NGF過表達質粒修飾BMSCs移植促進大鼠急性脊髓損傷修復
2020-08-26 08:07:40石勇霞曉燕
實用骨科雜志 2020年8期
石勇,霞曉燕
(綿陽市人民醫院神經外科,四川 綿陽 621000)
急性脊髓損傷(spinal cord injury,SCI)多由直接暴力引起,包括交通事故傷、高處跌落傷等,具有創傷大、預后差的特點[1-2]。既往文獻報道,急性SCI往往導致神經細胞的壞死和凋亡,而干細胞移植是一種有效的治療方案[3-4]。但是,也有研究表明,干細胞移植具有分化程度低、易老化的特點[5]。因此,尋找一種有效的干細胞促分化方案,有效抑制干細胞的老化具有重要的意義。神經生長因子(nerve growth factor,NGF)是一種常見的神經營養因子,具有保護神經元、促進神經元的分化和神經纖維的定向生長的作用[6-7]。本研究利用Cas-9基因編輯技術,構建NGF的基因過表達載體,通過慢病毒轉染的方法,轉染骨髓間充質干細胞(bone marrow mesenchymal stem cells,BMSCs)。并且通過構建大鼠的脊髓損傷(spinal cord injury,SCI)動物模型,注射處理后的BMSCs,利用反轉錄聚合酶鏈反應(reverse transcriotion-polymerase chain reaction,RT-PCR)、Western blot、免疫熒光等方法檢測NGF的表達和神經的修復,以期為臨床上急性脊髓損傷的治療提供新的思路和方法。
1 材料與方法
1.1 實驗材料 胎牛血清購自以色列BI公司;改良Eagle培養基(dulbecco's modification of eagle's medium,DMEM)及0.25%胰酶Trypsin購自美國Invitrogen公司;TRIZOL購自美國Invitrogen公司;氯仿、酒精及逆轉錄試劑盒購自日本TAKARA公司;電泳儀、Real-time熒光定量PCR儀購自美國bio-rad公司。無特定病原體(specefic pathogen free,SPF)級SD大鼠購自濟南齊魯實驗動物中心[SCXK(魯)2018-2892]。月齡6~7個月,體重398~412 g。
1.2 實驗方法
