基于SSR的光敏型飼草高粱分子輔助育種體系
2020-08-15 02:42:02牛皓平俊愛王玉斌張福耀呂鑫李慧明楚建強
中國農業科學 2020年14期
牛皓,平俊愛,王玉斌,張福耀,呂鑫,李慧明,楚建強
基于SSR的光敏型飼草高粱分子輔助育種體系
牛皓1,2,平俊愛1,2,王玉斌1,2,張福耀1,2,呂鑫1,2,李慧明1,2,楚建強1,2
(1山西農業大學(山西省農業科學院)高粱研究所/高粱遺傳與種質創新山西省重點實驗室,山西晉中 030600;2農業部黃土高原作物基因資源與種質創制重點實驗室,太原 030031)
【】利用分子標記技術對光敏型飼草高粱新品種傳統選育方法進行改良,構建分子輔助育種體系,從而提高品種選育效率,降低育種成本,為形成新的育種體系奠定理論基礎。以光敏型(不抽穗)飼草高粱品種晉光1R去雄后為母本、非光敏型(抽穗)飼草高粱BMRC-3-2為父本進行雜交,構建F2群體。按照光敏與非光敏性狀將群體分為兩類,各選30株,取葉片提取DNA,利用微衛星分子標記技術(SSR)和集團分離分析法(BSA)對晉光1R/BMRC-3-2的F2群體進行光敏感基因定位分析,以及特異性SSR引物的設計、篩選;將篩出的特異性引物對F1雜交種(以晉光1R為親本選育的高代穩定恢復系作為父本與其他不育系測配得到的F1代雜交種)進行光敏性鑒定,通過與田間表型對照,確定最終目標引物,從而構建新的光敏型飼草高粱新品種選育體系。經過SNP index分析,選取99%閾值線最高點前后約250 kb的區域作為性狀相關的候選區域,區域總長度為500 kb,共400個SNP位點,對這些SNP位點注釋,其中非同義突變或者stop-gain或者stop-loss的SNP位點有6個,最終確定2個候選基因與高粱光敏感性相關,這兩個基因位于高粱第7染色體。利用微衛星標記技術開發引物51對,對光敏和非光敏的親本及F1雜交種進行毛細管電泳檢測,選定1對特異性引物70.2-3,結果表明,以251 bp處出現“單峰”為判斷依據,引物70.2-3對50個非光敏型F1雜交種的鑒選準確率達到100%,所有材料均在251 bp附近出現峰值。以214和251 bp兩處附近出現“雙峰”為判斷依據,該引物對另外50個光敏型F1雜交種鑒選的準確率達到90%,其中F1-69、F1-70、F1-71這3個材料在214和262 bp附近出現“雙峰”;F1-81僅在251 bp處有“單峰”,而F1-86則在251和232 bp兩處附近出現“雙峰”。發現了控制高粱光敏性的候選基因和,位于高粱第7染色體810 000—1 310 000 bp,獲得特異性引物70.2-3,可在一定程度上改良傳統育種手段,用實驗室驗證替代田間測交觀察,節省了育種成本,提高了育種效率。
飼草高粱;光敏型;分子標記
0 引言
【研究意義】飼草高粱是中國主要青貯飼料,由于其再生能力強、產量高、蛋白質豐富、易消化等特點,被廣泛應用于農牧飼喂及漁業養殖領域[1-2]。光敏感特性是影響飼草高粱產量和品質的主要因素[3-4]。在北方夏季長日照地區,當光照時長超過12.3 h時,具有光敏感特性的飼草高粱不抽穗結實,只進行營養生長,其高度可超過4 m,部分材料在新疆地區可長至6 m,因而產量、蛋白質含量比普通飼草高粱高,若部分替代玉米作為哺乳期奶牛飼料,在增加牛奶乳脂肪含量上有顯著效果[5-9]。然而,在光敏型飼草高粱品種選育的過程中,穩定后代是否具有光敏特性,必須將其與普通的不育系測交,第二年種植于大田中,秋天觀察抽穗性狀才能判斷,選育過程中物力、人力、地力投入較多,育種成本非常高,因此,利用分子標記輔助育種技術對傳統育種方法進行改良,提高育種效率顯得尤為重要。