不同綿羊品種FSHR基因多態性及其與產羔數的關聯分析
2020-08-27 03:55:18劉玲玲馬海玉劉武軍
西北農業學報 2020年8期
劉玲玲,馬海玉,劉武軍
(新疆農業大學 動物科學學院,烏魯木齊 830052)
中國綿羊資源豐富,大多數地方綿羊品種為單羔品種,只有少數綿羊為多羔品種[1]。產羔數是家畜關鍵和復雜的經濟性狀,也是畜牧業可持續發展的重要因素[2]。產羔數為數量性狀,受多基因的控制,但產羔性狀的遺傳力較低,利用傳統選育方法獲得的遺傳進展較慢,標記輔助選擇能夠解決效率低下等問題,而尋找可靠的遺傳標記則成為關鍵。
目前已經確定能夠影響綿山羊產羔數的基因為BMPR1B(Bone morphogenetic protein receptor1B)基因[3-4],GDF9(Growth differentiation factor9)基因[5],SLC5A1(Solute carrier family5member1)基因[6],MTNR1A(Melatonin receptor1A)基因[7]。而排卵率和胚胎存活率也直接影響羊的產羔數[8],研究報道,KIT(Tyrosine-protein kinase kit)和NGF(Nerve growth factor)基因能夠影響卵泡的生長[9-10]。NCOA1(Nuclear receptor coactivator1)基因對胚胎的存活至關重要[11],Xu等[12]利用全基因組關聯分析研究高產綿羊與低產綿羊之間的差異,篩選出NCOA3(Nuclear receptor coactivator3)基因與Lcelandic綿羊產羔數有關。
KASP(Kompetitive allele specific polymerase chain reaction)技術是目前基因分型的一種重要手段。KASP主要是在廣泛的基因組DNA樣品中,對SNPs 和特定位點的InDels進行精準的雙等位基因判斷,從而進行基因分型。主要核心是利用2個通用熒光探針、2個通用淬滅探針,以及多個位點特異引物對SNP位點進行檢測。相比于其他的檢測SNP方法,KASP技術擁有更高的通量、更快的速度、更低的成本及更準確的結果[13-14]。Zhang等[15]利用KASP技術對造成荷斯坦牛遺傳缺陷的5個SNP及3個插入和缺失進行分析。Kusza等[16]利用KASP技術對影響山羊產奶性狀的48個SNP進行分析。