1.2.1 人BMSCs的分離和培養 選取10名健康志愿者,男性6例,女性4例;年齡19~25歲,平均(22.13±3.23)歲。實驗方案獲得了本院倫理委員會的批準,所有志愿者均簽署知情同意書。骨髓穿刺針提取志愿者的新鮮骨髓,按照1︰3體積比加入含0.01 mol/L的胎牛血清、100 U/mL青霉素的DMEM細胞培養液,充分混勻后,移入25 cm2Hank培養瓶中。差速培養法培養,3 d后去掉培養瓶中的油脂等雜質,之后每3天更換1次細胞培養液。BMSCs細胞融合率達70%~80%后進行傳代培養。
1.2.2 BMSCs的鑒定 采用流式細胞儀檢測BMSCs的表面蛋白,CD 29、CD 44、CD 71、CD 90和CD 106等表面蛋白的表達可以為BMSCs的鑒定提供依據。另外,采用倒置光學顯微鏡也可以觀察BMSCs的細胞形態。
1.2.3 慢病毒NGF過表達載體的構建 利用Pubmed基因庫數據平臺,篩選人源性NGF的基因序列,基因序列為NC_000001.11,基因ID:4803,選擇pHBLV-U6-ZsGreen-PGK-Puro作為質粒的克隆載體。NGF過表達載體的克隆切入點是XhoI/BamHI,編號為ENST00000005188。
1.2.4 SD大鼠急性SCI動物模型構建 利用鉗夾法[8]構建大鼠SCI模型,SD大鼠采取腹腔注射水合氯醛麻醉,麻醉滿意后取俯臥位,常規消毒,備皮。取后背正中切口,逐層分離皮膚、筋膜和肌肉,直達棘突,盡量咬除T9~11棘突和椎弓板,暴露T10脊髓,將動脈瘤夾放置在脊髓上,寬度為脊髓的一半,突然釋放夾子使脊髓受到暴擊,30 s后取下動脈瘤夾,磷酸鹽緩沖液(phosphate buffered saline,PBS)沖洗,逐層縫合(見圖1)。
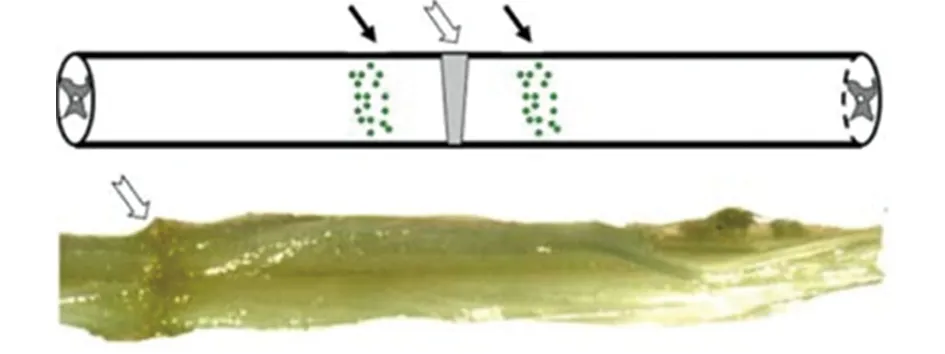
圖1 脊髓損傷模式圖和大體圖片
1.2.5 實驗動物分組 根據轉染的BMSCs類型,將實驗動物分成三組,每組5只SD大鼠。模型組:SD大鼠均未做任何處理;BMSCs組:SD大鼠SCI模型構建后,注射無慢病毒轉染的BMSCs;過表達質粒組:SD大鼠SCI模型構建后,注射NGF過表達質粒慢病毒轉染的BMSCs。
1.3 觀察指標
1.3.1 行為學觀察 三組SD大鼠均在損傷前和處理后12 h進行后肢功能恢復的行為學檢測,以運動功能(basso-beattie-bresnahan,BBB)評分作為指標,觀察指標為小鼠后肢踝關節活動度、腳掌腳背著地情況、軀干穩定性及尾巴的位置。