【前人研究進展】生物質產量是將木質素轉化為生物燃料或生物能源的生物質或生物能源作物最重要的屬性之一,生長期是生物質產量的首要決定因素。因此,假設環境條件允許產量潛力得以發揮,光敏型高粱積累最多的生物質,可達到普通高粱的2倍以上[10]。目前,已知與高粱光敏性相關的基因共有6個,為Ma—Ma[10-11]。早在1945年,美國學者Quinby等[12]認為有3個基因影響高粱的花期和成熟期,分別為Ma、Ma和Ma。Ma、Ma在沒有Ma的情況下不會表達,Ma在顯性Ma存在的情況下也不會表達。同時Ma的作用受光周期的影響,開花時間控制著葉片的數量、生長時間和植株的最終大小。1965年,Quinby[13-14]又發現了第4個與高粱成熟期相關的基因,并將它命名為Ma,認為這4個基因的表達受環境的影響,特別是受光周期和溫度的影響,且花期開始的早晚取決于品種的成熟基因型。20世紀90年代,Morgan等[11]、Major等[15]從環境、溫度等因素對這一概念又進行了闡述,之后有關高粱光敏基因的研究豐富起來。Childs等[16]認為Ma是第一個被克隆的與高粱成熟期相關的基因,并且有3個等位基因分別為Ma、ma、ma。Rooney等[17-18]和Morgan等[19]將高粱恢復系EBA-3去雄后與其他非光敏(photo-insensitive)恢復系雜交,發現8個F2群體中PS/PI分離比率為9﹕7,表明2個獨立的基因位點在互補顯性上位中相互作用,而這兩個與光周期敏感度相關的基因與先前的Ma、Ma、Ma和Ma不同,從而定名為Ma和Ma,并發明了基于Ma/Ma系統的生物質能源高粱的選育方法。Murphy等[20]認為蛋白PRR37控制高粱的開花時間。Yang等[21-22]認為高粱CONSTANS是一種開花的激活劑,在開花抑制劑PRR37的長時間內在轉錄后受到抑制,有助于光敏感型短日照高粱的開花。Wang等[23]對Ma的進化做了分析。另外,針對飼草高粱生物產量與光敏感特性的研究,Wolabu等[24-25]認為不同類型的高粱在溫帶地區的應用主要取決于其開花時間的控制,光周期不敏感的高粱用于糧食生產,而光周期敏感基因型則用于飼料和生物量原料生產。關于光敏感基因定位的研究,Murphy等[20]將Ma定位在高粱第6染色體上的標記Xtxi62和Xtxi58之間約86 kb的區段內。Mullet等[26]將Ma定位在高粱第2染色體67 923 811—68 393 290 bp,認為該基因可能通過抑制由光敏色素B和C介導的調節高粱光照依賴性產物來調控高粱的開花時間。Ma是第一個被成功克隆的調控高粱感光性的基因,被定位于高粱第1染色體長臂的末端,在60 910 479—60 917 763 bp。Ma至少有3種等位基因,分別為顯性Ma、隱性Ma和一種特殊的隱性基因Ma[27-29]。Ma被定位在高粱第10染色體,與RFLP標記txs1163相鄰[30-31]。Ma被定位到高粱第1染色體6 762 743—6 767 650 bp,在遺傳圖譜上的位置為23—26 cM,對應的基因編碼光敏色素C[26]。Ma被定位在高粱第6染色體39 379 760—42 610 705 bp,它和Ma均受Ma、Ma的遺傳上位性影響,通過抑制一系列成花素基因的表達,使高粱在長日照條件下開花時間延遲。Ma是調控高粱籽粒數量、株高和抽穗期的基因——[26]。牛皓等[32]對144對SSR引物(包括PRR37(Ma)、GHD7(Ma)、PHYB(Ma)、PHYC(Ma)等基因對應的引物)進行篩選,篩出4對疑似光敏感引物,但經過與F1雜交種擴大群體驗證,發現它們并不是特異性引物,不能將光敏和非光敏的材料區分開,因而不具備生產使用價值。【本研究切入點】高粱光敏感基因的理論研究豐富,但利用已有成果形成育種體系應用于田間育種的研究鮮有報道。【擬解決的關鍵問題】本研究以具有光敏感特性(photosensitive)的飼草高粱恢復系晉光1R和普通非光敏(photo-insensitive)飼草高粱恢復系BMRC-3-2為研究對象,通過雜交構建F2群體,利用微衛星分子標記技術,定位光敏基因,篩選特異性引物,為今后改良高粱品種傳統育種方式,提高育種效率,降低育種成本,進而形成新的育種體系奠定理論基礎。