筆者前期對新疆不同產羔數綿羊的LAF-Seq測序數據進行GWAS分析,篩選獲得可能與綿羊產羔數有關的關鍵基因之一FSHR[17]。促卵泡激素受體(Follicle-stimulating hormone receptor,FSHR)是GPCR(G protein-coupled receptors)家族的成員,這類受體具有3個受體蛋白鏈:胞外域、胞內域和跨膜域[18-19]。FSHR轉導FSH信號的機制是:FSHR的胞外α-螺旋環和胞外域在細胞膜的表面偶聯FSH,FSHR被激活后通過胞內域和跨膜域與G蛋白偶聯,最后完成信號轉導[20]。研究發現在豬FSHR基因的第10外顯子上,rs322800083和rs332115220構成的單倍型C-T可以作為香豬產仔數性狀輔助選育的分子標記[21];黔北麻羊FSHR基因的第1和第10外顯子突變對多羔性狀具有重要作用[22]。目前未見FSHR基因與阿勒泰羊、吐魯番黑羊、和田紅羊產羔數之間關系的報道。
本試驗以阿勒泰羊、吐魯番黑羊、和田紅羊為研究對象,基于NCBI中已有的FSHR基因的錯義突變位點(g.75132817C>T,g.75320579G>A,g.75320741G>A),利用KASP技術對突變位點進行分型,探究不同品種綿羊FSHR基因SNP位點的多態性,并與產羔數進行關聯分析,篩選影響不同品種高繁殖力的關鍵位點,為不同綿羊品種產羔數的選育提供分子標記。
1 材料與方法
1.1 試驗動物
65只阿勒泰羊來自福海種羊場,240 只吐魯番黑羊來自新疆吐魯番市托克遜縣,111 只和田紅羊來自新疆和田地區皮山縣,均為經產母羊 (2~3胎次)。
1.2 基因組DNA提取
綿羊血液樣本使用血液/組織DNA磁珠法提取試劑盒(志昂生物科技有限公司)對DNA進行提取,DNA質量利用 15 g/L瓊脂糖凝膠電泳檢測。
1.3 主要儀器設備
