不同濃度米諾環素對人膠質瘤細胞增殖、自噬、凋亡的影響及其機制
2020-09-22 00:57:20韓利民嚴婉約李巧巧李科劉麗趙海龍
天津醫藥 2020年9期
韓利民,嚴婉約,李巧巧,李科,劉麗,趙海龍△
腦膠質瘤是最常見的顱內原發性腫瘤,在我國發病率約為3.2/10 萬,約占中樞神經系統原發性腫瘤的24.7%[1]。目前膠質瘤的標準治療方案主要是以手術切除為主,同步進行放化療等綜合治療,但效果有限,患者的總生存期未明顯延長[2]。米諾環素是第二代四環素類抗生素,它能與轉運RNA(tRNA)結合從而達到抑菌效果。近年來研究發現米諾環素除了自身抗炎作用外,還有干擾腫瘤微環境[3]、心臟保護[4]、神經保護[5]等作用,并能通過提高自噬、抑制細胞外基質降解蛋白酶等途徑抑制膠質瘤的生長[6-7]。本課題組前期研究證實高濃度米諾環素(50~200 μmol/L)能誘導膠質瘤細胞自噬和凋亡[8],但對于此過程中自噬和凋亡發生的具體機制尚不清楚。為驗證較低濃度的米諾環素是否具有同樣作用,本研究分別采用5和10 μmol/L的米諾環素處理人膠質瘤細胞U87 和LN229,并進一步探究其具體作用機制。
1 材料與方法
1.1 主要材料 米諾環素購自MCE 公司。DMEM/F12 培養基、胎牛血清購自Gibco 公司;臺盼藍染色液、MTT 染色液購自碧云天生物技術公司;SYBR 熒光定量試劑盒購自Invitrogen公司;RNA提取和逆轉錄試劑盒購自Premega公司;Lipofectamine 2000轉染試劑盒購自Thermo公司。兔源沉默信息調節因子2相關酶1(SIRT1)抗體、哺乳動物雷帕霉素靶蛋白(mTOR)抗體、B 淋巴細胞瘤-2(Bcl-2)抗體、剪切活化后的含半胱氨酸的天冬氨酸蛋白水解酶-3(Cleaved-Caspase-3)抗體、磷酸化p70 核糖體蛋白S6 激酶(蘇氨酸389 位點)[phospho-p70 S6 Kinase,p-p70s6k(Thr389)]抗體、p70 核糖體蛋白S6 激酶(p70s6k)抗體、自噬基因相關蛋白5(Atg5)抗體、微管相關蛋白l 輕鏈3B 亞基(LC3B)抗體、信號傳導及轉錄激活蛋白3(STAT3)抗體均購自英國Abcam 公司,磷酸化AMP 依賴的蛋白激酶α亞基(蘇氨酸172位點)[phospho-adenosine 5-monophosphate-activated protein kinase-alpha,p-AMPKα(Thr172)]抗體、AMP 依賴的蛋白激酶 α 亞基(AMPKα)抗體購自Cell Signaling Technology 公司。β肌動蛋白(β-actin)抗體購自武漢三鷹生物技術有限公司。辣根過氧化物酶標記山羊抗兔二抗、山羊源抗兔熒光二抗分別購自KPL公司和Life Technologies。pLKO.1-puro質粒購自北京中源合聚生物科技有限公司,293FT由遵義醫科大學生命科學研究院饋贈。
1.2 不同濃度米諾環素處理后U87 和LN229 細胞增殖和凋亡相關指標的檢測
1.2.1 細胞培養及分組 U87和LN229細胞株由遵義醫科大學病理生理學教研室常規保存。2種細胞培養于含10%胎牛血清、100 mg/L鏈霉素和1×105U/L青霉素的DMEM/F12培養基中,置于5%CO2、37 ℃環境中常規傳代培養,每1~2 d觀察生長情況。細胞分組:2 種膠質瘤細胞分別按米諾環素處理濃度分為3組:對照組(米諾環素0 μmol/L,以DMSO代替),5 μmol/L米諾環素組和10 μmol/L米諾環素組。各組細胞均在接種24 h后,分別加入相應濃度米諾環素培養72 h。