1 材料與方法
1.1 試驗材料
親本材料:具有光敏基因的飼草高粱恢復系晉光1R,普通飼草高粱恢復系BMRC-3-2。
F2群體:2017年,在山西榆次以光敏恢復系晉光1R為母本,非光敏恢復系BMRC-3-2為父本,經人工去雄有性雜交得到F0種子;2017年冬,在海南三亞種植F0種子,并自交,得到F1種子,2018年,獲得晉光1R/BMRC-3-2的F2群體。
F1雜交種:以晉光1R為親本選育的高代穩定恢復系作為父本,與其他不育系測配得到的F1雜交種。
部分SSR標記序列來源于NCBI網站,部分由上海派森諾生物科技有限公司依據候選基因序列設計合成。
材料種子由山西省農業科學院高粱研究所提供。所有試驗材料均種植于山西省農業科學院高粱研究所東白試驗基地。田間管理與其他普通高粱一致,2018年8月底調查抽穗情況。
1.2 表型鑒定及遺傳分析
取樣前,對親本及群體單株進行光敏和非光敏性狀調查統計。根據表型鑒定結果計算分離比例,并進行卡方檢測,確定光敏性狀在F2群體中的分離情況,用于遺傳分析。
1.3 DNA提取及近等基因池的構建
為確保取樣的準確性,在8月底高粱抽穗尾期,按照光敏(不抽穗)與非光敏(抽穗)將F2分兩類,每類隨機各選30株,分別掛牌標記,取頂部新鮮幼葉放入2 ml離心管中,液氮速凍,由上海派森諾生物科技有限公司提DNA及進行后續相關試驗。同時,將1.1內容中F1雜交種,也按照光敏與非光敏分兩類,每類隨機各取50株頂端新鮮幼葉放入2 ml離心管中,液氮速凍,由上海派森諾生物科技有限公司進行微衛星標記驗證試驗。
1.4 BSA試驗
構建基于Illumina HiSeq測序平臺,利用第二代測序技術(next-generation sequencing,NGS),對插入片段為400的文庫進行雙末端(paired-end,PE)測序。通過對目標性狀區域分析,確定基因位置。
1.5 微衛星標記驗證試驗
將F1雜交種DNA進行毛細管電泳檢測,驗證特異性引物。
2 結果
2.1 表型鑒定和遺傳分析
晉光1R與BMRC-3-2雜交,對F2進行性狀調查,其中,251株不抽穗,169株抽穗,經c2檢驗光敏與非光敏(PS/PI)的分離比例符合9﹕7(表1)。
2.2 基因初步定位及功能注釋
經過測序,選取99%閾值線最高點前后約250 kb的區域作為性狀相關的候選區域,區域總長度為500 kb,這個范圍內滿足條件的SNP位點共400個,對這些SNP位點注釋,其中,非同義突變或者stop-gain或者stop-loss的SNP位點有6個,分布在4個基因。關聯區域位于第7染色體810 000—1 310 000 bp(圖1)。

表1 F2光敏性狀的遺傳模式
c20.05,1=3.84

圖1 子代混池 SNP index和△(SNP index)在染色體上的分布
4個候選基因分別為、、和。通過與GO、KEFG、Swiss Prot、NR等數據庫比對注釋,其中和都是剪切因子基因,因而,認為它們可能就是與飼草高粱光敏感相關的基因。
2.3 特異性SSR引物的獲得及分子輔助育種體系的建立
