生境梯度影響下的天然紅松種群空間格局與種內關聯
2020-09-17 00:01:14孫志虎谷會巖陳祥偉
生態學報 2020年15期
董 雪,杜 昕,孫志虎,谷會巖,陳祥偉
東北林業大學林學院, 哈爾濱 150040
植物種、植物群落及植被景觀有其自身的空間分布規律,稱為廣義空間分布格局;植物種群在群落中的非隨機分布稱為種群的分布格局,即狹義格局,其是植物群落斑塊與植物景觀斑塊形成的基礎[1-2]。種群格局分析的實質是研究單個或多個種群在群落內部的鑲嵌結構,進而揭示空間格局的形成原因。分析種群分布格局有助于認識種群的生物學特性、生態學過程及物種與環境因子間的關聯性。種群的空間格局對研究尺度具有很強的依賴性[3-4]。而相對于傳統格局分析方法,空間點格局分析能夠量化植物種群在不同尺度上的空間分布格局與種內、種間聯結關系;其以個體空間坐標為基礎,在分析過程中最大限度的利用二維空間點信息,具有較強的檢驗效力[5-9]。在點格局分析基礎上演伸出的標記點格局[10-11]及重復采樣點格局分析[12-15]等方法,可以更詳盡的探索不同空間尺度上的生態學問題。種群空間格局的形成受到物種自身生物學特性與群落環境的共同影響。不同生境中相同物種的空間分布特征存在差異性[16-17],而明確尺度的點格局分析可以清晰的揭示不同生境中的種群分布狀態與生態學過程[9,18-19]。
紅松是第四紀冰川孑遺種,以其為建群種的闊葉紅松林長期存在于東北亞地區,是該區的山地地帶性頂級植被,具有復雜的動植物成分[20]與系統穩定性。建群種紅松(Pinuskoraiensis)具有較寬的超體積生態位,在不同的環境下可與不同樹種混交,構成不同林型原始紅松林。曾有學者探討過地形因子對原始紅松林型的影響,認為闊葉紅松林林型的變化主要是由坡位、坡度等地形因子變化,進而造成土壤水分條件變動引起的。在同一分水嶺內隨著坡位下降,坡度不斷減小,土壤水分形成干燥、潮濕、濕潤、重濕潤的環境梯度,并產生與之對應的紅松林林型。基于此,許多學者對闊葉紅松林林型組與林型進行了劃分,如徐化成[21]將典型闊葉紅松林劃分為3個林型組,6個林型,其中高海拔林型組包括高海拔蘚類云冷杉紅松林;坡地林型組包括細葉苔草蒙古櫟紅松林、毛榛紫椴紅松林、灌木楓樺紅松林;谷地林型組包括蕨類云冷杉紅松林和春榆水曲柳紅松林。李文華[22]將闊葉紅松林劃分為谷地潮濕紅松林型組、緩坡潮濕紅松林型組、斜坡半濕潤紅松林型組及陡坡半干旱紅松林型組。
近年,已有較多研究者應用空間點格局分析的方法對闊葉紅松林中立木空間分布規律進行了探索,如劉妍妍[23-24]與侯紅亞和王立海[25]分別基于大樣地調查數據分析了典型紅松林中主要喬木樹種空間分布;夏富才[26]研究了南部闊葉紅松林與不同演替階段次生林中林木空間分布格局等。目前,研究多以闊葉紅松林或特定林型中各喬木樹種為研究對象,探討樹種的生物學特性與生長過程。然而需要指出的是,不同生境中的相同植物種群在其定植、生長的過程中所經歷的生態學過程必然有所差異,生境梯度下闊葉紅松林中各樹種的空間分布規律有待進一步探索。通過分析處于不同生境的原始紅松林中喬木空間分布格局的差異,加深對影響林木定植、生長的生態學過程的理解,將有利于闊葉紅松林的經營管理與維持紅松林生態系統的長期穩定。基于此,本研究參考李文華對不同原始紅松林型組的界定,選取7塊分別位于4種不同坡位的原始紅松林小樣地,以建群種紅松為研究對象,應用重復采樣點格局分析的方法研究生境梯度作用下不同林層紅松的空間分布格局,并回答以下科學問題:①隨著坡位升高、坡度增大、典型紅松林各林層中紅松種群空間分布格局有何變化規律?②生境梯度影響下典型紅松林中紅松種群種內聯結規律有何變化趨勢?
