藏黃牛與宣漢黃牛心臟miRNA表達(dá)譜比較
2020-08-15 02:08:28陳露露王會王吉坤王嘉博柴志欣陳智華鐘金城
中國農(nóng)業(yè)科學(xué) 2020年8期
陳露露,王會,王吉坤,王嘉博,柴志欣,陳智華,鐘金城
藏黃牛與宣漢黃牛心臟miRNA表達(dá)譜比較
陳露露,王會,王吉坤,王嘉博,柴志欣,陳智華,鐘金城
(西南民族大學(xué)青藏高原研究院/青藏高原動物遺傳資源保護(hù)與利用四川省、教育部重點(diǎn)實(shí)驗(yàn)室,成都 610041)
【目的】miRNA作為一類非編碼RNA,廣泛參與機(jī)體多種生命活動,研究旨在挖掘miRNA在藏黃牛和宣漢黃牛心臟組織中的差異miRNA,為進(jìn)一步研究藏黃牛低氧適應(yīng)的分子機(jī)制提供基礎(chǔ)數(shù)據(jù)。【方法】隨機(jī)選取健康的藏黃牛和宣漢黃牛各3頭,迅速采集其心臟組織,使用Trizol法提取RNA,瓊脂糖凝膠電泳切膠選取18—30nt的片段,連接3'端和5'端,反轉(zhuǎn)錄后進(jìn)行擴(kuò)增,凝膠電泳切膠純化后建立藏黃牛和黃牛各3個(gè)文庫,利用Illumina HiSeq4000測序平臺進(jìn)行高通量測序;將測序得到的序列進(jìn)行過濾,比對GenBank和Rfam數(shù)據(jù)庫,篩選出藏黃牛和宣漢黃牛的差異miRNA,進(jìn)行功能注釋及信號通路富集分析;隨機(jī)選擇8個(gè)miRNAs,利用實(shí)時(shí)熒光定量PCR檢測其在心臟組織的表達(dá)量,以驗(yàn)證測序數(shù)據(jù)的準(zhǔn)確性。【結(jié)果】藏黃牛和宣漢黃牛心臟組織的高質(zhì)量讀值的序列分別為17 463 446條和13 662 812條,干凈讀值為16 552 296條和12 055 