1.3.2 RT-PCR檢測NGF和凋亡標志因子Caspase-3的表達 各組大鼠干預2周后均按照動物倫理的要求無痛苦處死,取1 g修復的脊髓組織,加入1 mL Trizol試劑,充分研磨,抽提總RNA,然后根據逆轉錄試劑盒的要求將RNA逆轉錄為cDNA,根據TaKaRa熒光定量聚合酶鏈式反應(polymerase chain reaction,PCR)試劑盒說明書的要求,采用20 μL的反應體系,加入各樣品,進行熒光定量PCR的檢測。內參基因甘油醛-3-磷酸脫氫酶(glyceraldehyde-3-phosphate dehydrogenase,GAPDH)和NGF、Caspase-3引物由上海生工公司生產(見表1)。采用2-ΔΔCT的方法計算目的基因的相對表達量。

表1 目的基因的引物序列
1.3.3 Western blot檢測NGF和凋亡標志因子Caspase-3的表達 各組大鼠干預2周后均按照動物倫理的要求無痛苦處死,取1g修復的脊髓組織,加入1 mL放射免疫沉淀法緩沖液(radioimmunoprecipitation assay buffer,RIPA)裂解液,冰上裂解30 min,收集入1.5 mL的離心管中,在預冷4℃的離心機中離心,預設:5 000 r/min,5 min后提取上清,經95℃煮沸后收集蛋白備用。按照說明書制備濃縮膠10 mL,分離膠20 mL。上樣后電泳分離,轉膜后在4℃冰箱避光孵育過夜(>12 h)。NGF和Caspase-3一抗經一抗稀釋液1︰10 000稀釋后加入樣本離子膜。第2天用含吐溫20的磷酸鹽緩沖液(phosphate buffered solution tween,PBST)稀釋液洗膜,重復3次,加用山羊抗小鼠1 g G二抗經二抗稀釋液1︰1 000稀釋后孵育二抗1.5 h,用PBST稀釋液洗膜,重復3次,而后用二氨基聯苯胺(diaminobenzidine,DAB)顯影液處理樣本。Image J軟件可以用來分析蛋白條帶。
1.3.4 免疫熒光染色檢測NGF的定位表達 各組大鼠干預2周后均按照動物倫理的要求無痛苦處死,取1 g修復的脊髓組織,制備石臘組織,冰凍切片,二甲苯梯度處理,PBS溶液清洗后檸檬酸溶液浸潤20 min,PBS溶液清洗3次,BSA孵育30 min,避光孵育神經纖維標志物膠質纖維酸性蛋白(glial fibrillary acidic protein,GFAP)和神經元標志物神經元細胞核(neuronal nuclei,NeuN)蛋白一抗,4℃避光孵育過夜。PBS溶液清洗3次,辣根過氧化物酶(horseradish peroxidase,HRP)二抗避光孵育30 min,PBS溶液清洗3次,倒置光學顯微鏡下觀察。