1 研究區概況及研究方法
1.1 研究區概況
研究區位于黑龍江省涼水國家級自然保護區(47°10′55″N,128°53′20″E),屬小興安嶺南坡。研究區氣候類型為溫帶大陸季風氣候,年平均氣溫-0.3℃,年平均最高氣溫7.5℃,年平均最低氣溫-6.6℃,年平均降水量676 mm,全年降雨多集中于6—8月份,有效積溫2200—2600℃,無霜期100—120 d。原生植被類型是以典型闊葉紅松林為代表的溫性針闊混交林。目前,在逆行演替和進展演替共同作用,形成了以不同演替階段的次生林為基底,以人工林及原始林為斑塊的鑲嵌分布的森林景觀。地帶性土壤為暗棕壤,非地帶性土壤有草甸土、澤土、泥炭土、棕色森林土、白漿土等。
1.2 樣地布設與調查
在涼水自然保護區19林班內,參照李文華[22]對紅松林型生態系列的界定,綜合林木組成、林下植被、坡位、坡度以及土壤水分等信息,在由坡底到坡頂的生境梯度上平行等高線設置7塊50 m×50 m的樣地[27],各樣地基本信息見表1。樣地調查采用相鄰格子法,將調查樣地劃分為100個5 m×5 m的調查單元,調查并記錄全部株高大于10 cm的喬木樹種在樣地中的相對位置坐標與株高。綜合考慮徐化成[21]、李景文[28]、安慧君和張韜[29]對原始紅松林林層的劃分,并參考行業標準(LY:1572-2000)[30]中更新層的劃定,將原始紅松林林層劃分為主林層(H≥16 m)、亞林層(10 m≤H<16 m)、演替層(1.5 m≤H<10 m)與更新層(0.1 m≤H<1.5 m)。依據調查喬木株高,判定喬木所處林層,以待進一步分析。各樣地中,土壤取樣按五點法進行,去除土壤表面雜草、枯落物與腐殖質后,挖開表土,在5—10 cm土層取土樣,用于土壤含水率分析。
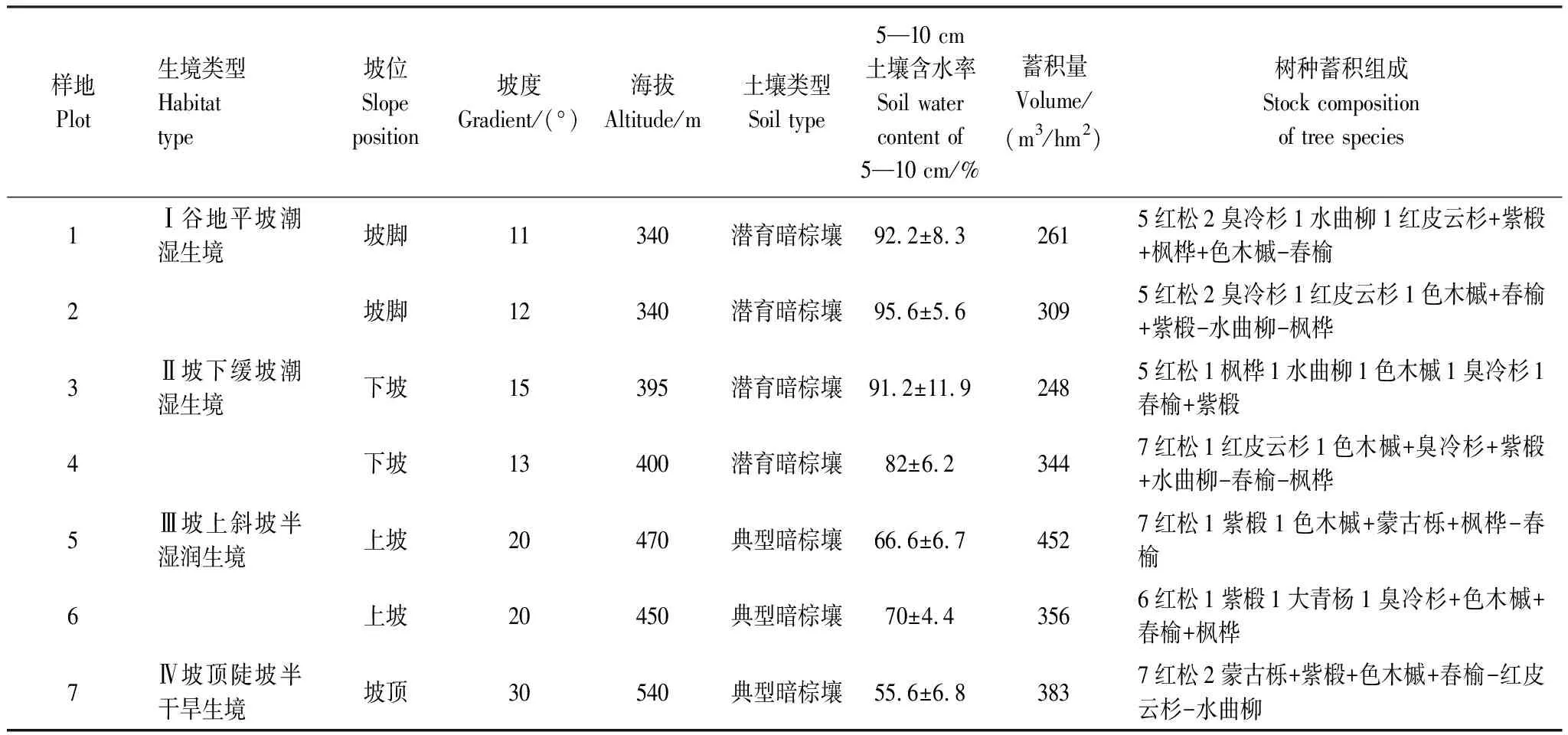
表1 紅松原始林各樣地基本信息表
1.3 點格局分析與重復采樣點格局分析
點格局分析(Point pattern analysis)是以植物個體空間坐標為基礎,分析不同尺度下種群格局與種間關系的分析方法[5-9]。而重復采樣點格局分析(Replicate pattern analysis),是基于多個小樣本空間的空間分析方法[12-15]。相對于依賴單一大樣本的點格局分析,重復采樣點格局分析可以增強試驗結果的普適性、提高結果的可靠程度。