304條,且在藏黃牛和宣漢黃牛中高質(zhì)量核酸序列長度富集最多的均是21nt,分別為37.5%和32.1%;且共篩選出219個(gè)差異miRNAs,其中48個(gè)顯著上調(diào),171個(gè)顯著下調(diào);GO功能注釋得到差異miRNA靶基因分子功能中顯著富集的條目有22條,如,GO:0005488(結(jié)合)、GO:0005515(蛋白質(zhì)結(jié)合)和GO:0043167(離子結(jié)合);細(xì)胞組分中顯著富集的條目有20條,如,GO:0005623(細(xì)胞)、GO:0044464(細(xì)胞組分)和GO:0005622(細(xì)胞內(nèi));生物過程中顯著富集的條目有13條,如,GO:0035556(細(xì)胞內(nèi)信號轉(zhuǎn)導(dǎo))、GO:0032774(RNA生物合成過程)和GO:0006351(轉(zhuǎn)錄,DNA模板化),KEGG信號通路分析得到差異表達(dá)miRNAs靶基因顯著富集到胰島素信號通路(139個(gè)靶基因)、mTOR信號通路(38個(gè)靶基因)和HIF-1 信號通路(92個(gè)靶基因)等232個(gè)信號通路中,其中有12個(gè)靶基因共同作用于這3個(gè)信號通路,說明miRNAs可能通過這3個(gè)信號通路,共同參與藏黃牛低氧適應(yīng)性的調(diào)控;隨機(jī)選取8個(gè)miRNAs進(jìn)行熒光定量驗(yàn)證,其表達(dá)趨勢與測序結(jié)果表達(dá)趨勢基本一致,表明高通量測序數(shù)據(jù)可信度較高。【結(jié)論】得到了miRNA在藏黃牛與宣漢黃牛心臟組織中的表達(dá)譜,為揭示藏黃牛低氧適應(yīng)性的調(diào)控機(jī)制研究奠定了基礎(chǔ)。
藏黃牛;宣漢黃牛;miRNA;心臟;高通量測序;低氧適應(yīng)性
0 引言
【研究意義】由于海拔升高導(dǎo)致分壓降低,當(dāng)升高的代謝需求超過氧氣供應(yīng)時(shí),心血管和呼吸系統(tǒng)的容量受到抑制,所以缺氧是高海拔環(huán)境下的主要環(huán)境脅迫類型,會導(dǎo)致代謝需求與氧氣對流運(yùn)輸之間的不平衡,最終可引起全身性低氧血癥;氧氣供應(yīng)不足影響動物的生理生化功能,從而抑制其生長[1-2]。