在F1材料的DNA中選取光敏與非光敏單株各2株,根據候選基因序列設計合成的引物共51對,進行熒光定量PCR,每組引物前2條帶為非光敏F1,后2條帶為光敏F1(圖2)。

M:DL2000 DNA Maker;1—51:70.2-1、70.2-2、70.2-3、70.2-152-1、70.2-152-2、70.2-152-3、71.2-229-1、71.2-229-2、71.2-229-3、72.2-1、72.2-2、72.2-3、73.2-1、73.2-2、73.2-3、75.2-833-1、75.2-833-2、75.2-833-3、75.2-392-1、75.2-392-2、75.2-392-3、75.2-384-1、75.2-384-2、75.2-384-3、75.2-036-1、75.2-036-2、75.2-036-3、75.2-258-1、75.2-258-2、75.2-258-3、77.2-751-1、77.2-751-2、77.2-751-3、77.2-050-1、77.2-050-2、77.2-050-3、78.2-409-1、78.2-409-2、78.2-409-3、78.2-005-1、78.2-005-2、78.2-005-3、79.2-488-1、79.2-488-2、79.2-488-3、79.2-924-1、79.2-924-2、79.2-924-3、79.2-797-1、79.2-797-2、79.2-797-3
根據圖2結果,將個別差異極不明顯的引物排除,其余引物擴大樣本至100個F1材料進行毛細管電泳驗證。結果表明,僅有引物70.2-3可以區分光敏與非光敏F1材料(圖3)。非光敏型F1雜交種在251 bp 附近出現“單峰”;而光敏型則在214和251 bp兩處附近出現“雙峰”;光敏親本和非光敏親本則分別在214和251 bp附近出現“單峰”。以251 bp處出現“單峰”為判斷依據,引物70.2-3對50個非光敏型F1雜交種的鑒選準確率達到100%,所有材料均在251 bp附近出現峰值。以214和251 bp兩處附近出現“雙峰”為判斷依據,該引物對另外50個光敏型F1雜交種鑒選的準確率達到90%,其中F1-69、F1-70、F1-71這三個材料在214和262 bp附近出現“雙峰”;F1-81僅在251 bp處有“單峰”而F1-86則在251和232 bp兩處附近出現“雙峰”。據此,認為所選引物70.2-3為特異性引物,可以區分光敏和非光敏雜交種,可以作為田間生產指導。其序列為FORWARD PRIMER:5′-CCTCCTCTTCCTCGGATAGC-3′和REVERSEPRIMER:5′-ATGATCGGTGGT TG GAAGAG-3′。

Sample1和Sample2為非光敏型F1雜交種,Sample3和Sample4為光敏型F1雜交種,Sample5為光敏親本晉光1R,Sample6為非光敏親本BMRC-3-2
綜上所述,對傳統的光敏感飼草高粱選育體系進行改良,利用實驗室篩選代替田間測交,形成新的分子輔助育種體系(圖4)。
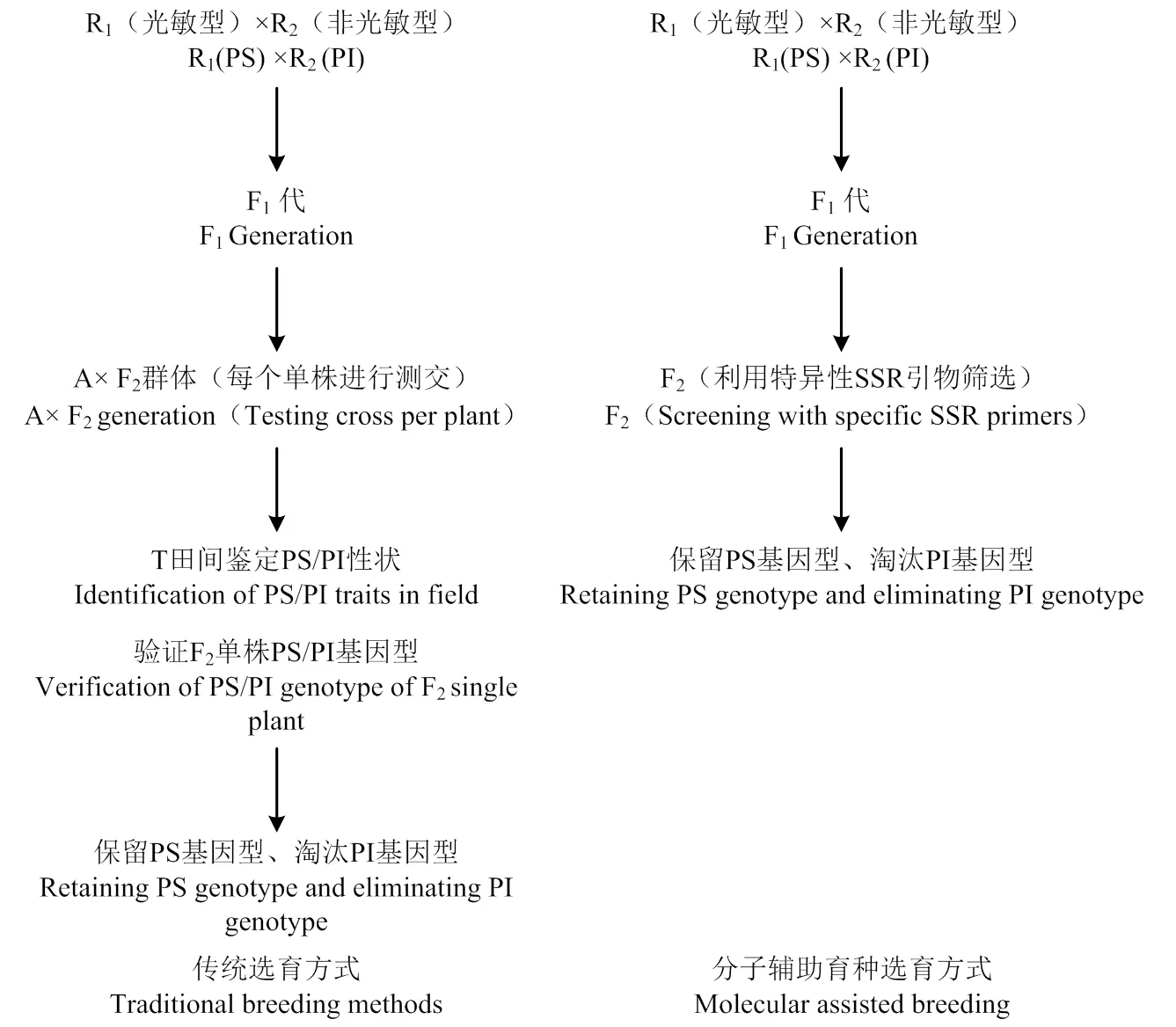
圖4 新舊育種體系對比示意圖
3 討論
3.1 傳統選育方式
光敏型飼草高粱恢復系的選育,以傳統高粱恢復系選育方法為基礎,選擇具有光敏特性的飼草高粱恢復系與普通非光敏但具有其他優良性狀的飼草高粱恢復系人工去雄有性雜交,獲得F0種子,通過南繁加代也可下一年種植大田得到F1,F1套袋自交后第二年得到F2群體。從這一步開始傳統的育種方法需另選不育系A與選出的F2群體單株一一測交,得到雜交種T,翌年田間種植后再觀察抽穗情況,從而判定上年測交F2群體中哪株為光敏型。
3.2 分子輔助育種選育方式