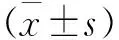
2 結 果
2.1 人BMSCs的培養和鑒定 原代BMSCs細胞成梭形或者多角形;培養至第2代后,呈旋渦狀或者輻射狀生長;流式細胞儀檢測第2代BMSCs的表面蛋白,CD 44和CD 29的表達為60.2%和58.3%;CD 34和CD 45的表達為3.4%和2.6%(見圖2)。
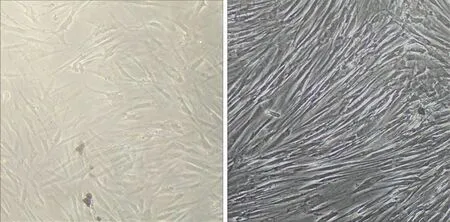
a 原代BMSCs b 第2代BMSCs
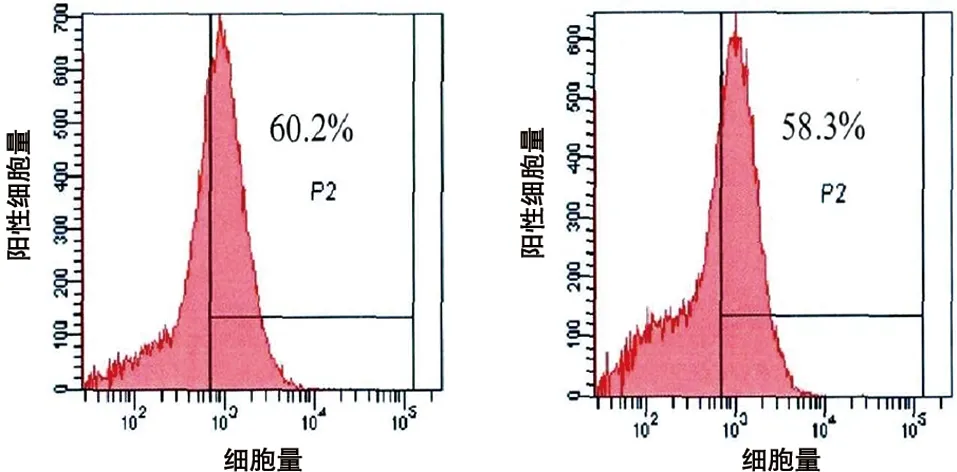
c 表面蛋白CD44的表達 d 表面蛋白CD29的表達
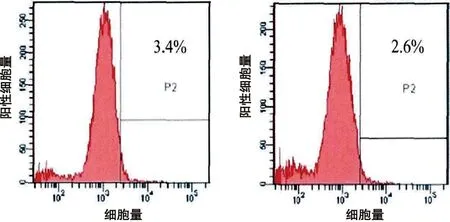
e 表面蛋白CD34的表達 f 表面蛋白CD45的表達
2.2 NGF過表達載體轉染效率 轉染效率=熒光視野下細胞數/白光視野下細胞數。結果顯示,慢病毒轉染效率為90%以上,轉染效率較高(見圖3)。
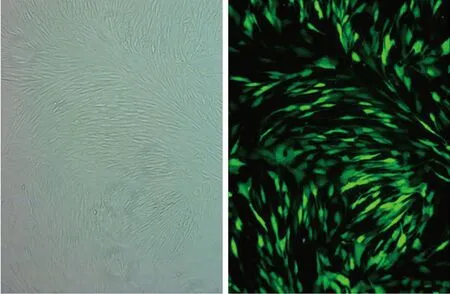
a 白光下視野 b 瑩光下視野
2.3 行為學檢測 術前SD大鼠BBB評分(21.91±4.7)分,分,模型組術后1 d的BBB評分降至最低,而BMSCs組要高于模型組(F=9.898,P<0.05),過表達質粒組高于模型組和BMSCs組(q值分別為2.198和2.004,P<0.05)。術后3 d的BBB評分明顯回升,術后12 d與術后14 d較平緩,比較差異無統計學意義(P>0.05,見圖4a)。
后肢踏空率的比較中,BMSCs組要明顯高于模型組(F=9.077,P<0.05),過表達質粒組要明顯高于模型組和BMSCs組(q值分別為2.129和2.099,P<0.05,見圖4b)。
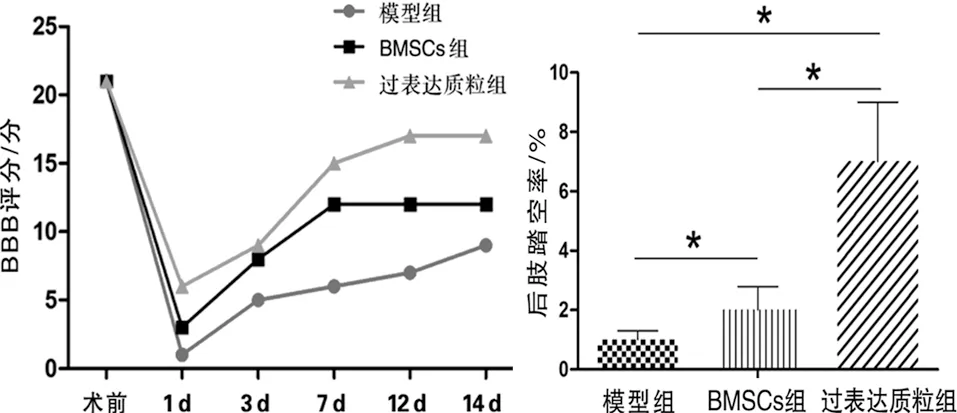
a 三組小鼠的BBB評分 b 三組小鼠的后肢踏空率