宣漢黃牛廣泛分布于我國四川宣漢、通江等平原地區(qū),是我國優(yōu)質(zhì)的地方品種,但由于長期生活在平原,對于高原環(huán)境沒有良好的適應(yīng)機(jī)制。藏黃牛是以產(chǎn)乳為主, 乳、肉、役兼用的小型地方原始品種,由于近親繁殖,造成了其個(gè)體小、成熟晚、生產(chǎn)性能低等缺陷,但其能適應(yīng)低氧和低溫的高海拔惡劣環(huán)境。分布在高海拔的藏黃牛與平原地區(qū)的宣漢黃牛在適應(yīng)低氧環(huán)境機(jī)制上形成鮮明對比,本研究擬對藏黃牛和宣漢黃牛進(jìn)行轉(zhuǎn)錄組比較分析,探索藏黃牛低氧適應(yīng)的分子機(jī)制。【前人研究進(jìn)展】微小核糖核酸(microRNA,miRNA)是由 18—22個(gè)核苷酸堿基組成的非編碼內(nèi)源性小分子 RNA,由DNA轉(zhuǎn)錄產(chǎn)生,廣泛存在于動物、植物、微生物等生物體內(nèi)[3]。研究顯示,在缺氧條件下, miRNA可以與低氧轉(zhuǎn)錄因子相互作用,參與生物體內(nèi)的細(xì)胞代謝、細(xì)胞增殖、細(xì)胞分化、免疫應(yīng)答、細(xì)胞凋亡和癌癥的發(fā)生發(fā)展等一系列生物學(xué)過程[4-10]。miRNA通過指導(dǎo)RNA誘導(dǎo)沉默復(fù)合物(RISC)對mRNA進(jìn)行切割或者阻斷,影響 mRNA 的表達(dá),從而調(diào)控其蛋白表達(dá)水平[11-12]。一個(gè)miRNA可以調(diào)控多個(gè)靶基因,同時(shí)也存在多個(gè)miRNA調(diào)節(jié)同一個(gè)靶基因,研究表明,人體約1/3的基因受到miRNA的調(diào)控[13-14]。同時(shí),低氧能調(diào)節(jié)miRNA活性并控制轉(zhuǎn)錄過程中關(guān)鍵蛋白的活性[15]。在低氧條件下,miR-143和miR-101可以直接定位在組織細(xì)胞中的酵解酶己糖激酶,從而保護(hù)細(xì)胞免受缺氧性損傷[16-17]。