分子輔助育種選育方式與傳統育種方式在獲得F2群體的方法上保持一致,但將工作量最繁瑣的田間測交驗證這一步用實驗室熒光定量PCR方法取代,利用篩選的特異性SSR引物70.2-3將所有F2群體中的個體進行毛細管電泳檢測,觀察214和251 bp處的“峰值”情況,兩處均出現“峰”則判定為光敏型,非光敏型則淘汰。結果表明,該引物篩選光敏材料準確度非常高,但仍有極少部分材料出現峰值位移,這與部分“中間型”材料取樣分類誤差有關。同時,候選基因和定位在第7染色體,與目前所有已知的與高粱光敏感性相關的基因位置都不同,是否是新的光敏基因,其調控機理如何還需進一步研究確定,但F2群體中光敏與非光敏(PS/PI)分離比例為9﹕7,這說明飼草高粱光敏感特性是由2個獨立的互補顯性基因控制,這與Rooney[17]研究結果相同。
3.3 分子輔助育種體系優點
本研究創造的光敏型飼草高粱分子輔助育種體系是對傳統育種體系的改良,優點主要體現在:1)節約育種時間;2)節省育種成本。通過對比可發現,傳統的育種體系比分子輔助育種體系的育種年限要多1年,而多出來的這一年必然會引起育種成本的增加。假設F2群體共200株,每株均與不育系A測交,收獲測交種子200份,第二年以每15 m2小區種6行,每2行種1個材料來計算,大約需要67個小區0.1 hm2,以2019年每公頃地育種成本來計算,需約6 000元,而實驗室熒光定量PCR耗材費用不超過2 000元。如果F2群體數量龐大,后者將節省更多的育種成本。
綜上所述,調控高粱抽穗期性狀的基因很多,而且等位基因也很豐富,并且影響不同雜交組合產生后代的抽穗期性狀的QTL位點各不相同,調控高粱抽穗期性狀的基因之間互作方式也是多種多樣。因此,盡可能多的發掘調控高粱抽穗期性狀的基因位點,明確基因功能及互作模式,對于揭示高粱光敏感特性的機理、提高育種效率、節約育種成本有著重要的意義。
4 結論
發現控制高粱抽穗期的候選基因和,位于高粱第7染色體810 000— 1 310 000 bp。同時特異性SSR引物70.2-3的獲得,在飼草高粱光敏性鑒定上有極高的準確率,基于該引物構建的光敏型飼草高粱選育體系,利用實驗室驗證替代田間測交觀察,節省育種成本,提高育種效率。
[1] 平俊愛, 謝國強, 張福耀, 何余湧, 吳志勇, 王榮民, 呂鑫, 杜志宏, 李慧明, 牛皓, 王玉斌. 晉牧1號在南方紅壤區的產草量: 營養價值及飼養效果研究. 安徽農業科學, 2018, 46(35): 94-96.
PING J A, XIE G Q, ZHANG F Y, HE Y Y, WU Z Y, WANG R M, Lü X, DU Z H, LI H M, NIU H, WANG Y B. Breeding and characteristic ofבJinmu1’., 2018, 46(35): 94-96. (in Chinese)
[2] 平俊愛, 張福耀, 牛皓,楊慧勇, 呂鑫, 杜志宏, 李慧明, 王玉斌. 基于SSR標記的飼草高粱種質資源遺傳多樣性分析. 分子植物育種, 2018, 16(14): 4663-4670.
PING J A, ZHANG F Y, NIU H, YANG H Y, Lü X, DU Z H, LI H M, WANG Y B. Genetic diversity analysis of germplasm resources of forage sorghum based on SSR marker., 2018, 16(14): 4663-4670. (in Chinese)