本研究采用O-ring(以下簡稱Or)函數為基本函數,進行點格局分析與重復采樣點格局分析[31]。Or函數基于Ripely′s K函數與Mark相關函數,通過計算點外特定距離上的發生頻率確定空間格局[3]。相對于Ripely′s K函數與Ripely′s L函數,以特定寬度環為統計基礎的Or函數可以消除小尺度上的尺度累積效應[32-33]。
1.4 生境間種群空間分布規律比較
參考Pamela等[18]與Wang等[19]的方法,進行不同生境中紅松種群空間分布規律的比較。該方法通過Or函數計算值、蒙特卡洛(Monte Carlo)法構建的包絡線(envelop)及完全空間隨機(Complete spatial random,CSR)假設下Or函數的理論值(Theoretical value,Tv)間關系計算不同生境中種群空間格局的無偏估計y,對比不同生境的種群空間分布格局。
如果Or>99%置信區間上限(CL+),則y=(Or-CL+)/|CL+-Tv|>0
如果Or<99%置信區間上限(CL+),且Or>99%置信區間下限(CL-),則y=0
如果Or<99%置信區間下限(CL-),則y=(Or-CL-)/|CL--Tv|-<0
對單一種群(林層)進行重復采樣點格局分析,完全空間隨機假設的理論值Tv為:
研究兩種群(林層)間空間關系時,完全空間隨機假設的理論值Tv為:
式中,j為重復采樣次數;Areaj為第j次采樣的樣地面積;nj為第j次采樣時種群一中個體數量;mj為第j次采樣時種群二中個體數量。
研究中Or函數環寬設置為5 m,Monte Carlo法獲取99%置信區間。采用Programita 2014、Excel 2013、SigmaPlot 12.5進行數據分析及繪圖。
2 研究結果
2.1 生境梯度下紅松種群數量特征
從圖1可以看出,四類生境中,紅松種群結構差異較大。在由坡底到坡頂的生境梯度上紅松種群密度分別為340、412、160、308 株/hm2。不同林層中紅松種群密度亦有較大差異,生境Ⅰ與生境Ⅱ中,紅松更新分別達到了226 株/hm2與208 株/hm2,生境Ⅲ與生境Ⅳ中紅松更新密度僅為20 株/hm2與60 株/hm2。生境Ⅰ、Ⅱ、Ⅲ、Ⅳ演替層中紅松密度分別為56、118、18、72 株/hm2。而在生境梯度上紅松原始林亞林層與主林層中紅松種群密度有增大趨勢,各生境亞林層與主林層紅松種群密度分別為8、18、18、52 株/hm2及50、68、104、124 株/hm2。總體而言,在由坡底到坡頂的生境梯度上,紅松種群結構有從倒J型向J型轉換的趨勢。