ZHANG等[18]發(fā)現(xiàn)在魚類缺氧時(shí),酵解酶和己糖激酶顯著上調(diào),其調(diào)節(jié)的miRNA顯著下調(diào),表明這些酶調(diào)節(jié)的miRNA在應(yīng)對缺氧時(shí)有重要作用。【本研究切入點(diǎn)】近年來有大量關(guān)于高原動物適應(yīng)低氧的相關(guān)研究,但從miRNA角度研究藏黃牛適應(yīng)低氧的研究鮮有報(bào)道。【擬解決的關(guān)鍵問題】本研究首次對藏黃牛和宣漢黃牛進(jìn)行miRNA轉(zhuǎn)錄組比較分析,篩選低氧適應(yīng)性相關(guān)miRNA及其靶信號通路,為進(jìn)一步研究藏黃牛低氧適應(yīng)機(jī)制提供了基礎(chǔ)數(shù)據(jù)。
1 材料與方法
1.1 材料
2018年4月,于西藏自治區(qū)昌都市類烏齊縣和成都市郫縣各選取3頭健康的藏黃牛和宣漢黃牛,采集心臟組織,DEPC沖洗,錫箔紙包裝迅速置于液氮保存,帶回青藏高原動物遺傳資源保護(hù)與利用四川省、教育部重點(diǎn)實(shí)驗(yàn)室進(jìn)行試驗(yàn)。
1.2 總RNA提取
Trizol法提取總RNA,使用Nanodrop ND-1000分光光度計(jì)和Bioanalyzer 2100生物分析儀(Agilent公司,美國)對RNA濃度及RNA完整性進(jìn)行鑒定。
1.3 小RNA文庫構(gòu)建及Solexa測序
構(gòu)建6個(gè)測序文庫(藏黃牛3個(gè),宣漢黃牛3個(gè)):從樣品中提取總RNA后,瓊脂糖凝膠電泳切膠選取18—30nt的片段,連接3' 端和5' 端,對已連接兩端的small RNA進(jìn)行反轉(zhuǎn)錄和PCR擴(kuò)增,瓊脂糖凝膠電泳回收純化約140 bp的條帶后構(gòu)建藏黃牛和宣漢黃牛各3個(gè)文庫,構(gòu)建好的文庫使用 Agilent 2100以及qPCR進(jìn)行質(zhì)控,最后利用測序平臺Illumina HiSeq4000進(jìn)行測序。
