 9 nm拉伸至0.151 9 nm,斷裂,二面角6N—1C—9C—4C從-123.9°變為-133.5°,C—H 鍵大幅拉伸斷裂及骨架的形變需要給予很高的能量,T2_1a(b)產生的內稟能壘為283.3 kJ·mol-1。較氣相環境下單體Ala在此過程的內稟能壘316.3 kJ·mol-1[5]有明顯下降,說明Ca2+對氫遷移反應有催化作用。由于Ca2+與兩性Ala 羧基的2 個氧同時配位,α-氫的NBO 電荷從0.251 e 增至0.307 e,11O 的NBO 電荷從-0.619 e 增至-0.813 e,導 致 羰基氧11O 對α-氫13H 的庫侖引力明顯增加。由于前一基元的反應放熱148.5 kJ·mol-1可 被利 用,因此S-Az_1a(b)越過T2_1a(b)需要的能量只有134.8 kJ·mol-1(283.3-148.5)。I2_1a(b)較S-A_1 在勢能面上的位置低、穩定性好,原因是I2_1a(b)的骨架原子基本共面,形成了共軛大π 鍵。
接下來的反應歷程分為a 和b 通道。
2.1.1 a 通道
第3 基元:I2_1a(b)經13H 在紙面內從11O 向1C遷移的過渡態T3_1a異構成中性的R-型產物R-Az_1a,S-A_1 實現手性轉變。從I2_1a(b)到T3_1a,11O—13H 鍵長從0.097 9 nm 拉伸至0.122 5 nm,斷裂,此處O—H 鍵拉伸幅度小于第2 基元的S-Az_1a(b)到T2_1a(b)的C—H 鍵拉伸幅度,又O—H 鍵較C—H 鍵易斷裂,因此T3_1a產生了181.4 kJ·mol-1的內稟能壘,較T2_1a(b)產生的內稟能壘小得多。結構分析表明,T3_1a和R-Az_1a分別與T2_1a(b)和S-Az_1a(b)鏡像對稱。至此,S-Az_1a(b)實現了手性對映體轉變。
第4 基元:R-Az_1a經1C—9C 鍵向內旋轉的過渡態R-T4_1a異構成兩性的產物配合物R-A_1a。從R-Az_1a到R-T4_1a,二面角11O—9C—1C—6N從171.9°變為-97.5°,同時6N—12H 鍵斷裂,12H 從6N 遷移到羰基氧10O,大角度的C—C 鍵向內旋轉以及12H 從6N 遷移到10O,需要一定的能量,RT4_1a產生的 內稟能 壘為152.2 kJ·mol-1。R-T4_1a靠近R-A_1a,為晚期過渡態。結構分析表明,R-A_1a與S-A_1 鏡像對稱。至此,S-A_1 實現了手性對映體轉變。
縱觀S-A_1 在a 通道整個反應歷程的駐點構象和勢能面,發現所有駐點構象及各個駐點的能量關于I2_1a(b)對稱,體現了Ala 分子手性對映體轉變的過程美和內在美!