[3] Bhosale S U, Benjamin S, Rattunde H F W, Weltzien E, Haussmann B I, Haussmann C T, Ramu P, Cuevas H E, Paterson A H, Melchinger A E, Parzies H K. Association analysis of photoperiodic flowering time genes in west and central[(L.) Moench]., 2012, 12: 32-41.
[4] Casto A L, Mattison A J, Olson S N, Thakran M, Rooney W L, Mullet J E. Maturity2, a novel regulatorof flowering time in, increases expression of SbPRR37 and SbCO in long days delaying flowering., 2019, 14(4): e0212154.
[5] Sukumaran S, Li X, Li X R, Zhu C S, Bai G H, Perumal R, Tuinstra M R, Prasad P V V, Mitchell S E, Tesso T T, Yu J. QTL mapping for grain yield, flowering time, and stay-green traits in sorghum with genotyping-by-sequencing markers., 2016, 56(4): 1429-1442.
[6] Tuinstra M R, Grote E M, Goldsbrough P B, Ejeta G. Genetic analysis of post-flowering drought tolerance and components of grain development in(L.) Moench., 1997, 3(6): 439-448.
[7] Spindel J E, Dahlberg J, Colgan M, Hollingsworth J, Sievert J, Staggenborg S H, Hutmacher R, Jansson C, Vogel J P. Association mapping by aerial drone reveals 213 genetic associations for Sorghum bicolor biomass traits under drought., 2018, 19: 679-696.
[8] Wang Y H, Upadhyaya H D, Burrell A M, Sahraeian S M, Klein R R, Klein P E. Genetic structure and linkage disequilibrium in a diverse, representative collection of the C4 model plant., 2013, 3(5): 783-793.
[9] Harper M T, Oh J, Giallongo F, Lopes J C, Roth G W, Hristov A N. Using brown midrib 6 dwarf forage sorghum silage and fall-grown oat silage in lactating dairy cow rations., 2017, 100(7): 5250-5265.