1.2.2 MTT 法檢測細胞增殖能力 取對數生長期的3 組細胞,胰酶消化后,按1×104個/mL 重懸細胞,200 μL/孔接種于96孔板。培養24、48和72 h后添加MTT 溶液(20 μL/孔),繼續培養4 h,酶標儀讀取各組細胞560 nm 處的光密度(OD)值,以觀察細胞增殖能力變化。
1.2.3 免疫熒光染色檢測LC3B 的表達 取對數生長期的3組細胞,在含細胞爬片的12孔板上調整細胞密度為1×105/mL。培養72 h 后依次進行固定(4%多聚甲醛)、通透(室溫下0.3%TritonX 100 處理5 min)、封閉(5%BSA 處理2 h),加入一抗兔源LC3B(1∶1 000 稀釋)4 ℃后過夜處理,接著加入山羊源抗兔熒光二抗(1∶5 000稀釋)并在室溫下孵育2 h,然后用Hoechst33342染色液進行染色,15 min后用PBS漂洗干凈,最后在激光共聚焦顯微鏡下觀察爬片。
1.2.4 Western blot 檢測自噬和凋亡相關蛋白的表達 米諾環素處理72 h后收集各組細胞并提取總蛋白,BCA法測定蛋白濃度后各組以30 μg 上樣量進行10%SDS-PAGE 電泳、濕轉PVDF 膜,然后常溫下使用5%BSA 封閉1 h,添加SIRT1、mTOR、Bcl-2、Cleaved-Caspase-3、p-p70s6k、p70s6k、Atg5、LC3B、STAT3、p-AMPKα、AMPKα、β-actin一抗(均為1∶1 000稀釋)于4 ℃孵育過夜,次日TBST 震蕩洗滌3 次后與辣根過氧化物酶標記山羊抗兔二抗(1∶10 000 稀釋)孵育1 h,TBST洗滌3次后進行ECL化學發光顯影,所有蛋白條帶灰度值均使用Image J 進行對比,以β-actin作為內參蛋白。
1.3 shRNA 干擾SIRT1 表達后觀察米諾環素對U87 和LN229細胞自噬相關分子表達的影響
1.3.1 SIRT1-shRNA 載體構建 在GenBank 中查找SIRT1基因的mRNA 序列(GenBank 登錄號:NM_001142498.1),利用Sigma 在線軟件設計2 對干擾序列(表1)。SIRT1-shRNA引物由華大公司合成,將合成的正反義Oligo DNA經過變性、退火后形成shRNA 模板,與pLKO.1-puro 質粒載體連接,最后經篩選、酶切鑒定后再經華大公司測序。

Tab.1 Sequences of SIRT1 interference表1 SIRT1干擾序列
1.3.2 慢病毒包裝和轉染 消化、計數293FT 細胞,并將細胞稀釋在慢病毒培養基中。在6孔板中加入DNA-脂質體復合物、慢病毒培養基、293FT細胞懸液,輕輕混勻后37 ℃培養過夜。次日換液,加入慢病毒培養基2 mL繼續培養48 h。將待轉染的細胞(U87、LN229)用不含抗生素培養基培養,收集293FT細胞產生的病毒上清液,向待轉染的細胞中加入1 mL慢病毒培養基和1 mL 病毒上清液,最后用抗性藥物篩選穩定細胞株。選擇SIRT1 表達干擾效果較好的穩定細胞株進行重新接種,24 h后分別加入相應濃度米諾環素培養至72 h。細胞分為4組:Control組、shSIRT1組、shSIRT1+5 μmol/L米諾環素組和 shSIRT1+10 μmol/L 米諾環素組,其中 Control 組轉染靶向GFP的shRNA質粒并加入等量DMSO作為對照。