2.4 NGF和凋亡信號分子Caspase-3的表達量 RT-PCR結果顯示,BMSCs組NGF mRNA的相對表達水平要明顯高于模型組(F=8.997,P<0.05),過表達質粒組NGF mRNA的相對表達水平要明顯高于模型組和BMSCs組(q值分別為2.971和1.2.099,P<0.05)。Western blot的結果與RT-PCR結果相同(見圖5)。
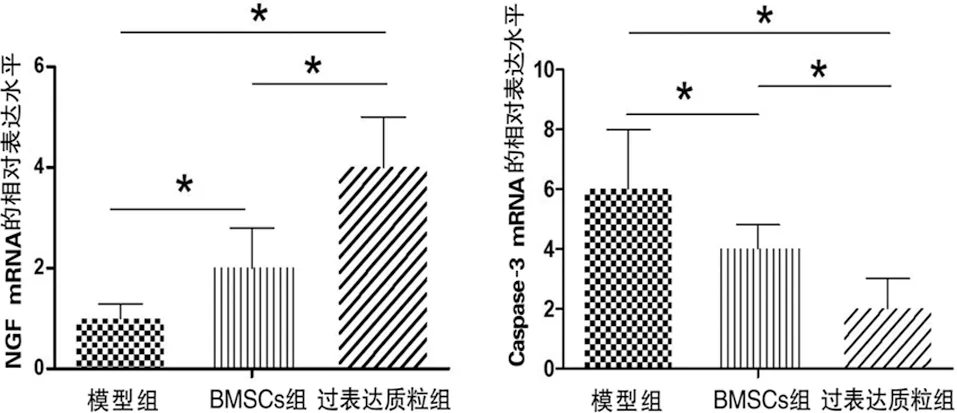
a NGF mRNA的相對表達水平 b Caspase-3 mRNA的相對表達水平
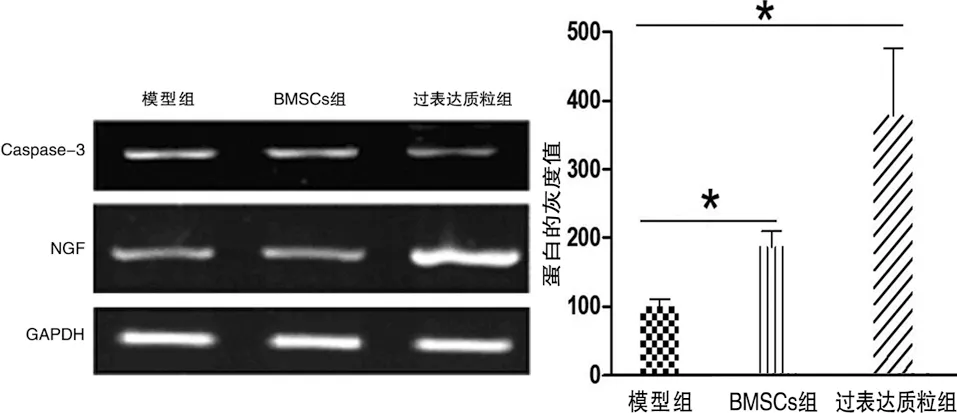
c Western blot檢測結果 d 蛋白的灰度值
2.5 免疫熒光染色結果 結果顯示,過表達質粒組脊髓修復程度要優于BMSCs組和模型組。可見斷裂間距明顯縮小,斷端修復較好(見圖6~7)。
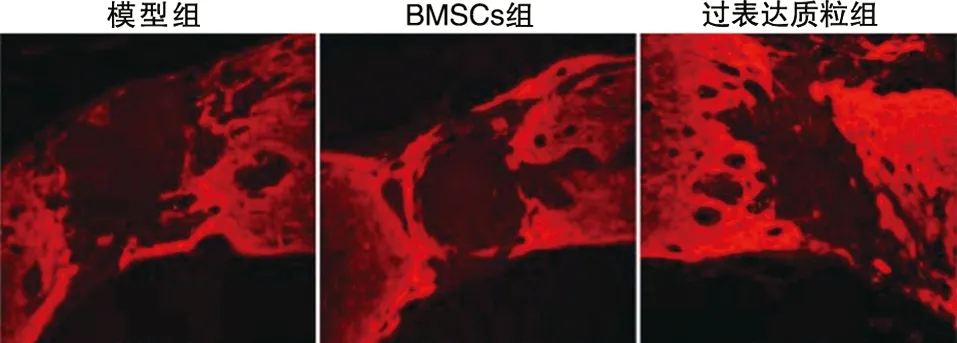
圖6 脊髓標本的GFAP染色結果(免疫熒光染色,×800)
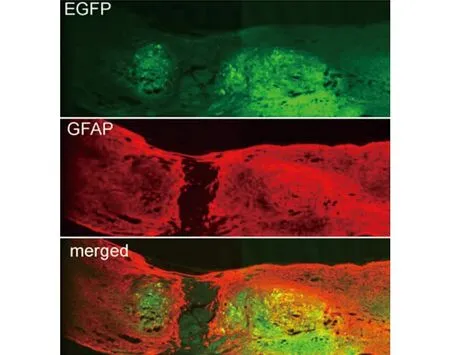
圖7 增強綠色熒光蛋白EGFP和GFAP共染結果(免疫熒光染色,×600)
2.6 NGF與神經元標志物NeuN共染結果 過表達質粒組NGF可與NeuN共染,神經元細胞量增多(見圖8)。
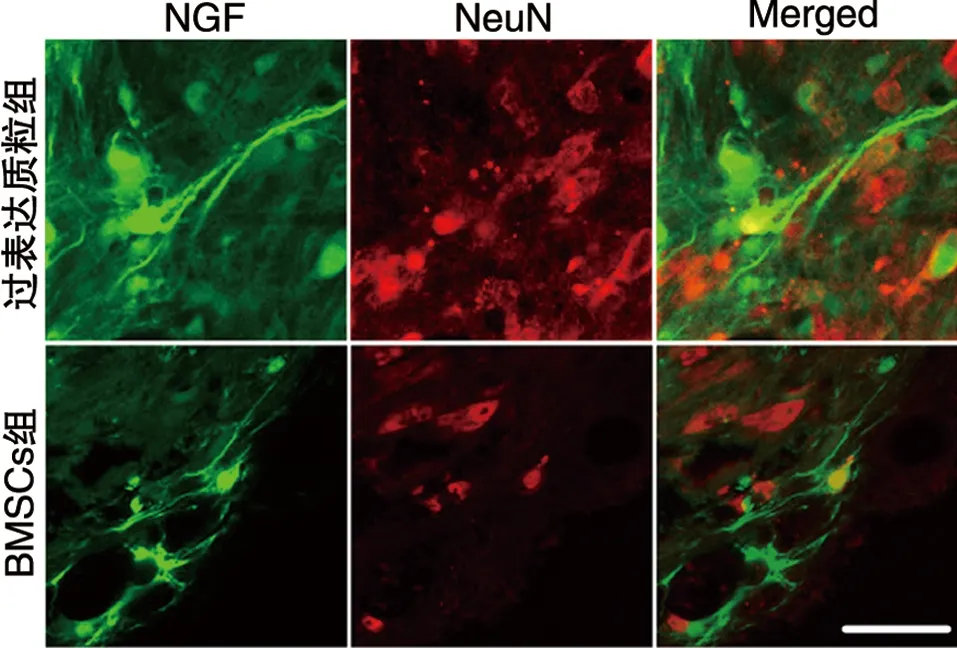
圖8 NGF與神經元標志物NeuN共染結果(免疫熒光染色,×600)
3 討 論
本研究通過構建NGF的過表達載體,通過慢病毒轉染技術轉染人BMSCs。并且利用鉗夾法構建SD大鼠的脊髓損傷動物模型,接種NGF過表達載體修飾后的BMSCs,利用RT-PCR和Western blot進行定量檢測,表明過表達載體組脊髓組織中可見NGF的過表達和凋亡標志因子Caspase-3的低表達。說明NGF基因過表達載體可以抑制脊髓組織神經元細胞的凋亡。另外,通過免疫熒光定位檢測,可見NGF蛋白可以與神經元細胞標志蛋白NeuN共染,與神經膠質纖維標志蛋白GFAP不共染。說明NGF過表達載體可以促進神經細胞元的增殖,進一步促進脊髓的修復。而且,我們也通過行為學檢測,利用BBB評分和后肢踏空率對動物的行為進行評估,結果表明,NGF過表達載體組小鼠的行為較另外兩組明顯改善。利用干細胞的多分化潛能對急性SCI進行治療一直是骨科研究的重點和難點問題[9-10]。