2.1.2 b 通道
第3 基元:I2_1a(b)經8H 在紙面內從氨基向α-C遷移的過渡態T3_1b異構成R-型中間體R-I3_1b,S-A_1 實現手性轉變。從I2_1a(b)到T3_1b,6N—8H鍵長從0.102 9 拉伸至0.134 2 nm,斷裂,T3b_1 產生了199.3 kJ·mol-1的內稟能壘。較氣相環境下單體Ala 在此基元的內稟能壘106.5 kJ·mol-1[6]有大幅提升,說明Ca2+與羧基氧的配位對此基元反應有極強的負催化作用。原因是Ca2+與羰基氧配位導致I2_1a(b)8H 的NBO 電荷從0.436 e 增至0.485 e,6N 的NBO 電 荷 從0.795 e 增 至0.796 e,1C 的NBO 電 荷從-0.245 e 驟降至-0.081 e,導致6N 對8H 的庫侖引力增加,1C 對8H 的庫侖引力顯著減小。
第4 基元:R-I3b_1 經氨基氫左右翻轉的過渡態R-T4_1b異構成中間體配合物R-4_1b。從R-I3b_1到R-T4_1b, 二 面 角 7H—6N—1C—2H 從-147.7°變為-162.6°,非骨架二面角的小幅增加需要的能量很少,R-T4_1b產生的內稟能壘只有0.4 kJ·mol-1,此基元幾乎是無勢壘反應。
第5 基元:R-I4_1b經11O—14Ca 配位鍵斷裂和9C—10O—14Ca 剪式運動的過渡態R-T5_1b,異構成14Ca 與6N 和10O 配位的R-I5_1b。從R-I4_1b到R-T5_1b,較弱的11O—14Ca 配位鍵(鍵長0.256 1 nm)斷裂,9C—10O—14Ca 從106.1°到119.3°小幅剪式運動,所需能量較少,R-T5_1b產生的內稟能壘為2.2 kJ·mol-1。R-T5_1b靠近R-I4_1b,是11O—14Ca配位鍵斷裂和9C—10O—14Ca 剪式運動協同非同步的早期過渡態。
第6 基元:R-I5_1b經9C—11O 鍵向內旋轉的過渡態T6m_1b或T6n_1b,實現了13H 從羧基外側向羧基內側旋轉,異構成穩定的產物配合物R-A_2_1b。至此實現了S-Ala_1 向R-Ala_2 的異構。從R-I5_1b到T6m_1b,9C—11O 鍵 向 內 旋 轉81.0°需 要 的 能 量 較 少,從R-I5_1b到T6n_1b,9C—11O 鍵向內旋轉100.3°需要的能量也較少,因此,T6m_1b和T6n_1b產生的能壘較小,分別為37.7 和37.1 kJ·mol-1。
從圖4 中可以看出,S-A_1 手性轉變反應的a 通道具有優勢,反應的總包能壘為134.8 kJ·mol-1(來自于過渡態T2_1a(b)),遠低于氣相環境下單體Ala 手性轉變優勢通道的活化能266.0 kJ·mol-1[8](本文理論水平下的計算值為257.5 kJ·mol-1),說明Ca2+對S-Ala_1 的手性轉變具有極好的催化作用。134.8 kJ·mol-1的能壘在通常情況下可以極緩慢地越過,亦即配合物S-A_1 在該通道可以微量地消旋。b 通道的總包能壘為152.7 kJ·mol-1,越過此能壘的概率很小,S-A_1 在此通道只能痕量地消旋。從圖4 中還可看出,反應物中S-Az_1a(b)的分布遠高于S-A_1,產物中主要為R-Az_1a,而R-A_2_1b極少。此外,S-A_1 手性轉變反應的優勢通道a 是α-氫以羰基氧為橋遷移,而單體Ala 手性轉變的優勢通道是α-氫以氨基氮為橋遷移[8],說明Ca2+的存在改變了S-Ala_1 手性轉變反應的機理。