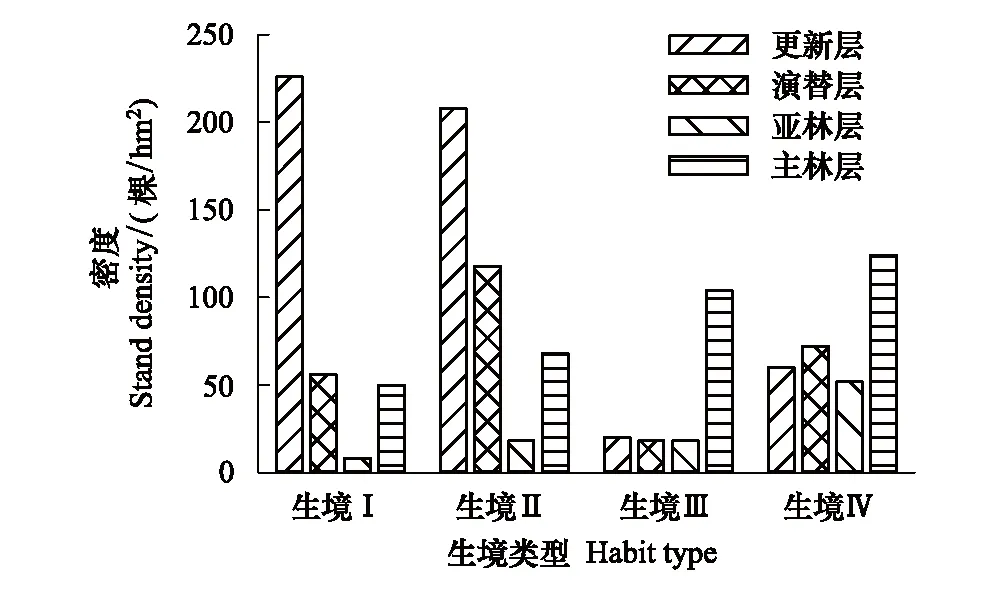
圖1 不同生境中原始紅松林紅松種群結構
2.2 四類生境中各林層紅松種群重復采樣點格局
將隸屬于不同生境的紅松林樣地作為重復采樣,對不同林層的紅松立木進行空間點格局分析,并根據零模型與包絡線的關系簡要換算,用于比較不同生境間紅松種群空間分布規律的差異(其中2號樣地亞林層中紅松不足兩株,生境Ⅰ亞林層規律為1號樣地亞林層規律)。結果如圖2所示:不同生境原始紅松林中紅松種群在由更新層向主林層生長的過程中,種群空間格局均由聚集分布向隨機分布轉變;各生境原始紅松林主林層與亞林層中紅松種群基本均服從隨機分布,四類生境原始紅松林內紅松種群空間分布規律的差異主要體現在更新層及演替層的較小研究尺度上。各生境中,更新層內紅松種群聚集強度及聚集規模排序為生境Ⅰ>生境Ⅱ>生境Ⅳ>生境Ⅲ。生境Ⅰ與生境Ⅱ演替層中紅松種群在較小研究尺度上體現為聚集分布,而生境Ⅲ與生境Ⅳ中紅松種群則表現為空間隨機。總體而言,處于相對濕潤生境中的原始紅松林更新層與演替層中紅松種群聚集規模與聚集強度大于處于相對較干旱生境中的原始紅松林。
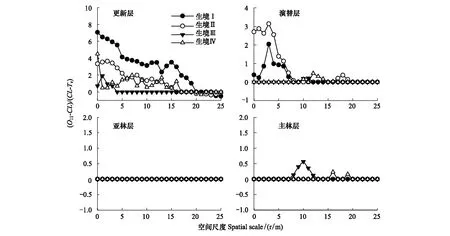
圖2 不同生境中原始紅松林中各林層中紅松種群分布規律比較
2.3 不同生境梯度下紅松種群種內聯結差異
將分布于不同生境中的原始紅松林樣地分別作為重復采樣,合并分析生境梯度影響下紅松種群種內聯結關系。基于所得結果中零模型與包絡線間的關系,比較不同生境中原始紅松林的紅松種內聯結關系。結果如圖3所示:各生境中,亞林層中紅松種群在全部研究尺度上均未與主林層表現出空間關聯性;更新層、演替層與主林層的紅松種群間均表現出一定的空間關聯性,相對于更新層與主林層,演替層與主林層中紅松種群空間關聯性更強。生境Ⅰ原始紅松林中更新層中紅松種群在1—2 m研究尺度下與主林層表現出微弱的空間負關聯;生境Ⅱ更新層與主林層中紅松種群在1—5、7、9—12 m的尺度上均表現出弱空間負關聯;生境Ⅲ更新層中紅松種群與主林層在全部研究尺度上均未表現出空間相關性;生境Ⅳ中,紅松林更新層與主林層在7—11 m尺度上表現出空間微弱正關聯,其余尺度未表現出空間相關性。生境Ⅰ及生境Ⅱ中原始紅松林演替層與主林層在1—5 m研究尺度上表現出空間負關聯,而生境Ⅲ與生境Ⅳ中原始紅松林則具有相反規律,在較小研究尺度上與主林層表現為空間正關聯。
3 討論