[10] Mullet J E, Rooney W L, Klein P E, Bryan D M, Bryan R M, Brady J A. Discovery and utilization of sorghum genes (Ma5/Ma6). 13/744405[P].2013-09-12.
[11] Morgan P W, Finlayson S A, Lee I J, Childs K L, He C J, Creelman R A, Drew M C, Mullet J E. Regulation of circadianly rhythmic ethylene production by phytochrome b in sorghum., 1997(1): 105-111.
[12] Quinby J R, Karper R E. The inheritance of three genes that influence time of floral initiation and maturity date in Milo1., 1945(11): 916-936.
[13] Quinby J R. Fourth maturity gene locus in sorghum., 1965(6): 516-518.
[14] Quinby J R. The maturity genes of sorghum., 1967(19): 267-305.
[15] Major D J, Rood S B, Miller F R. Temperature and photoperiod effects mediated by the sorghum maturity genes., 1990, 30(2): 305-310.
[16] Childs K L, Lu J L, Mullet J E, Morgan P W. Genetic regulation of development in: X. Greatly attenuated photoperiod sensitivity in a phytochrome-deficient sorghum possessing a biological clock but lacking a red light-high irradiance response., 1995, 108(1): 345-351.
[17] Rooney W L, Jürg B, Bean B, Mullet J E. Designing sorghum as a dedicated bioenergy feedstock., 2007, 1(2): 147-157.
[18] Rooney W L, Aydin S. Genetic control of a photoperiod-sensitive response in(L.) Moench., 1999, 39(2): 397-400.
[19] Morgan P W, Finlayson S A, Childs K L, Mullet J E, Rooney W L. Opportunities to improve adaptability and yield in grasses., 2002, 42(6): 1791-1799.
[20] Murphy R L, Klein R R, Morishige D T, Brady J A, Rooney W L, Miller F R, Dugas D V, Klein P E, Mullet J E. Coincident light and clock regulation of pseudo response regulator protein 37 (PRR37) controls photoperiodic flowering in sorghum., 2011, 108(39): 16469-16474.
[21] Yang S S, Weers B D, Morishige D T, Mullet J E. CONSTANS is a photoperiod regulated activator of flowering in sorghum., 2014, 14(1): 148-162.
[22] Yang S S, Murphy R L, Morishige D T, Klein P E, Rooney W L, Mullet J E. Sorghum phytochrome B inhibits flowering in long days by activating expression of SbPRR37 and SbGHD7, repressors of SbEHD1, SbCN8 and SbCN12., 2014, 9(8): e105352.
[23] Wang Y, Tan L B, Fu Y C, Zhu Z F, Liu F X, Sun C, Cai H W. Molecular evolution of the sorghum maturity gene Ma3., 2015, 10(5): e0124435.
[24] Wolabu T W, Tadege M. Photoperiod response and floral transition in sorghum., 2016, 11(12): e1261232.
[25] Wolabu T W, Zhang F, Niu L F, Kalve S, Mathur P B, Muszynski M G, Tadege M. Three FLOWERING LOCUS T-like genes function as potential florigens and mediate photoperiod response in sorghum., 2016, 210(3): 946-959.
[26] Mullet J E, Rooney W L. Method for production of sorghum hybrids with selected flowering times. 13/886130[P].2016-08-30.
[27] Childs K L, Miller F R, Cordonnier Pratt M M, Pratt L H, Morgan P W, Mullet J E. The sorghum photoperiod sensitivity gene, Ma3encodes a phytochrome B., 1997, 113(2): 611-619.
[28] Childs K L, Cordonnier Pratt M M, Pratt L H, Morgan P W. Genetic regulation of development in Sorghum bicolor: VII. Ma3Rflowering mutant lacks a phytochrome that predominates in green tissue., 1992, 99(2): 765-770.
[29] Childs K L, Pratt L H, Morgan P W. Genetic regulation of development in: VI. The Ma3Rallele results in abnormal phytochrome physiology., 1991, 97(2): 714-719.
[30] Hart G E, Schertz K F, Peng Y, Syed N H. Genetic mapping of(L.) moench QTLs that control variation in tillering and other morphological characters., 2001, 103(8): 1232-1242.
[31] Chantereau J, Trouche G, Rami J F, Deu M, Barro C, Grivet L. RFLP mapping of QTLs for photoperiod response in tropical sorghum., 2001, 120(2):183-194.