2.2 S-A_2 的手性轉變
S-A_2 的手性轉變反應有a、b、c 和d 4 個通道,其中,a 和b 通道共用第1 和第2 基元,c 和d 通道也共用第1 和第2 基元,反應歷程、駐點構象及過渡態矢量見圖5,反應的吉布斯自由能勢能剖面見圖6。
2.2.1 a 和b 通 道
第1 基元:S-A_2 經羧基內質子遷移的過渡態S-T1_2a(b),12H 沿 過 渡 態矢量的 負 方 向 從10O 遷 移到11O,異構成中間體配合物S-I1_2a(b)。從S-A_2到S-T1_2a(b),10O—12H 鍵的鍵 長 從0.097 7 nm 拉伸至0.135 0 nm,斷裂,10O—12H 鍵的大幅度拉伸使S-T1_2a(b)產生了188.2 kJ·mol-1的內稟能壘。這與氣相環境下單體Ala 在此基元反應的內稟能壘195.1 kJ·mol-1[5]相差無幾,說明Ca2+的存在對此基元反應的影響很小。
第2 基 元:S-I1_2a(b)經13H 從1C 向10O 遷移 的過渡態T2_2a(b),異構成中間體配合物I2_2a(b)。從SI1_2a(b)到T2_2a(b),1C—13H 鍵 的 鍵 長 從0.110 3 nm拉伸至0.128 9 nm,斷裂,骨架二面角6N—1C—9C—4C 從-126.8°變為-140.0°,C—H 鍵的斷裂及骨架的形變需要較高的能量,T2_2a(b)產生了300.0 kJ·mol-1的內稟能壘。較氣相環境下單體Ala 在此基元反應的能壘316.3 kJ·mol-1[5]略有降低,說明Ca2+對此基元反應的催化作用很小。
2.2.2 a 通道
第3 基元:I2_2a(b)經與T2_2a(b)鏡像對稱的過渡態T3_2a,實現了13H 在紙面內側從10O 向α-碳1C的遷移,異構成中間體配合物R-3a_2。至此,S-A_2實現了手性轉變。從I2_2a(b)到T3_2a,10O—13H 鍵的鍵長從0.096 9 nm 拉伸至0.128 6 nm,斷裂,T3_2a產生的內稟能壘為258.0 kJ·mol-1。此基元與2.1 節中a 通道的第3 基元反應相似,不再贅述。
第4 基元:R-I3_2a經過與S-T1_2a(b)鏡像對稱的過渡態R-TS4_2a,實現12H 在羧基內的回遷,異構成產物配合物R-A_2a。結構分析表明,R-A_2a與S-A_2 鏡像對稱。至此,R-A_2a實現了手性對映體轉變。從R-I3_2a到R-TS4_2a的過程中,11O—12H 鍵的鍵長從0.097 9 nm 拉伸至0.124 9 nm,斷裂,R-TS4_2a產 生 的 能 壘 為102.3 kJ·mol-1。此 基元反應的能壘小于第1 基元反應,原因是此基元反應比第1 基元反應O—H 鍵的拉伸幅度小。
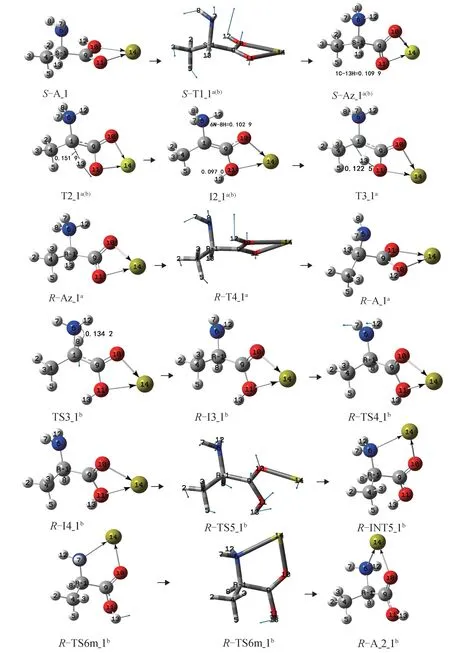
圖3 Ca2+催化S-Ala_1 的手性轉變歷程(鍵長單位:nm)Fig.3 Reaction process of chiral transition of S-Ala_1 catalyzed by Ca2+(bond length unit:nm)