3.1 生境對不同林層紅松種群空間分布格局的影響
研究顯示,各生境中原始紅松林更新層與演替層中紅松種群表現出一定的空間聚集性,其聚集強度隨著研究尺度增大逐漸減弱,種群空間分布格局逐漸由聚集分布像隨機分布轉變;亞林層與主林層中紅松種群的分布格局趨近于隨機分布,規律與前人相同[23-25]。各生境原始紅松林演替層中紅松種群聚集規模與聚集強度相較于更新層中紅松種群均有所下降,體現出紅松更新在生長過程中的環境過濾與種內、種間競爭。
各生境原始紅松林更新層中紅松種群聚集強度與規模有隨著坡位增高而減弱的趨勢,更新層中紅松種群的聚集強度與林下更新數量相關,即更新數量越多,聚集強度越大。各生境更新層中,0—1 m尺度上紅松種群聚集強度為生境Ⅰ>生境Ⅳ>生境Ⅱ>生境Ⅲ,而根據調查數據(未發表數據),在0—1 m尺度上更新層中紅松種群聚集強度較高的生境Ⅰ與生境Ⅳ中,紅松苗高均較低。生境Ⅱ演替層中紅松種群聚集強度較大,其原因是該生境調查樣地演替層中紅松立木多且樹高普遍較低(1.5—5 m),這也可側印以上論點。
相對于生境Ⅲ,生境Ⅳ中林下植被稀疏。根據徐化成的劃分,生境Ⅳ中分布的紅松林型為典型的陡坡細葉苔草蒙古櫟紅松林[21],樣地內坡度較大處灌木層僅有少量衛矛(Euonymusverrucosus)分布,為紅松更新空出了生態位。這或也是相對生境Ⅲ,生境中Ⅳ具有更多紅松更新,進而具有更高聚集規模與強度的原因。
總體而言,四類生境中,紅松種群從更新層向主林層的發育過程即是一種種群空間格局由聚集分布向隨機分布轉變的過程;相較于生境Ⅲ與生境Ⅳ中紅松種群,處于相對濕潤的生境Ⅰ與生境Ⅱ中更新層與演替層中紅松種群在較小研究尺度上具有更大的聚集強度與聚集規模,這應與林下幼樹、幼苗數量有關;而相對于生境Ⅲ,生境Ⅳ更新層與演替層中聚集規模與聚集強度均更大,這是由于在生境Ⅲ中林下植被發育較為強烈,侵占了紅松更新的最適生態位,而生境Ⅳ中林下植被不發達,為紅松更新預留了生態位。
3.2 生境梯度下紅松種群種內聯結差異性
在自坡底到坡頂的生境梯度上,不同的原始紅松林型中,亞林層與主林層中的紅松種群間空間關系均表現為空間獨立、演替層—主林層、更新層—主林層中的紅松種群則表現出一定的空間關聯性。演替層與主林層間的空間關聯強度較之更新層更強,分析其原因應與紅松幼苗生長過程中受到的生境過濾作用有關,這使紅松幼苗從更新層生長到演替層的過程中逐漸體現出了物種適宜的生態位。