1.3.3 qPCR 檢測shRNA 干擾后SIRT1的表達 分別收集分組處理72 h后的U87和LN229細胞,進行細胞裂解后根據說明書提取總RNA,逆轉錄為cDNA 后進行qPCR 檢測。反應條件:95 ℃預變性3 min;95 ℃變性5 s,60 ℃退火延伸30 s,40個循環,同時收集熒光信號,以β-actin為內參。每組重復操作 3 次,SIRT1 和 β-actin 的 mRNA 表達量采用 2-ΔΔCt法計算,具體序列見表2。

Tab.2 Sequences of the primers in qPCR表2 qPCR引物序列
1.3.4 Western blot 檢測SIRT1敲低后細胞自噬相關蛋白的表達 細胞收集、蛋白提取、電泳、轉膜、抗體孵育、指標分析等步驟同1.2.4。
1.4 統計學方法 采用SPSS 18.0 軟件進行統計學分析,計量資料以均數±標準差()表示,多組間均數比較用單因素方差分析,組間多重比較采用LSD-t檢驗,P<0.05為差異有統計學意義。
2 結果
2.1 不同濃度米諾環素對膠質瘤細胞增殖能力的影響 MTT 結果顯示,與對照組相比,5 μmol/L 和10 μmol/L 米諾環素處理后 U87 在 48 h 增殖水平顯著下降(P<0.05),而LN229在72 h增殖水平顯著下降(P<0.05),見圖1。
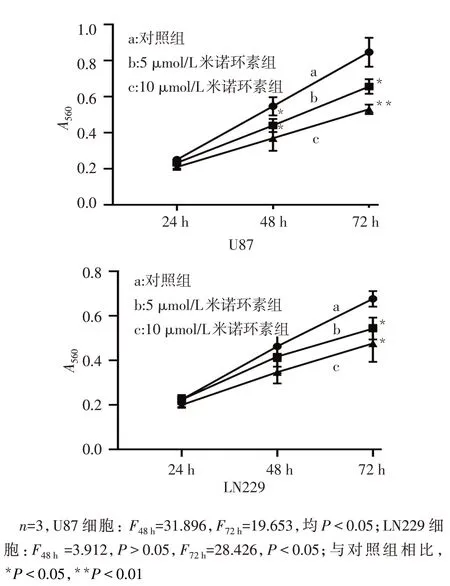
Fig.1 Proliferation of U87 and LN229 cells treated with different concentrations of minocycline within 72 h圖1 不同濃度米諾環素處理后U87和LN229細胞72 h內增殖情況
2.2 不同濃度米諾環素對U87 和LN229 細胞自噬水平的影響 免疫熒光染色結果顯示,與對照組相比,5 μmol/L和10 μmol/L米諾環素組細胞自噬標記蛋白LC3B 熒光斑點比例明顯增多(P<0.01),并呈劑量依賴性升高,見圖2、表3。Western blot 結果顯示,與對照組相比,5 μmol/L 和10 μmol/L 米諾環素組自噬抑制蛋白mTOR 明顯減少,而自噬蛋白Atg5明顯增加,且伴有AMPKα磷酸化水平和SIRT1表達水平顯著增加(P<0.01),見圖3、表4。
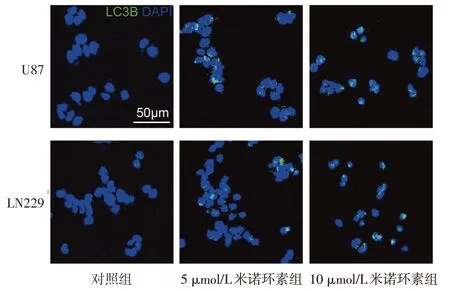
Fig.2 Immunofluorescence assayresults(LC3B)in U87 and LN229 cells treated with different concentrations of minocycline圖2 不同濃度米諾環素處理后U87和LN229細胞胞漿內自噬標記蛋白LC3B顯影情況