縱觀S-A_2 在a 通道反應歷程的駐點構象和勢能面,發現所有駐點構象及各個駐點的能量關于I2_2a(b)對稱,體現了Ala 分子手性對映體轉變的過程美和內在美!
2.2.3 b 通道
第3 基元:I2_2a(b)經8H 在紙面內從氨基向α-碳遷移的過渡態T3_2b,異構成R-型中間體R-I3_2b,S-A_2 實現了手性轉變。從I2_2a(b)到T3_2b,6N—8H 鍵的鍵長從0.102 6 nm 拉伸至0.142 2 nm,斷裂,T3b_2 產生的內稟能壘為198.5 kJ·mol-1。較氣相環境下單體Ala 在此基元反應的內稟能壘106.5 kJ·mol-1[6]有大幅提升,Ca2+與羧基氧的配位對此基元有極強的負催化作用。原因與a 通道第3 基元相似,不再贅述。
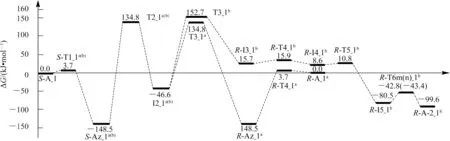
圖4 Ca2+催化S-Ala_1 手性轉變反應的吉布斯自由能勢能剖面Fig.4 Gibbs free energy surfaces of chiral transformation reaction of S-Ala_1 catalyzed by Ca2+
第4 基元:R-3_2b經11O—9C 鍵俯視順時針或逆時針旋轉的過渡態R-TS4m_2b或R-TS4n_2b,異構成中間體R-I4m_2b或R-I4n_2b。從R-I3_2b到R-TS4m_2b,二面角12H—11O—9C—10O 從0.2°變為-96.1°,二面角11O—9C—1C—6N 從2.4°變為-72.7°,二面角14Ca—6N—1C—9C 從3.6°變為128.5°,11O—9C 鍵俯視順時針旋轉與9C—1C 鍵左視逆時針旋轉及1C—6N 鍵俯視逆時針旋轉協同進行。3 個化學鍵向內旋轉使R-TS4m_2b產生了60.9 kJ·mol-1的 內 稟 能 壘。同 樣,R-TS4n_2b產 生了70.8 kJ·mol-1的內稟能壘。
第5 基元:R-I4m_2b經12H 從11O 向6N 遷移的過渡態R-T5m_2b,異構成R-I5_2b(R-INT5_1b)。從R-I4m_2b到R-T5m_2b,11O—12H 鍵的鍵長從0.101 6 nm 拉伸至0.106 9 nm,O—H 鍵小幅拉伸需要的能量很少,R-T5m_2b產生的內稟能壘只有0.3 kJ·mol-1。同 樣,R-T5n_2b產 生 的 內 稟 能 壘 只 有1.8 kJ·mol-1。 結 構 分 析 表 明,R-I5_2b同 于RINT5_1b,R-I5_2b的 異 構 體 也 同 于R-INT5_1b(見2.1.2 節b 通道的第6 基元)。
2.2.4 c 和d 通 道
第1 基元:S-A_2 經9C—10O 鍵向內旋轉的過渡態S-T1m_2c(d)或S-T1n_2c(d),羧基從順式平面結構改變為反式平面結構,異構成中間體S-I1_2c(d)。從S-A_2 到S-T1m_2c(d),9C—10O 鍵沿S-T1m_2c(d)虛頻的負向旋轉97.0°,S-T1m_2c(d)產生的內稟能壘為57.2 kJ·mol-1。同樣,S-T1n_2c(d)產生的內稟能壘為57.7 kJ·mol-1。