磁力架(Life Technologies DynaMagTM-2),離心機(Eppendorf Centrifuge 5424),Douglas Scientific NEXAR,Douglas Scientific Soellex,Douglas Scientific ARRAY,試劑盒(志昂生物科技有限公司)。
1.4 引物設計與合成
利用Prime 3軟件設計引物,由百邁客生物科技有限公司負責完成引物設計及合成。引物具體信息見表1。

表1 引物信息Table 1 Primer information of genes
1.5 PCR反應程序
PCR反應體系:2.5 μL的2×KASP Master Mix,0.07 μL引物,30~50 μg的DNA,總反應體積為5 μL(ddH2O補足)。為避免對試驗結果的錯誤判斷,因此設置空白對照(NTC),其DNA模板用ddH2O代替。PCR反應程序:95 ℃預變性10 min; 95 ℃變性20 s,10個循環,61~55 ℃復性及延伸60 s(-0.6 ℃/循環),10個循環;95 ℃變性20 s,26個循環;最后55 ℃復性及延伸 60 s,26個循環。反應結束后利用帶FRET功能的酶標儀進行熒光數據讀取(數據讀取在40 ℃以下)。利用LGC_OMEGA軟件生成基因的分 型圖。
1.6 數據統計方法
利用KASP分型技術對位點分型成功后,計算每個位點的等位基因頻率和基因型頻率,并進行哈代-溫伯格平衡檢驗。利用SPSS 19.0中單因素方差分析和t檢驗對平均產羔數和突變位點基因型進行關聯分析,產羔數以“平均數±標準差”表示。
2 結果與分析
2.1 3個位點在3 個群體中的KASP分型結果
KASP分型技術是一種靈活的方法,其基因分型錯誤率為0.7%~1.6%[13],利用此方法進行FSHR基因的3個位點在阿勒泰羊、吐魯番黑羊、和田紅羊群體中的分型結果可靠。阿勒泰羊群體中,g.75132817C>T位點存在3種基因型:CC、CT、TT;g.75320579G>A位點存在2 種基因型:GG、GA、g.75320741G>A位點存在3種基因型:GG、GA、AA(圖1)。