1.4 數(shù)據(jù)處理統(tǒng)計(jì)
將測序得到的原始序列過濾掉低質(zhì)量、有5'端和沒有3'端、不含插入片段和插入片段小于18nt、含有polyA的序列,得到的有效序列進(jìn)行總數(shù)、種類和長度分布統(tǒng)計(jì),并做后續(xù)分析。
1.5 基因?qū)Ρ确治?/h3>
1.5.1 比對GenBank和Rfam數(shù)據(jù)庫 運(yùn)用blastn軟件,盡可能去除樣本中的rRNA、scRNA、snoRNA、snRNA和tRNA。
1.5.2 基因組對比 通過Bowtie 1.1.2軟件確定測序得到的sRNA序列在基因組上的位置,根據(jù)外顯子和內(nèi)含子在基因組的位置,找出來自mRNA降解片段的sRNA,通過 RepeatMasker version open-4.0.6軟件將sRNA與重復(fù)序列進(jìn)行比對,得到repeat associate sRNA。
1.5.3 miRNA鑒定 利用Bowtie(version 1.1.2)軟件與miRBase數(shù)據(jù)庫中所有物種的miRNA 序列進(jìn)行比對分析,鑒定出已知保守的miRNA 及其含量,利用MIREAP_v0.2 軟件預(yù)測miRNA。
1.6 差異表達(dá)分析
利用edgeR 軟件,使用edgeR默認(rèn)參數(shù),對已存在miRNA、已知miRNA和預(yù)測的新miRNA進(jìn)行差異表達(dá)分析。差異表達(dá)miRNA的篩選標(biāo)準(zhǔn)為表達(dá)量變化2倍以上并且<0.05。
1.7 靶基因預(yù)測及靶基因富集分析
使用RNAhybrid(v2.1.2)+svm_light(v6.01),Miranda(v3.3a),TargetScan(Version: 7.0)3種方法進(jìn)行靶基因預(yù)測,然后取3種方法得到的靶基因預(yù)測結(jié)果的交集作為miRNA靶基因預(yù)測的結(jié)果。利用R語言中clusterProfilter軟件包和org.Hs.eg.db數(shù)據(jù)庫,根據(jù)靶基因注釋信息進(jìn)行GO富集和KEGG通路分析。