第2 基元:S-I1_2c(d)經14Ca—6N 配位鍵斷裂與14Ca—11O—9C 做沿虛頻正向剪式運動協同進行的過渡態S-T2_2c(d)異構成S-I2_2c(d)。從S-I1_2c(d)到S-T2_2c(d),14Ca—6N 鍵 斷 裂 且14Ca 從 羧 基 的 外 側擺動到內側,需要較多能量,S-T2_2c(d)產生了92.6 kJ·mol-1的內稟能壘。
2.2.5 c 通道
第3 基元:S-I2_2c(d)經氨基左右翻轉的過渡態S-T3_2c,異 構 成S-I3_2c。從S-I2_2c(d)到S-T3_2c,二面角7H—6N—1C—8H 從131.5°變為-162.6°,非骨架的形變所需能量很少,S-I3_2c產生的內稟能壘為7.8 kJ·mol-1。
第4 基元:S-I3_2c經α-氫13H 從α-碳向氨基氮遷移的過渡態T4_2c,異構成I4_2c[I2_1a(b)]。從S-I3_2c到T4_2c,1C—13H 鍵 長從0.110 5 nm 拉伸至0.123 3 nm,斷裂,C—H 鍵大幅拉伸需要的能量較高,T4_2c產生了136.6 kJ·mol-1的內稟能壘。該能壘較氣相環境下單體Ala_2 在此基元的能壘248.2 kJ·mol-1[10]大幅降低(本文理論值為248.2 kJ·mol-1,文獻[10]的理論值約為260.3 kJ·mol-1),說明Ca2+與羧基氧的配位對此基元有極好的催化作用。原因是Ca2+與羧基氧的配位導致α-氫的紅外伸縮振動頻率從3 052.1 Hz 降至2 997.7 Hz;1C—13H鍵長從0.109 9 nm 拉伸至0.110 5 nm,1C—13H 鍵被活化。結構分析表明,I4_2c同于I2_1a(b)。I4_2c繼續的異構同于I2_1a(b)之后的異構。
2.2.6 d 通道
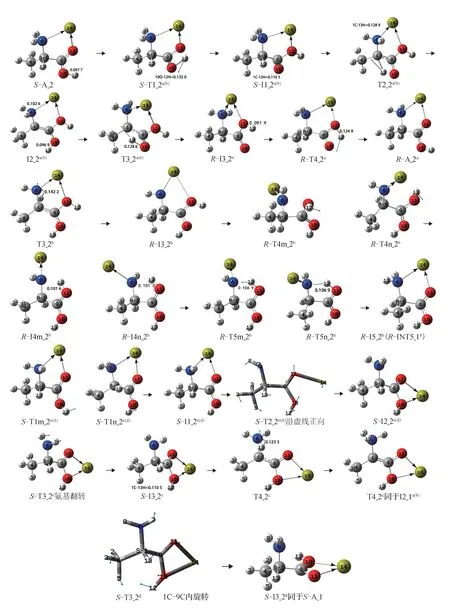
圖5 Ca2+催化S-Ala_2 的手性轉變歷程(鍵長單位:nm)Fig.5 Reaction process of chiral transition of S-Ala_2 catalyzed by Ca2+(bond length unit:nm)
第3 基元,S-2_2c(d)經1C—9C 鍵沿過渡態矢量的負方向向內旋轉的過渡態S-T3_2d異構成S-I3_2d。從S-I2_2c(d)到S-T3_2d,二面角10O—9C—1C—6N 從153.9°變為-173.6°,S-T3_2d產生的內稟能壘 為6.9 kJ·mol-1。結 構 分 析 表 明,S-I3_2d同 于SA_1。至此,完成了S-A_2 向S-A_1 的異構。S-I3_2d繼續的異構同于S-A_1 的異構。
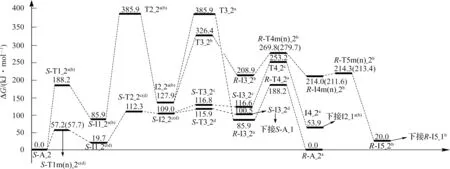
圖6 Ca2+催化S-Ala_2 手性轉變反應的吉布斯自由能勢能剖面Fig.6 Gibbs free energy surfaces of chiral transformation reaction of S-Ala_2 catalyzed by Ca2+