2.3 不同濃度米諾環素對U87 和LN229 細胞凋亡相關蛋白表達的影響 Western blot 結果顯示,與對照組相比,5 μmol/L 米諾環素處理后僅LN229 細胞Cleaved Caspase-3 表達下降(P<0.05),其他蛋白表達未見明顯變化(P>0.05),而10 μmol/L 米諾環素處理后,2 種細胞中 p-p70s6k 和 Bcl-2 表達明顯減少,Cleaved Caspase-3 蛋白明顯增加(P<0.05),見圖4、表5。
Tab.3 Quantification of LC3B punctae in U87 and LN229 cells after treatment with different concentrations of minocycline表3 3組不同濃度米諾環素處理后的LC3B斑點數/細胞數變化 (n=3,)

Tab.3 Quantification of LC3B punctae in U87 and LN229 cells after treatment with different concentrations of minocycline表3 3組不同濃度米諾環素處理后的LC3B斑點數/細胞數變化 (n=3,)
**P<0.01;a與對照組相比,b與5 μmol/L米諾環素組相比,P<0.05
組別對照組5 μmol/L米諾環素組10 μmol/L米諾環素組F LC3B(%)U87 0.19±0.17 44.67±7.51a 71.33±5.86ab 128.173**LN229 0.83±0.76 36.00±11.00a 51.67±6.51ab 37.209**
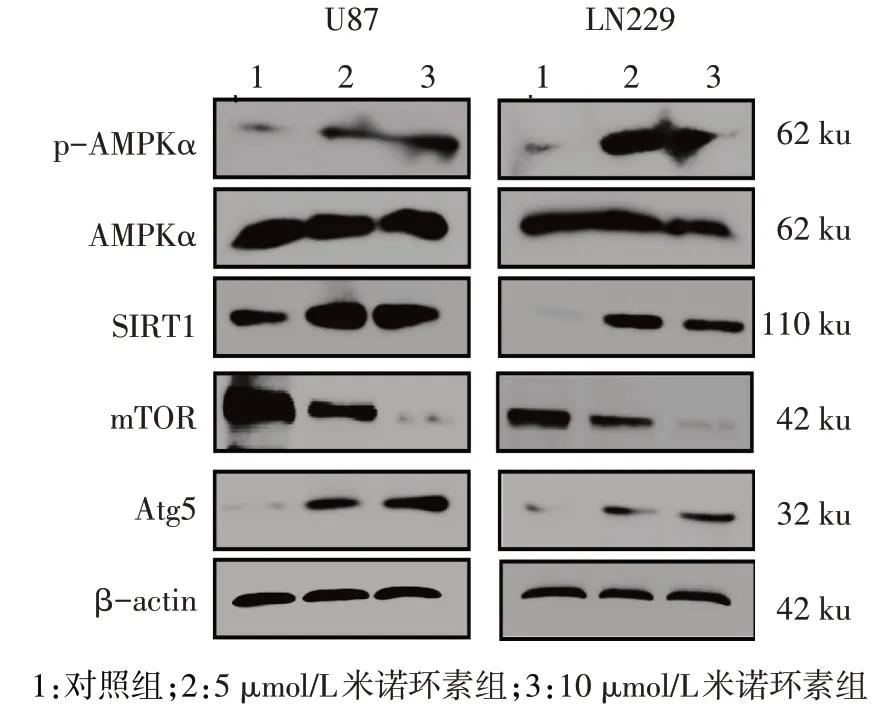
Fig.3 Western blot results of autophagy-associated proteins in U87 and LN229 cells treated with different concentrations of minocycline圖3 不同濃度米諾環素處理后U87和LN229細胞自噬相關蛋白的表達變化

Fig.4 Western blot results of apoptotic proteins in U87 and LN229 cells treated with different concentrations of minocycline圖4 U87和LN229細胞在不同濃度的米諾環素處理后凋亡相關蛋白水平的變化
2.4 SIRT1敲低后米諾環素對U87和LN229細胞自噬水平的影響 本研究構建的2 種靶向SIRT1 的shRNA載體(shSIRT1-1,shSIRT1-2),通過慢病毒轉染 U87 和 LN229 細胞后,經 qPCR 和 Western blot 確認shSIRT-1 的敲低效果相對較好(圖5),故選取shSIRT1-1 轉染的U87 和LN229 細胞進行下一步實驗。SIRT1 表達敲低后,shSIRT1 組 mTOR 和 LC3B表達水平較對照組明顯下降;而經過米諾環素處理后,mTOR 的表達出現明顯升高,而LC3B 表達水平僅部分恢復,見圖6、表6。
Tab.4 The expression levels of autophagy-associated proteins inU87 and LN229 cells after treatment with different concentrations of minocycline表4 3組不同濃度米諾環素處理后的U87和LN229細胞自噬相關蛋白水平變化 (n=3,)