吐魯番黑羊群體中,g.75132817C>T位點存在3種基因型:CC、CT、TT;g.75320579G>A位點存在3種基因型:GG、GA、AA;g.75320741G>A位點存在3種基因型:GG、GA、AA(圖2)。和田紅羊群體中,g.75132817C>T位點存在3種基因型:CC、CT、TT;g.75320579G>A位點存在2種基因型:GG、GA;g.75320741G>A位點存在3種基因型:GG、GA、AA(圖3)。
2.2 3個群體FSHR基因不同位點的基因多態性分析
吐魯番黑羊群體中,3個位點均為野生型基因型頻率高于雜合型和突變型的基因型頻率,說明突變的個體數較少,突變頻率比較低;阿勒泰羊與和田紅羊群體中,g.75132817C>T、g.75320741G>A 2個位點的野生型的基因型頻率高于雜合型和突變型的基因型頻率,而g.75320579G>A位點的野生型基因型頻率低于雜合型的基因型頻率,說明g.75320579G>A位點在這2個群體中的突變發生率較高。3個位點在3個群體中均處于哈代-溫伯格平衡狀態(P>0.05)。在3個綿羊群體中,FSHR基因g.75132817位點的C為優勢等位基因;g.75320579位點的G為優勢等位基因;g.75320741位點的G為優勢等位基因(表2)。FSHR基因g.75132817C>T位點在3個群體中均表現為中度多態(0.25
2.3 FSHR基因的3個位點與不同品種羊產羔數的關系
g.75132817C>T位點多態性與和田紅羊產羔數有顯著關聯性(P<0.05),突變型TT個體產羔數顯著高于野生型CC個體。此位點的突變降低阿勒泰羊的產羔數,增加吐魯番黑羊與和田紅羊的產羔數;g.75320579G>A位點多態性與阿勒泰羊產羔數有顯著關聯性(P<0.05),野生型GG個體產羔數顯著高于雜合型GA個體。此位點的突變降低阿勒泰羊的產羔數,增加和田紅羊的產羔數;g.75320741G>A位點多態性與阿勒泰羊和吐魯番黑羊的產羔數均有關聯性,在阿勒泰羊群體中,野生型GG個體產羔數極顯著高于雜合型GA個體和突變型AA個體(P< 0.01),突變導致阿勒泰羊產羔數降低;吐魯番黑羊群體中,突變型AA個體產羔數顯著高于野生型GG個體(P<0.05),突變導致吐魯番黑羊產羔數增加;和田紅羊群體中,不同基因型產羔數的差異不顯著(表5)。