[32] 牛皓, 平俊愛, 張福耀, 呂鑫, 杜志宏, 李慧明, 王玉斌. 基于SSR的高粱光敏特性分析. 安徽農業科學, 2018, 46(35): 103-105.
NIU H, PING J A, ZHANG F Y, Lü X, DU Z H, LI H M, WANG Y B. Analysis on the photosensitivity of sorghum based on SSR., 2018, 46(35): 103-105. (in Chinese)
Molecular Aided Breeding System of Photosensitive Forage Sorghum Based on SSR
NIU Hao1,2, PING JunAi1,2, WANG YuBin1,2, ZHANG FuYao1,2, Lü Xin1,2, LI HuiMing1,2, CHU JianQiang1,2
(1Sorghum Institute of Shanxi Agricultural University (Shanxi Academy of Agricultural Sciences)/Shanxi Key Laboratory of Sorghum Genetic and Germplasm Innovation, Jinzhong 030600, Shanxi;2Key Laboratory of Crop Gene Resources and Germplasm Enhancement on Loess Plateau, Ministry of Agriculture, Taiyuan 030031)
【】The aim of this study is to use molecular marker to construct a breeding system to improve the traditional breeding method,thereby reducing breeding costs and improving breeding efficiency.At the same time, it lays a theoretical foundation for the formation of a new breeding system.【】The F2population was constructed by crossing the photosensitive(non-heading) forage sorghum variety Jinguang 1R as female and the photoinsensitive(heading) sorghum variety BMRC-3-2 as male.According to heading and non-heading phenotypes, F2population was divided into two groups. Thirty plants were selected from each group and DNA was extracted from leaves,SSR and BSA were used to map photosensitive genes in F2population of Jinguang 1R/BMRC-3-2 and screen for specific SSR primers.The specific primers were used to identify the photosensitivity trait in F1generation(F1Hybrids breeded from high generation stable restorer line with Jinguang 1R as parent and matched with other male sterile lines), and the final target primers were determined by comparing with the phenotype in the field, so as to construct a new photosensitive breeding system for forage sorghum.【】After SNP index analysis, the region of about 250 Kb before and after the highest point of 99% threshold line was selected as the candidate region for trait correlation. The total length of the region was 500 kb, with 400 SNP loci, and six of them were non-synonymous mutations or stop gain or stop loss SNP loci.Finally, two candidate genes were identified to be related to photosensitivity, which were located on chromosome 7.A pair of specific primers 70.2-3 was developed by SSR. The primerscan distinguish between heading and non-heading sorghum plants to a great extent.The results showed that according to the "single peak" at 251bp, the accuracy of primers 70.2-3 in the identification and selection of 50 non-photosensitive F1hybrids reached 100%, and all the materials showed a peak value near 251bp.Based on the "double peaks" in the vicinity of 214bp and 251bp, the accuracy rate of the primer for the identification and selection of the other 50 photosensitive F1hybrid species reached 90%. Three materials, No. F1-69, F1-70 and F1-71, showed "double peaks" near 214 bp and 262 bp. Material F1-81 has a "single peak" at 251bp, while material No. 86 has a "double peak" near 251 bp and 232 bp.【】In this study, we revealed that the candidate genes controlling heading date of sorghum were LOC8068537 and LOC8068548, which were between 810000 and 1310000 bp on chromosome 7 of sorghum. The acquisition of specific primers 70.2-3 improved the traditional breeding methods. The primer validation in laboratory could replace the field crossing observation, saving the breeding cost and improving the breeding efficiency.
foragesorghum;photosensitive type; molecular markers

10.3864/j.issn.0578-1752.2020.14.004
2019-06-06;
2019-08-08
山西省農業科學院優勢課題組自選項目(YYS1706)、現代農業產業技術體系建設專項資金(CARS-06)、山西省農業科學院生物育種工程(17yzgc031)、山西省重點研發計劃重點項目(201703D211002-9-1)、釀造專用高粱育種及利用山西省科技創新重點團隊運行補助(201805D131012-6)
牛皓,E-mail:nkyglsnh@126.com。通信作者平俊愛,E-mail:pingja1029@163.com
(責任編輯 李莉)