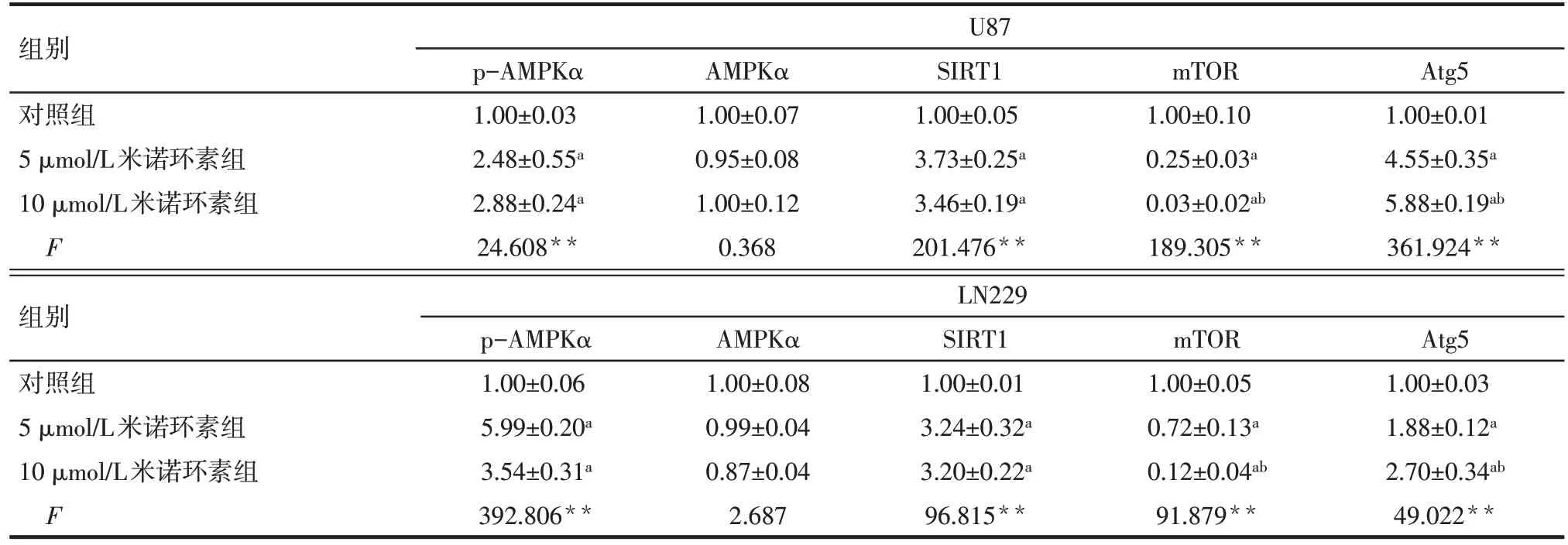
Tab.4 The expression levels of autophagy-associated proteins inU87 and LN229 cells after treatment with different concentrations of minocycline表4 3組不同濃度米諾環素處理后的U87和LN229細胞自噬相關蛋白水平變化 (n=3,)
**P<0.01;a與對照組相比,b與5 μmol/L米諾環素組相比,P<0.05
組別對照組5 μmol/L米諾環素組10 μmol/L米諾環素組F U87 p-AMPKα 1.00±0.03 2.48±0.55a 2.88±0.24a 24.608**AMPKα 1.00±0.07 0.95±0.08 1.00±0.12 0.368 SIRT1 1.00±0.05 3.73±0.25a 3.46±0.19a 201.476**mTOR 1.00±0.10 0.25±0.03a 0.03±0.02ab 189.305**Atg5 1.00±0.01 4.55±0.35a 5.88±0.19ab 361.924**組別對照組5 μmol/L米諾環素組10 μmol/L米諾環素組F LN229 p-AMPKα 1.00±0.06 5.99±0.20a 3.54±0.31a 392.806**AMPKα 1.00±0.08 0.99±0.04 0.87±0.04 2.687 SIRT1 1.00±0.01 3.24±0.32a 3.20±0.22a 96.815**mTOR 1.00±0.05 0.72±0.13a 0.12±0.04ab 91.879**Atg5 1.00±0.03 1.88±0.12a 2.70±0.34ab 49.022**
Tab.5 The expression levels of apoptotic proteins in U87 and LN229 cells after treatment with minocycline表5 不同濃度米諾環素處理后的U87和LN229細胞凋亡相關蛋白水平變化 ()
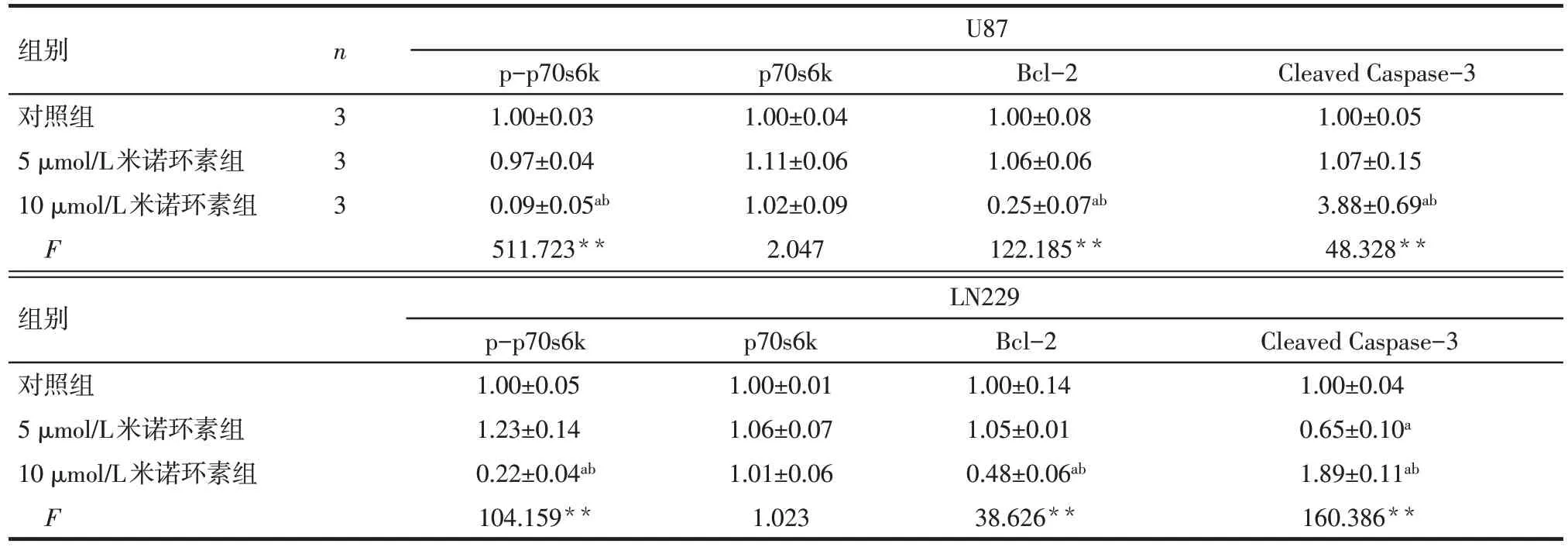
Tab.5 The expression levels of apoptotic proteins in U87 and LN229 cells after treatment with minocycline表5 不同濃度米諾環素處理后的U87和LN229細胞凋亡相關蛋白水平變化 ()