在自坡底到坡頂的生境梯度上,演替層與主林層的空間關聯性有從空間負關聯向空間正關聯轉變的趨勢,造成這一現象的可能原因有,生境梯度上紅松在主林層中的株數占比不斷提高,因此紅松幼樹有更大機會分布在主林層中紅松立木周圍。這一現象的產生也與不同生境中灌木、草本層的空間分布有關,如生境Ⅲ中林窗中狗棗獼猴桃(Actinidiakolomikta)發育旺盛[13],闊葉樹林冠下生境中則常分布有較為茂密的毛榛(Corylusmandshurica)灌叢,這些林下植被的分布,侵占了紅松更新及幼樹的生態位,使紅松幼樹、幼苗分布在林下植被較為稀疏的紅松林冠下。不同生境中,林下優勢灌、草有所不同,在坡位較低、坡度較小的林型內,東北珍珠梅(Sorbariasorbifolia)、東北山梅花(Philadelphusschrenkii)優勢度較大,而在坡位較高的生境中,毛榛、刺五加(Acanthopanaxsenticosus)等優勢度較大,而林下植被變化均會影響幼樹、幼苗的空間分布規律。此外,坡位較高的闊葉紅松林中光資源可獲取性較強,更新層中紅松種群間生長到演替層的過程中受到光資源的制約較小,紅松樹冠下光環境可以滿足幼苗、幼樹生長的需求;而在坡底的生境中,紅松幼樹、幼苗更傾向分布于闊葉樹樹冠下[34],這與不同林冠下紅松幼樹、幼苗的保存率有關,中國科學院林業土壤所得材料表面,紅松林冠下15年以下的更新不少于闊葉樹林冠下,而15年以上的幼苗則以闊葉樹下的為高。同時,由于光資源的制約,坡位較低生境中紅松樹冠下的幼苗生長更慢[35],因此下坡位生境中演替層與主林層中的紅松種群在較小研究尺度上表現出空間負關聯。植物的建植與生長受到生境過濾的影響,而生境過濾又涵蓋環境過濾與生物互做兩層意義,前者指非生物環境對植物的篩選作用,后者指植物間的正、負關聯[36-38],在探討生境梯度上植物空間分布規律時,二者都應加以考慮。
4 結論
(1)小興安嶺不同生境的原始紅松林中,主林層與亞林層中的紅松種群均表現為隨機分布;而在自坡底到坡頂的生境梯度上,更新層與演替層中紅松種群的聚集規模與聚集強度均有減弱趨勢,其原因與在自坡底到坡頂的生境梯度上,紅松幼樹、幼苗減少有關。
(2)小興安嶺不同類型原始紅松林內,亞林層與主林層中的紅松種群均表現為空間獨立;而在較小研究尺度上,幼苗、幼樹與主林層中紅松種群間的空間關系則隨著坡位上升、坡度增大由空間負關聯向空間正關聯轉變,這是生物與非生物因子共同作用的結果。相較于更新層,演替層與主林層中紅松種群的空間正、負關聯的程度均更強,這是生境過濾的結果。