A:g.75132817C>T;B:g.75320579G>A;C:g.75320741G>A;下同 The same below
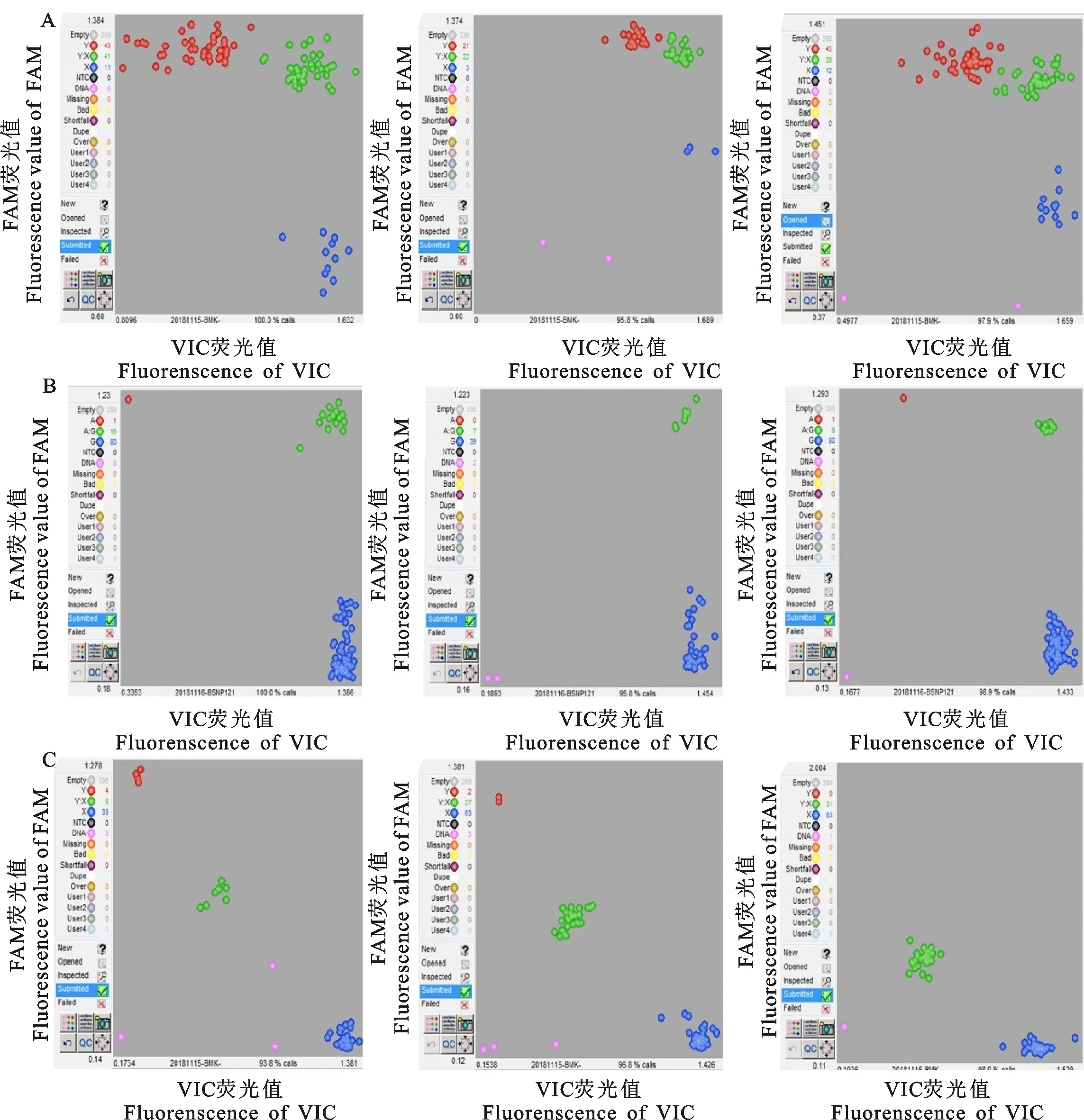
圖2 吐魯番黑羊FSHR基因3個位點的分型圖Fig.2 KASP results of three loci of FSHR gene in Turpan black sheep
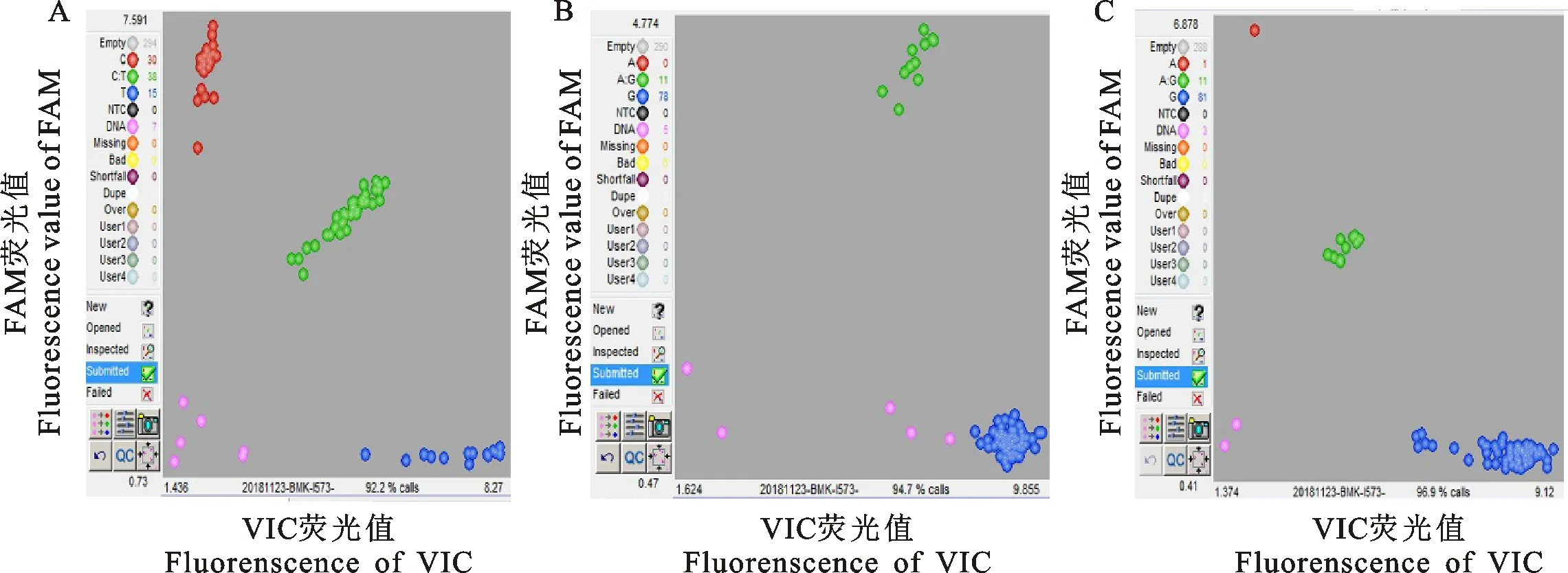
圖3 和田紅羊FSHR基因3個位點的分型圖Fig.3 KASP results of three loci of FSHR gene in Hetian sheep

表2 綿羊FSHR基因不同位點的基因型頻率和基因頻率Table 2 Frequencies of genotype and gene in different loci of FSHR genes in sheep

表3 FSHR基因3個位點在不同綿羊群體中的群體遺傳學分析Table 3 Population genetics analysis of three loci of FSHR genes in different sheep populations
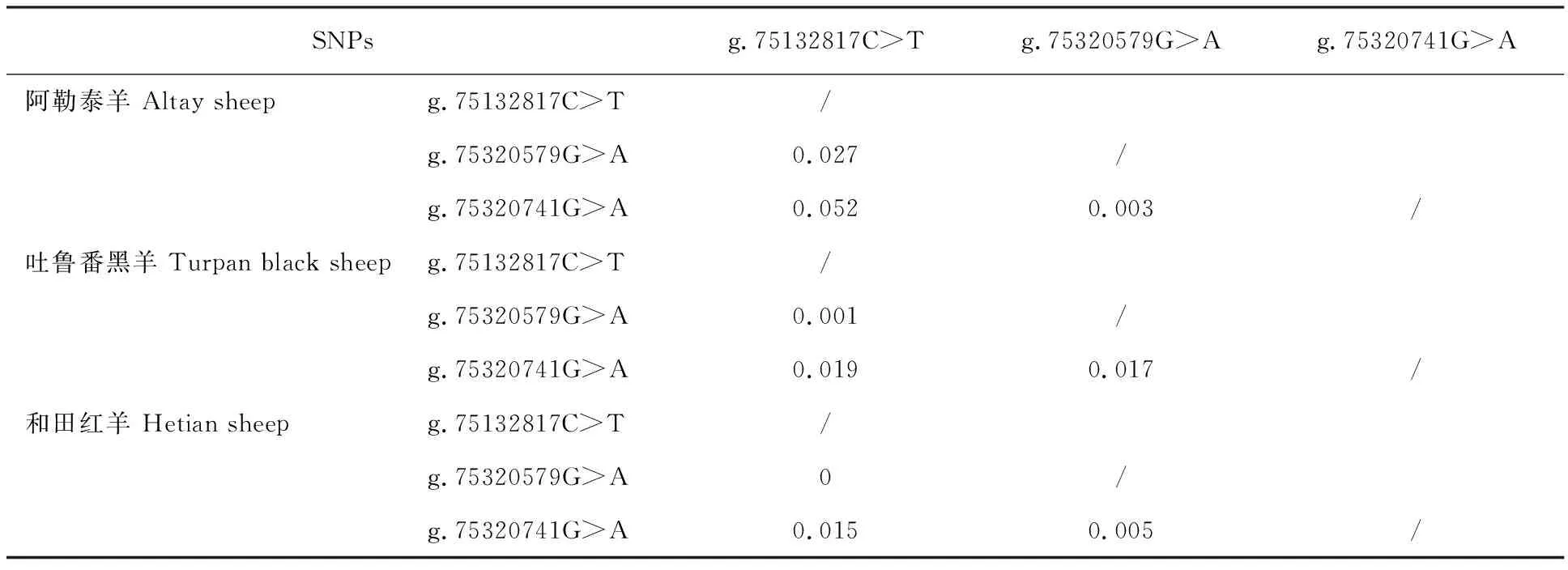
表4 不同綿羊群體FSHR基因SNPs 位點連鎖不平衡分析Table 4 Analysis of linkage disequilibrium of SNPs of FSHR gene in different sheep populations
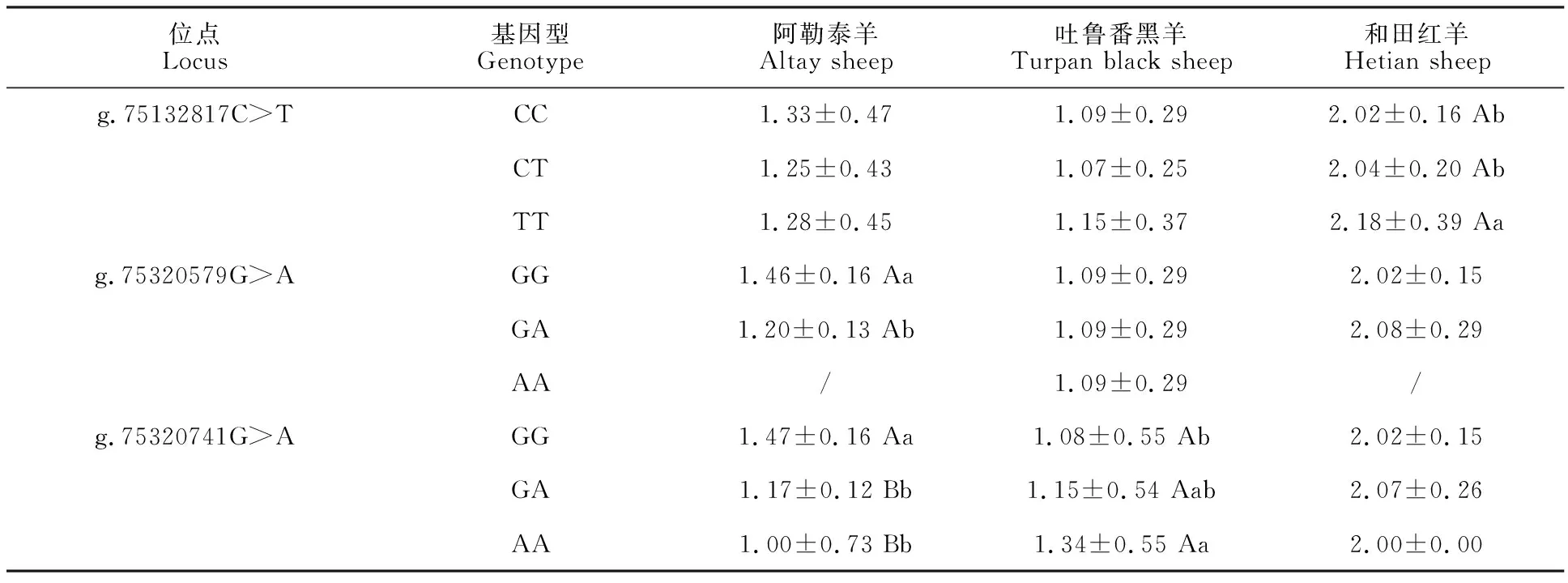
表5 FSHR基因SNPs位點各基因型與綿羊產羔數關聯分析Table 5 Association analysis of each genotype of FSHR gene SNPs locus with litter size
3 討 論