由圖6 并結合圖4 可知,S-A_2 手性轉變反應的d 通道具有優勢,總包能壘為(100.5+134.8)kJ·mol-1=235.3 kJ·mol-1(來自于過渡態T2_1a(b)),低于單體Ala_2 手性轉變優勢通道的總包能壘249.3 kJ·mol-1(本文理論值為249.3 kJ·mol-1,文獻[10]的理論值約為260.1 kJ·mol-1),說明Ca2+對SAla_2 的手性轉變具有催化作用。但235.3 kJ·mol-1的能壘已經很高,通常難以逾越,說明S-Ala_2 的Ca2+配合物可以很好地保持其手性特征。從圖6 中還 可 以 看 出,S-A_2 比S-A_1 穩 定 得 多,S-A_1 很容易 向S-A_2 異 構,并且S-1_2c(d)與S-A_2 共存,只是S-A_2 的分布要高得多。由圖4 和圖6 知,在SAla 的Ca2+配合物中,兩性的S-Az_1a(b)最穩定,然而在氣相環境下兩性的Ala 不存在[27]。
S-Ala_1 相對于S-Ala_2 的吉布斯自由能為3.0 kJ·mol-1,對于氣相環境下的S-Ala,S-Ala_1 的分布較S-Ala_2 少得多。由圖4 和圖6 知,Ca2+的存在使S-Ala 形成大量的S-A_2 和S-Az_1a(b),旋光異構后會得到少量的R-Az_1a和微量的R-A_2_1b。
2.3 水汽環境下水分子對S-A_1 和S-A_2 手性轉變的影響
由前述討論可知,S-A_1 和S-A_2 手性轉變反應優勢通道的總包能壘均來自過渡態T2_1a(b),因此,為節省篇幅,又不失對問題討論的完整性,只討論水分子對S-A_1 手性轉變優勢通道的作用。
水分子對S-A_1 的作用有2 種,一是水分子會與20Ca 形成配位鍵,二是水分子(簇)會與S-A_1 的氫及氧形成氫鍵。由于配位鍵強于氫鍵,因此實際情況應是水分子與20Ca 充分配位后,再與S-A_1 的氫及氧形成氫鍵。參與配位的水分子個數取最大值6(Ca2+與O 最多可以八配位,在S-A_1 中Ca2+已經與羰基氧二配位)。計算結果表明,6 個水分子與SA_1 的20Ca 配位后,1C—9C 鍵向內旋轉,形成與6N—1C—9C—10O—11O—20Ca 共 面 的S-Ala_1和6 個水分子與20Ca 配位的配合物S-A_1←6H2O,故本節討論的第1 基元反應是羧羥基氫從羧羥基氧向氨基氮遷移。
田子德等[11]的研究表明,不參與質子轉移的、與底物形成氫鍵的水分子,對質子轉移能壘影響很小;文獻[9, 28-29]認為,對于質子從丙氨酸的α-碳向羰基氧遷移,二聚水和三聚水的催化作用相差很小,但遠大于單個水分子;劉芳等[30]的研究表明,質子從羧基向氨基遷移水分子作橋能壘改變很小,且此基元不是決速步。基于上述原因,為使問題簡便且節省篇幅,同時為了得到連續的反應過程勢能面,在S-A_1←6H2O 的前面植入二聚水與α-氫12H 和羰基氧10O 形成氫鍵,優化得到反應物S-A_1←6H2O·(H2O)2m(m表示(H2O)2在S-A_1 之前,相似之處不再說明),見圖7。對于第1 基元,羧羥基氫從羧羥基氧向氨基氮遷移時不考慮水分子作媒介;對于第2 基元,只討論二聚水作為12H 從1C 向10O 遷移媒介的情況;對于第3 基元,只討論二聚水與I2_1a(b)的羧羥基氫16H 和α-碳1C 形成氫鍵,作為16H 從10O 向 1C 遷 移 媒 介 的 情 況 。S-A_1←6H2O·(H2O)2m的手性轉變反應歷程及過渡態見圖7,反應的吉布斯自由能勢能剖面見圖8。