Zhao等[11]將BMSCs細胞接種于急性SCI損傷動物模型中,得到良好的治療效果,但是也存在一些問題,比如干細胞的不定性分化和易老化等。這些問題限制的干細胞在脊髓修復領域的進一步應用。陳乃耀等[12]利用人源臍血間充質干細胞治療創傷性腦損傷(traumatic brain injury,TBI)大鼠模型,結果證明,人源臍血間充質干細胞可以促進TBI大鼠模型的神經損傷程度的改善,有利于損傷腦組織的修復。從而說明,可以利用人源性干細胞治療大鼠模型,具有良好的兼容性和有效性。既往文獻報道,NGF可以促進神經細胞的增殖和分化,并且抑制神經元細胞的凋亡[11]。本研究在前人的基礎上,采用Cas-9技術,篩選NGF的基因,通過擴增基因序列,促進NGF的蛋白表達,借助慢病毒轉染技術,將NGF過表達質粒轉染人源BMSCs,可以促進干細胞的定向分化,抑制凋亡。通過RT-PCR和Western blot等方法檢測NGF和凋亡標記因子Caspase-3的表達,結果表明,NGF轉染后可以抑制凋亡。原因可能與NGF的過度表達有關。并且利用免疫熒光染色定位NGF與神經元標志物NeuN的共染,結果表明,NGF過表達質粒可以促進NeuN的表達,說明NGF可以促進BMSCs向神經元細胞分化。其原因可能與NGF的促神經元分化相關。而且,既往文獻中也對空白質粒慢病毒轉染對實驗結果的影響進行了相應研究,他們的研究結果表明空白質粒對研究結果無影響,結果說明空白質粒不會影響研究結果[13-14]。本研究結果與前人研究結果相同,空白質粒慢病毒轉染BMSCs,不會影響BMSCs的分化和增殖。
為了進一步評估NGF過表達質粒轉染BMSCs后對SCI動物模型脊髓損傷的修復作用,本研究還利用免疫熒光染色共染神經膠質纖維標志蛋白GFAP,結果表明,過表達質粒組脊髓修復程度要優于BMSCs組和模型組。可見斷裂間距明顯縮小,斷端修復較好。而且通過行為學檢測,利用BBB評分和后肢踏空率對動物的行為進行評估,結果表明,NGF過表達載體組小鼠的行為較另外兩組明顯改善。其原因可能為神經元的增殖,促進神經膠質纖維的生成,從而對脊髓組織起到很好的修復效果,進一步影響SCI動物模型的行為。
綜上所述,NGF過表達質粒轉染后可以促進BMSCs向神經元細胞分化,促進SCI動物模型的功能恢復。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
海峽科技與產業(2016年3期)2016-05-17 04:32:12