3.1 FSHR基因多態性與繁殖性能的關系
對哺乳動物FSHR基因多態性與繁殖性狀之間的關系研究較多。研究表明,FSHR基因在動物的繁殖中起關鍵作用[23-24]。吳井生等[25]對梅山豬的產羔數研究發現,FSHR基因第10外顯子存在 2 個多態位點,這 2 個位點對小梅山豬產仔數有顯著影響。Zhang等[26]發現FSHR基因第10外顯子有3個SNP位點(C1 491T、G1 885A、C1 977T),結果表明這些SNP可作為豬品種選育的分子標記。Chu等[27]在湖羊FSHR基因的5′側翼區域發現 2 個突變(g.681T> C和g.629C> T)和小尾寒羊中3個新突變(g.200G> A,g.197G> A和g.98T> C)這5個突變位點均與產羔數有關。Pan等[28]在小尾寒羊和湖羊中發現FSHR基因的5′側翼區 1 個新的突變位點(g.47C> T),此位點與產羔數相關。Wang等[29]同樣發現FSHR基因的位點(g.47C>T)與小尾寒羊和湖羊的產羔數有關。FSHR基因在山羊中也有研究,潭廣梅等[30]對海門山羊FSHR基因進行分析,發現野生型比突變純合型基因型多0.34只,等位基因與海門山羊產羔數呈顯著正相關。馬向明等[31]研究發現,在波爾山羊FSHR基因5′上游處檢測1個SNP(G-739A),該突變與平均排卵數(P<0.01)及平均胚胎可利用率顯著相關。FSHR基因在豬、羊中均有研究,研究最多的為FSHR基因的第10外顯子。在陳祥等[22]的研究中,FSHR基因第1外顯子的突變在黔北麻羊和貴州黑山羊群體中存在3種基因型,與本研究中FSHR基因第1外顯子 (g.75132817)的結果一致。
3.2 FSHR基因與阿勒泰羊、吐魯番黑羊、和田紅羊產羔數的關系
本研究中,g.75132817位點G突變為A,導致丙氨酸(Ala)變為蘇氨酸(Thr);g.75320579位點G突變為A,導致精氨酸(Arg)變為組氨酸(His);g.75320741位點C突變為T,導致蘇氨酸(Thr)變為異亮氨酸(Ile)。