第1 基元:S-A_1←6H2O·(H2O)2m經過渡態ST1_1a(b)←6H2O·(H2O)2m,實現了13H 從11O 向6N 遷移,異構成S-Az_1a(b)←6H2O·(H2O)2m。從S-A_1←6H2O·(H2O)2m到S-T1_1a(b)←6H2O·(H2O)2m,11O—13H 鍵長從0.101 2 nm 拉伸至0.127 7 nm ,斷裂,O—H 鍵較小的拉伸需要的能量不高。又二面角6N—13H—11O 為139.1°,6N—13H—11O 形成了中等強度的氫鍵,結構分析表明,6N—13H—11O—9C—1C 基本共面,過渡態S-T1_1a(b)←6H2O·(H2O)2m的五元環結構穩定。因此S-T1_1a(b)←6H2O·(H2O)2m產生的內稟能壘很低,只有12.3 kJ·mol-1。
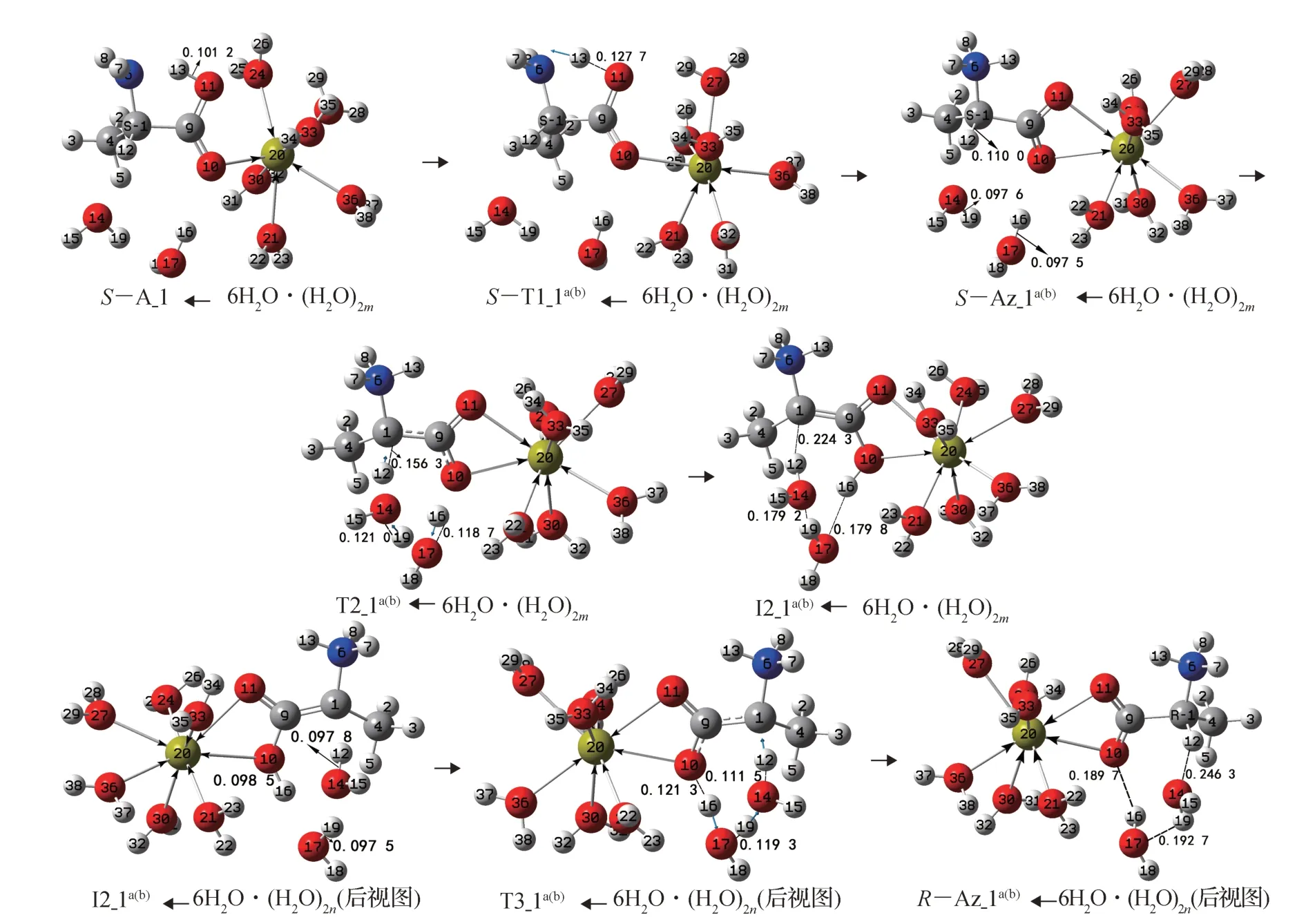
圖7 水分子作用下的S-A_1 手性轉變反應過程Fig.7 Chiral transition reaction process of S-A_1 under the action of water molecules

圖8 水分子作用下的S-A_1 手性轉變反應過程的吉布斯自由能勢能面Fig.8 Gibbs free energy surfaces of S-A_1 chiral transition reaction process under the action of water molecules
第2 基元:S-Az_1a(b)←6H2O·(H2O)2m經過渡態T2_1a(b)←6H2O·(H2O)2m,實現了質子從α-碳1C 向羰基氧10O 凈遷移,異構成中間體I2_1a(b)←6H2O·(H2O)2m。從S-Az_1a(b)←6H2O·(H2O)2m到T2_1a(b)←6H2O·(H2O)2m,1C—12H、14O—19H 和17O—16H鍵長從0.110 0 ,0.097 6 和0.097 5 nm 分別拉伸至0.156 3 ,0.121 0 和0.118 7 nm,斷裂;骨架二面角6N—1C—9C—4C 從-123.7°變為-128.9°。3 個化學鍵的拉伸斷裂,其中還有一個C—H 鍵斷裂,需要一定的能量,但結構分析表明,T2_1a(b)←6H2O·(H2O)2m的氫鍵角1C—12H—14O、14O—19H—17O和17O—16H—10O 分別為163.2°,161.6°和164.1°,接近平角,3 個氫鍵較強,具有八元環結構的過渡態T2_1a(b)←6H2O·(H2O)2m較 穩 定。因 此,T2_1a(b)←6H2O·(H2O)2m產生的內稟能壘不會很高,為159.3 kJ·mol-1,遠低于2.1 節中沒有水分子存在時T2_1a(b)產生的內稟能壘283.3 kJ·mol-1,說明水分子簇對此基元反應起了極好的催化作用。
第3 基元:I2_1a(b)←6H2O·(H2O)2n(n表示(H2O)2在I2_1a(b)之后,相似之處不再說明)經過渡態T3_1a(b)←6H2O·(H2O)2n,實現了質子在紙面內從10O 向1C的凈遷移,異構成R-型產物R-Az_1a(b)←6H2O·(H2O)2n,S-Az_1a(b)←6H2O 實現了手性對映體轉變。