1.8 RT-qPCR驗(yàn)證
根據(jù)反轉(zhuǎn)錄試劑盒(TaKaRa公司)說明書,對總RNA進(jìn)行反轉(zhuǎn)錄合成cDNA,標(biāo)記后于-20℃保存。
隨機(jī)選取8個(gè)miRNAs,用U6[19](F: GCTTCGG CAGCACATATACTAAAAT;R: CGCTTCACGAATT TGCGTGTCAT)作為內(nèi)參基因,每個(gè)樣品設(shè)置 3個(gè)重復(fù)。根據(jù)這8個(gè)miRNAs的成熟序列設(shè)計(jì)反轉(zhuǎn)錄及熒光定量PCR引物,引物序列見表1。利用Prime ScriptTMRT反轉(zhuǎn)錄試劑盒和TB Green Premix Ex TaqTMⅡ定量試劑盒,對目標(biāo)miRNA在藏黃牛和宣漢黃牛心臟組織中的表達(dá)水平進(jìn)行驗(yàn)證。
反應(yīng)體系10 μL:5 μL SYBR premix Dimer Eraser(2×),上下引物各0.4 μL,無菌去離子水3.2 μL,cDNA1.0 μL。
定量程序:95℃預(yù)變性30 s,95℃變性5 s,60℃退火30 s,72℃延伸30 s,39個(gè)循環(huán)。
采用2-ΔΔCT法計(jì)算目的基因表達(dá)量,結(jié)果以“平均值±標(biāo)準(zhǔn)誤差(Mean±SEM)”表示。
2 結(jié)果
2.1 數(shù)據(jù)處理統(tǒng)計(jì)
本次試驗(yàn)構(gòu)建的6個(gè)文庫,共獲得95 223 513條原始讀值,經(jīng)過一系列過濾后得到藏黃牛和宣漢黃牛的高質(zhì)量讀值的序列分別為17 463 446條和13 662 812條,干凈讀值的序列分別為16 552 296條和12 055 304條(表2)。
通過高通量測序分析得到藏黃牛和宣漢黃牛高質(zhì)量核酸序列長度均主要分布在21nt,分別為37.5%和32.1%,具體見圖1。