群體遺傳學分析結果表明,FSHR基因g.75132817C>T位點在阿勒泰羊、吐魯番黑羊、和田紅羊群體中為中度多態,說明這個位點的選擇潛力比較大,而g.75320579G>A和g.75320741G>A 2 個位點在3個綿羊群體中為低度多態,說明這 2 個位點的選擇潛力較小。卡方檢驗結果表明,3個位點在3個綿羊群體中均處于哈代-溫伯格平衡狀態 (P>0.05),表明這3個位點在長期的選擇過程中已趨于穩定。關聯分析結果表明,同一位點在不同綿羊群體中受到的選擇壓力不一致,g.75132817C>T位點在和田紅羊群體中,突變型TT個體產羔數顯著高于野生型CC個體,在吐魯番黑羊群體中,TT個體產羔數高于CC個體,但差異不顯著,在阿勒泰羊群體中,TT個體產羔數低于CC個體,說明此位點的突變一定程度上增強和田紅羊與吐魯番黑羊的產羔能力;g.75320579G>A位點在阿勒泰羊中,野生型GG個體產羔數高于于GA個體,說明此位點的突變一定程度上降低阿勒泰羊產羔能力;在和田紅羊中,GA個體產羔數高于GG個體的產羔數,說明此位點的突變增強和田紅羊的產羔能力;g.75320741G>A位點在和田紅羊和阿勒泰羊群體中突變型AA個體產羔數低于野生型GG個體,在吐魯番黑羊中,AA個體產羔數高于GG個體,說明此突變在一定程度上降低和田紅羊和阿勒泰羊的產羔能力,增強吐魯番黑羊的產羔能力。
不同位點基因型與不同綿羊品種產羔數的關聯結果不同,在陳祥等[22]的研究中,黔北麻羊和貴州黑山羊群體中,FSHR基因(第1外顯子)突變的不同基因型個體之間產羔數差異不顯著,與本研究結果不同。可能是由于地域不同造成的自然選擇不同或者是品種不同造成的,綿羊FSHR基因第1外顯子和第9外顯子的突變與產羔數的關系具有種屬差異性。
4 結 論
研究表明FSHR基因g.75132817C>T、g.75320579G>A和g.75320741G>A 3個位點的多態性與阿勒泰羊、吐魯番黑羊、和田紅羊產羔數有關聯性,3個位點的突變均導致阿勒泰羊產羔數降低;g.75132817C>T位點突變導致和田紅羊和吐魯番黑羊的產羔數升高,并且顯著提高和田紅羊的產羔數,說明此位點可作為和田紅羊選育的輔助標記;g.75320741G>A位點的突變導致吐魯番黑羊產羔數升高,g.75320741G>A位點可作為吐魯番黑羊選育的輔助標記。