從I2_1a(b)←6H2O·(H2O)2n到T3_1a(b)←6H2O·(H2O)2n,10O—16H、17O—19H 和14O—12H 鍵 的鍵長從0.098 5 ,0.097 5 和0.097 8 nm 分別拉伸至0.121 3,0.119 3 和0.111 5 nm,斷裂,骨架二面角6N—1C—9C—4C 從150.7°變為129.0°。3 個化學鍵的拉伸斷裂及骨架不小的形變需要一定的能量。結構分析表明,T3_1a(b)←6H2O·(H2O)2n的氫鍵角10O—16H—17O、17O—19H—14O 和14O—12H—1C 分 別 為164.1°,161.6°和163.2°,接近平角,3 個氫鍵較強,八元環結構的過渡態T2_1a(b)←6H2O·(H2O)2m較穩定,并 且 從I2_1a(b)←6H2O·(H2O)2n到T3_1a(b)←6H2O·(H2O)2n斷裂的都是O—H 鍵。因此T2_1a(b)←6H2O·(H2O)2m產生的內稟能壘不高,低于T2_1a(b)←6H2O·(H2O)2m產 生 的 內 稟 能 壘,為50.8 kJ·mol-1,遠 低 于2.1 節中沒有水分子存在時T3_1a(b)產生的內稟能壘181.4 kJ·mol-1,說明水分子(簇)對此基元反應起了極好的催化作用。
第4 基元:R-Az_1a(b)←6H2O·(H2O)2n經與第1基元反應相似的逆過程,可以異構成R-A_1←6H2O·(H2O)2n,反應能壘為130.8 kJ·mol-1。此基元反應很難進行,R-Az_1a(b)←6H2O·(H2O)2n是該反應通道的主要產物,為節省篇幅,省略了該基元的反應歷程及勢能。
從圖8 中可以看出,在水分子作用下,S-A_1 手性轉變反應優勢通道a 的總包能壘為40.8 kJ·mol-1;由圖8 并結合圖6 發現,在水分子作用下,S-A_2 手性轉變反應優勢通道d 的總包能壘為141.3 kJ·mol-1(100.5+40.8=141.3)。通常情況下40.8 kJ·mol-1的能壘容易越過,141.3 kJ·mol-1的能壘可以緩慢地越過,亦即在水汽環境下,S-A_1 可以較快地消旋,S-A_2 可以緩慢地消旋。
3 結 論
在M06/6-311++G(2df, pd)//M06/6-31++G(d,p)雙水平下,研究了Ca2+與2 種穩定構型Ala 配合物的手性轉變及水分子的作用,得到以下結論:
3.1S-A_1 手性轉變優勢通道a 的總包能壘為134.8 kJ·mol-1,遠低于Ala_1 手性轉變的總包能壘257.5 kJ·mol-1;S-A_2 手性轉變優勢通道d 的總包能壘為235.3 kJ·mol-1,低于Ala_2 手性轉變的總包能壘249.3 kJ·mol-1,Ca2+對Ala_1 和Ala_2 的手性轉變分別具有較強和較弱的催化作用。在氣相環境下,Ala_2 的分布遠高于Ala_1,Ca2+與Ala 的配合物只能微量消旋。
3.2 水分子(簇)的催化使S-A_1 和S-A_2 手性轉變反應優勢通道的總包能壘分別降至40.8 和141.3 kJ·mol-1,在水汽環境下,Ala_1 與Ca2+的配合物可以很快消旋,Ala_2 與Ca2+的配合物可以緩慢消旋。
3.3 Ca2+對Ala 手性轉變反應的某些基元有正催化作用,對另一些基元則有負催化作用,對整個手性轉變過程起正催化作用,Ca2+的存在改變了Ala 手性轉變的反應機理。
3.4 Ca2+與Ala 配位后,使不同構象Ala 的相對穩定性發生了顯著變化。