表1 熒光定量引物
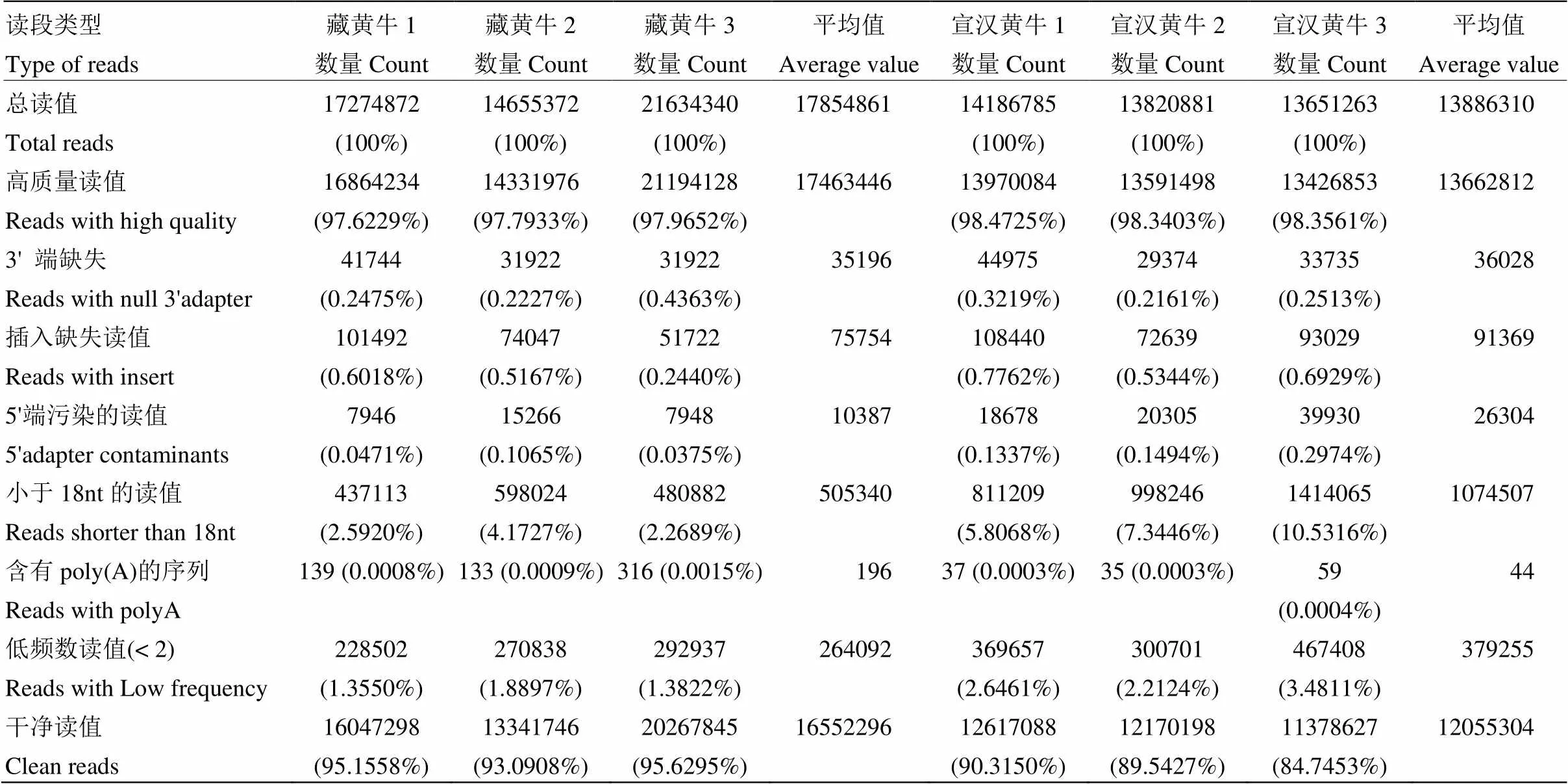
表2 數(shù)據(jù)過濾及去端頭情況統(tǒng)計(jì)
2.2 miRNA 表達(dá)分析
差異表達(dá)分析共篩選出219個(gè)差異表達(dá)miRNAs,其中48個(gè)顯著上調(diào),171個(gè)顯著下調(diào)(圖2)。表3為差異表達(dá)中表達(dá)量較高的10個(gè)miRNAs。
2.3 miRNA靶基因預(yù)測及靶基因富集分析
2.3.1 miRNA 靶基因預(yù)測 由表4可知,藏黃牛靶基因預(yù)測數(shù)量為58 111個(gè),宣漢黃牛為58 541個(gè),對應(yīng)的靶基因位點(diǎn)數(shù)量為2 584 253個(gè)和3 017 725個(gè)。
2.3.2 miRNA 靶基因富集分析 對差異表達(dá)miRNA的靶基因進(jìn)行GO功能注釋結(jié)果表明:miRNA靶基因顯著富集55條GO條目,其中生物過程22條,細(xì)胞組分20條,分子功能13條。由圖3可知,在生物過程中,細(xì)胞過程和單一生物過程富集水平最高,且顯著富集前三的是GO:0035556(細(xì)胞內(nèi)信號轉(zhuǎn)導(dǎo))、GO:0032774(RNA生物合成過程)和GO:0006351(轉(zhuǎn)錄,DNA模板化);在分子功能中,結(jié)合和催化活性富集水平最高,且富集顯著前三的是GO: 0005488(結(jié)合)、GO:0005515(蛋白質(zhì)結(jié)合)和GO:0043167(離子結(jié)合);在細(xì)胞組分中,細(xì)胞和細(xì)胞部分富集水平最高,且富集顯著前三的是GO: 0005623(細(xì)胞)、GO:0044464(細(xì)胞組分)和GO: 0005622(細(xì)胞內(nèi))。
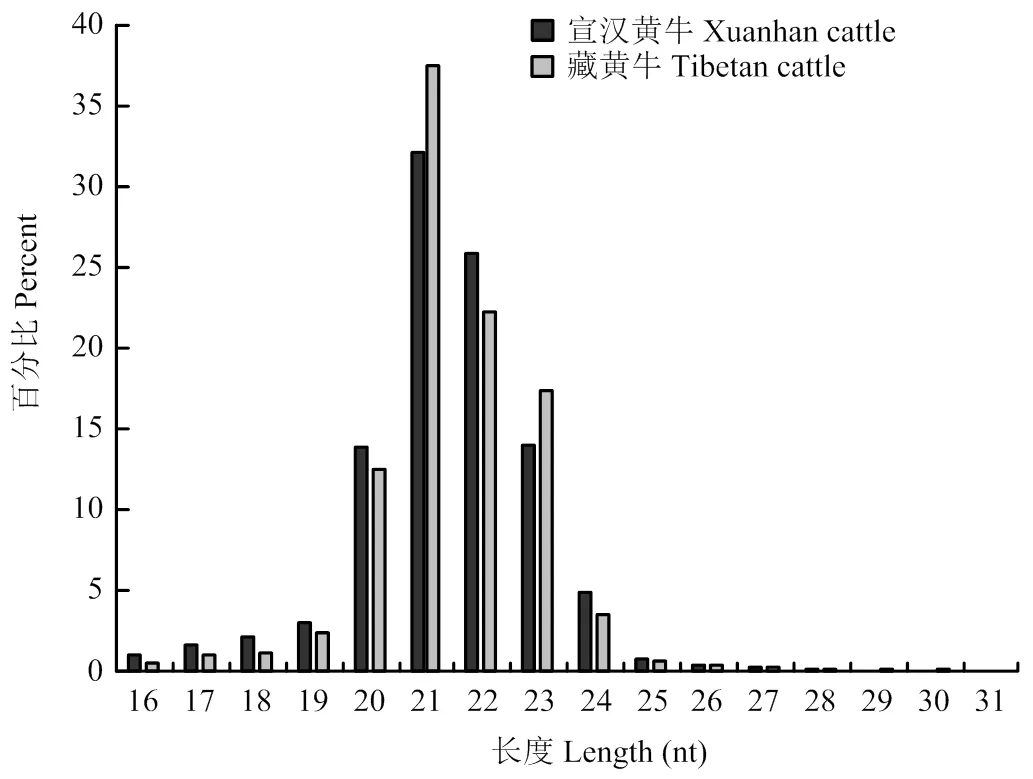