**P<0.01;a與對照組相比,b與5 μmol/L米諾環素組相比,P<0.05
組別對照組5 μmol/L米諾環素組10 μmol/L米諾環素組F n 333 U87 p-p70s6k 1.00±0.03 0.97±0.04 0.09±0.05ab 511.723**p70s6k 1.00±0.04 1.11±0.06 1.02±0.09 2.047 Bcl-2 1.00±0.08 1.06±0.06 0.25±0.07ab 122.185**Cleaved Caspase-3 1.00±0.05 1.07±0.15 3.88±0.69ab 48.328**組別對照組5 μmol/L米諾環素組10 μmol/L米諾環素組F LN229 p-p70s6k 1.00±0.05 1.23±0.14 0.22±0.04ab 104.159**p70s6k 1.00±0.01 1.06±0.07 1.01±0.06 1.023 Bcl-2 1.00±0.14 1.05±0.01 0.48±0.06ab 38.626**Cleaved Caspase-3 1.00±0.04 0.65±0.10a 1.89±0.11ab 160.386**

Fig.5 Western blot results of SIRT1 after shSIRT1-1 or shSIRT1-2 interfering with U87 and LN229 cells圖5 shSIRT1-1、shSIRT1-2干擾U87和LN229細胞后SIRT1的蛋白水平變化

Fig.6 Western blot results of autophagy-associated proteins in shSIRT1 transfected U87 and LN229 cells treated with different concentrations of minocycline圖6 shSIRT1轉染的U87和LN229細胞在不同濃度米諾環素處理后自噬相關蛋白水平變化
Tab.6 The expression levels of autophagy-associated proteins in shSIRT1 transfected U87 and LN229 cells treated with different concentrations of minocycline表6 shSIRT1轉染各組的U87和LN229細胞在不同濃度米諾環素處理后自噬相關蛋白水平變化(n=3,)

Tab.6 The expression levels of autophagy-associated proteins in shSIRT1 transfected U87 and LN229 cells treated with different concentrations of minocycline表6 shSIRT1轉染各組的U87和LN229細胞在不同濃度米諾環素處理后自噬相關蛋白水平變化(n=3,)
**P<0.01;a 與對照組相比,b 與 shSIRT1 組相比,c 與 shSIRT1+5μmol/L米諾環素組相比,P<0.05
組別對照組shSIRT1組shSIRT1+5 μmol/L米諾環素組shSIRT1+10 μmol/L米諾環素組F U87 SIRT1 1.00±0.06 0.60±0.02a 0.16±0.05ab mTOR 1.00±0.10 0.70±0.07a 1.80±0.25ab LC3B 1.00±0.05 0.02±0.01a 0.23±0.05ab 0.02±0.02abc 3.08±0.38abc 0.30±0.02ab 348.214**63.046**427.207**組別對照組shSIRT1組shSIRT1+5 μmol/L米諾環素組shSIRT1+10 μmol/L米諾環素組F LN229 SIRT1 1.00±0.07 0.35±0.06a 0.06±0.02ab mTOR 1.00±0.09 0.46±0.05a 1.77±0.13ab LC3B 1.00±0.02 0.06±0.02a 0.05±0.02a 0.66±0.12abc 85.861**1.82±0.06ab 164.777**1.85±0.10abc 801.987**
3 討論
膠質瘤是一種中樞神經系統的惡性腫瘤,具有高病死率、易復發的特點,單純手術、放化療治療后效果欠佳。盡管近年來部分研究者提出基因治療、免疫治療、分子靶向治療等方式,但實際效果并不顯著,因此探索新的治療策略仍有必要。米諾環素作為一種半合成的抗生素,因其抗菌譜廣且能跨越血腦屏障,曾被廣泛用于尋常型痤瘡和一些性傳播疾病治療;但隨著研究的深入,米諾環素的其他非抑菌作用也逐漸被發現[5,9-10]。近幾年的研究證實米諾環素能抑制多種腫瘤的生長,其通過抑制AKT/mTOR/p70S6k/4E-BP1 信號通路引起缺氧誘導因子(HIF)-1α下調并抑制卵巢癌的生長[11],還能通過下調MKK1/2-ERK1/2 介導Rad51 表達進而抑制非小細胞肺癌的生長[12]。而目前米諾環素對膠質瘤作用的研究相對較少,Liu等[7]的研究表明米諾環素通過誘導自噬抑制膠質瘤的生長。本課題組前期研究結果也顯示50~200 μmol/L 米諾環素均能抑制人類膠質瘤U87 和LN229 細胞的生長和增殖,并伴有兩種膠質瘤細胞的凋亡和自噬水平均明顯升高[8],因此推測其原因可能與細胞凋亡和自噬有關。
細胞自噬和凋亡均能引起細胞發生程序性死亡(programmed cell death,PCD)從而抑制腫瘤細胞的生長,這也是放化療方案治療膠質瘤的作用機制之一。細胞凋亡常被認為是Ⅰ型PCD,其包含有外源性死亡受體途徑和內源性線粒體兩大途徑,其中Caspase-3 是該過程中關鍵酶和主要效應因子。本研究發現10 μmol/L 米諾環素組中2 種膠質瘤細胞中活化形式的Caspase-3蛋白表達顯著升高,這表明細胞凋亡參與了抑制膠質瘤細胞生長的過程,但5 μmol/L 米諾環素組并未發生明顯的細胞凋亡,分析原因可能與SIRT1一定程度上抑制凋亡相關信號通路或其他機制有關。細胞自噬也稱為Ⅱ型PCD,適度的自噬通過降解被破壞的細胞器促進營養物質被細胞重新利用,進而保護細胞,但過度的自噬反而會導致細胞發生自噬性死亡。本研究發現5 μmol/L和10 μmol/L米諾環素組中LC3B和Atg5的表達水平均較對照組明顯升高,提示細胞自噬同樣參與了抑制膠質瘤細胞的生長。在此基礎上,本研究還進一步揭示了米諾環素誘導膠質瘤細胞自噬與AMPK/SIRT1 通路有關,而誘導膠質瘤細胞凋亡與p70s6k/Bcl-2通路有關。此外,本研究還發現在shSIRT1干擾降低SIRT1 表達水平后,米諾環素同時處理U87和LN229 細胞,兩種細胞中mTOR 蛋白水平明顯升高,但U87 細胞中LC3B 蛋白變化不顯著,而LN229細胞中LC3B 蛋白有增加趨勢。這預示米諾環素除通過SIRT1/mTOR 信號通路調控自噬之外,可能還存在其他途徑作用于自噬相關通路。筆者將在后續實驗對上述問題進行進一步探討。