圖1 藏黃牛和宣漢黃牛miRNA的長度分布及頻率百分比
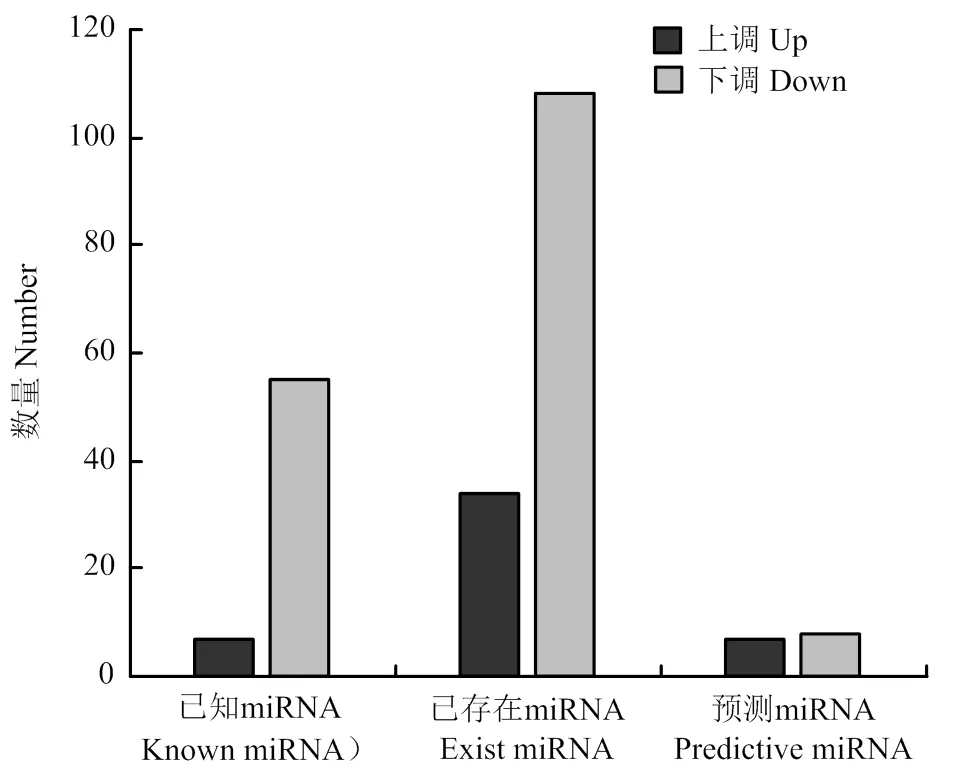
圖2 藏黃牛和宣漢黃牛miRNA 差異表達(dá)統(tǒng)計(jì)圖
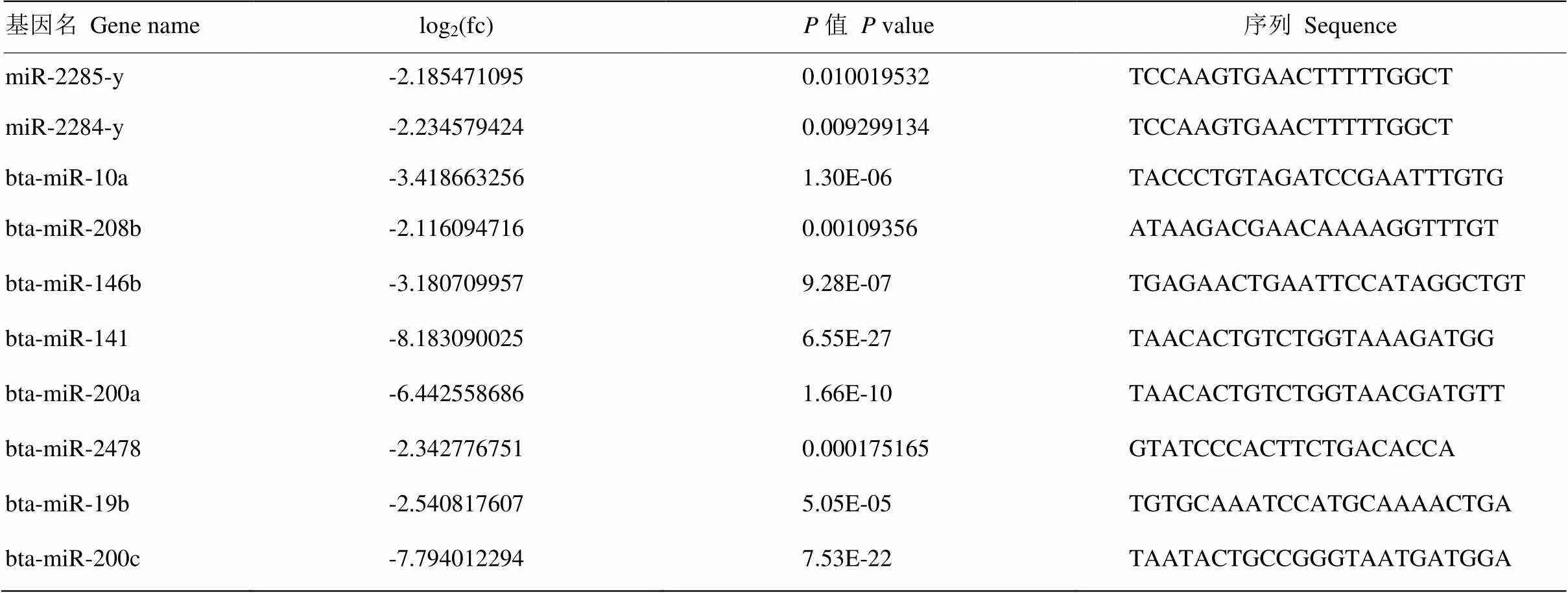
表3 差異表達(dá)中表達(dá)量較高的10個(gè) miRNAs
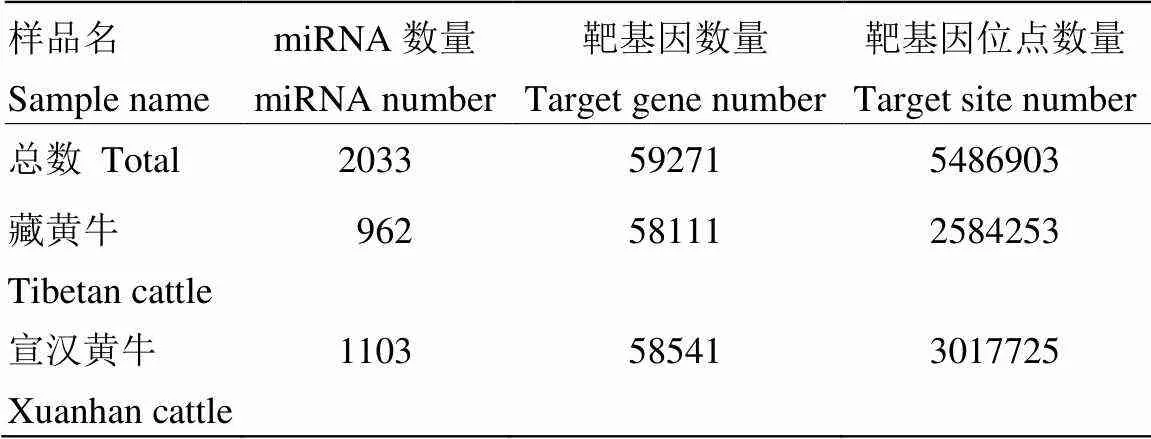
表4 藏黃牛和宣漢黃牛miRNA靶基因位點(diǎn)預(yù)測統(tǒng)計(jì)
KEGG Pathway富集分析結(jié)果表明,差異表達(dá)miRNAs靶基因顯著富集到MAPK信號通路、mTOR信號通路、軸突導(dǎo)向和胰島素信號通路等信號通路,圖4為其中20條信號通路;在低氧適應(yīng)相關(guān)通路中發(fā)現(xiàn)胰島素信號通路涉及差異表達(dá)miRNAs的靶基因135個(gè),mTOR信號通路涉及差異表達(dá)miRNAs的靶基因38個(gè),HIF-1信號通路涉及差異表達(dá)miRNAs的靶基因65個(gè),且有12個(gè)靶基因(、、、、、、、、、、、)共同參與胰島素信號通路、HIF-1信號通路和mTOR信號通路(表5)。
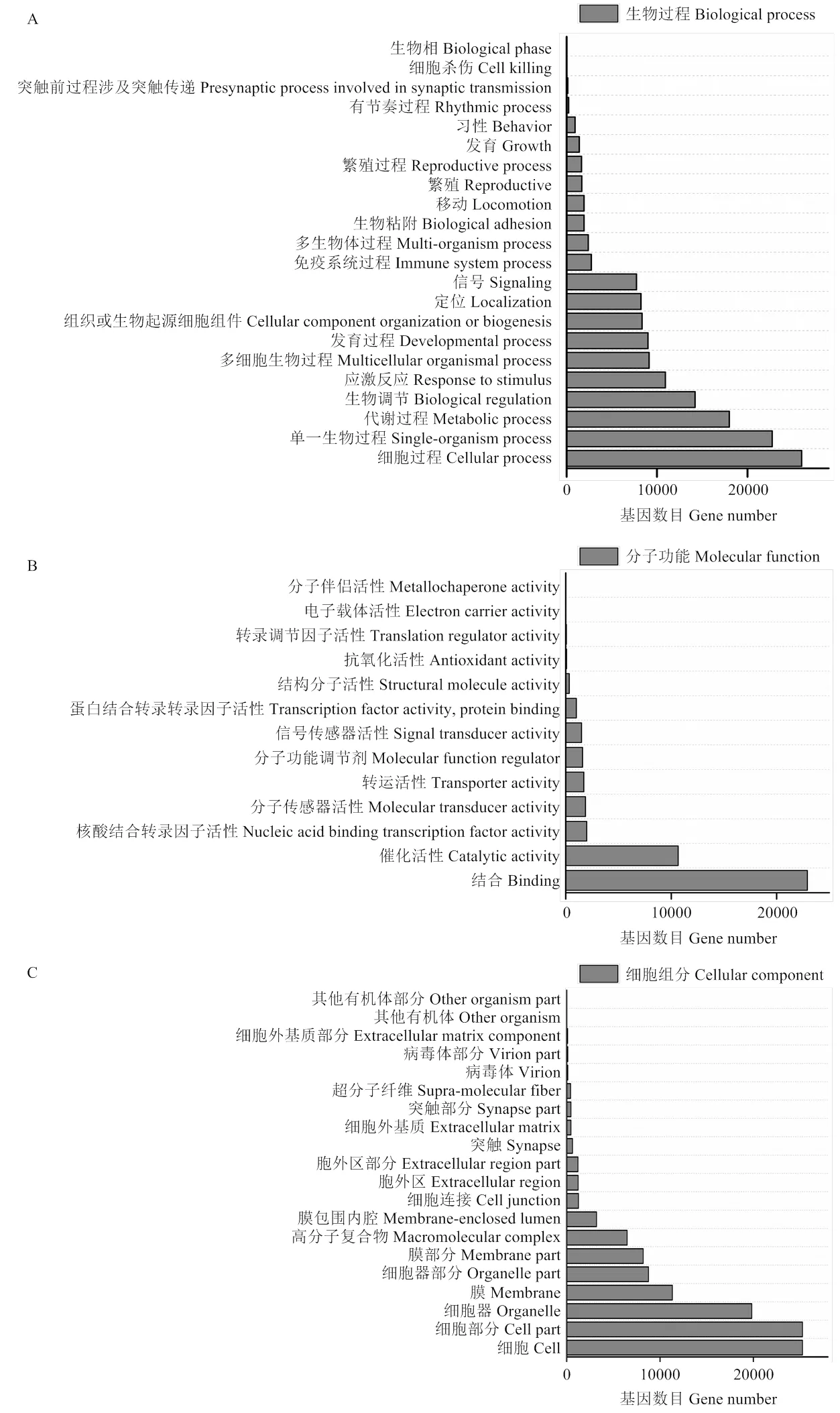
圖3 藏黃牛和宣漢黃牛miRNA靶基因GO 富集分類柱狀圖(A:生物過程 B:分子功能 C:細(xì)胞組分)
2.4 RT-qPCR驗(yàn)證
由圖5可知,RT-qPCR結(jié)果表明,miRNA在宣漢黃牛和藏黃牛心臟組織中的表達(dá)趨勢與高通量測序所得表達(dá)趨勢基本一致,說明測序得到數(shù)據(jù)的可靠性高;且在RT-qPCR 驗(yàn)證中發(fā)現(xiàn)bta-miR-99a-5p在藏黃牛心臟組織中的表達(dá)量顯著高于宣漢黃牛心臟組織中的表達(dá)量(<0.05),其他基因在藏黃牛和宣漢黃牛心臟組織中的表達(dá)則不具有顯著性差異(>0.05)。
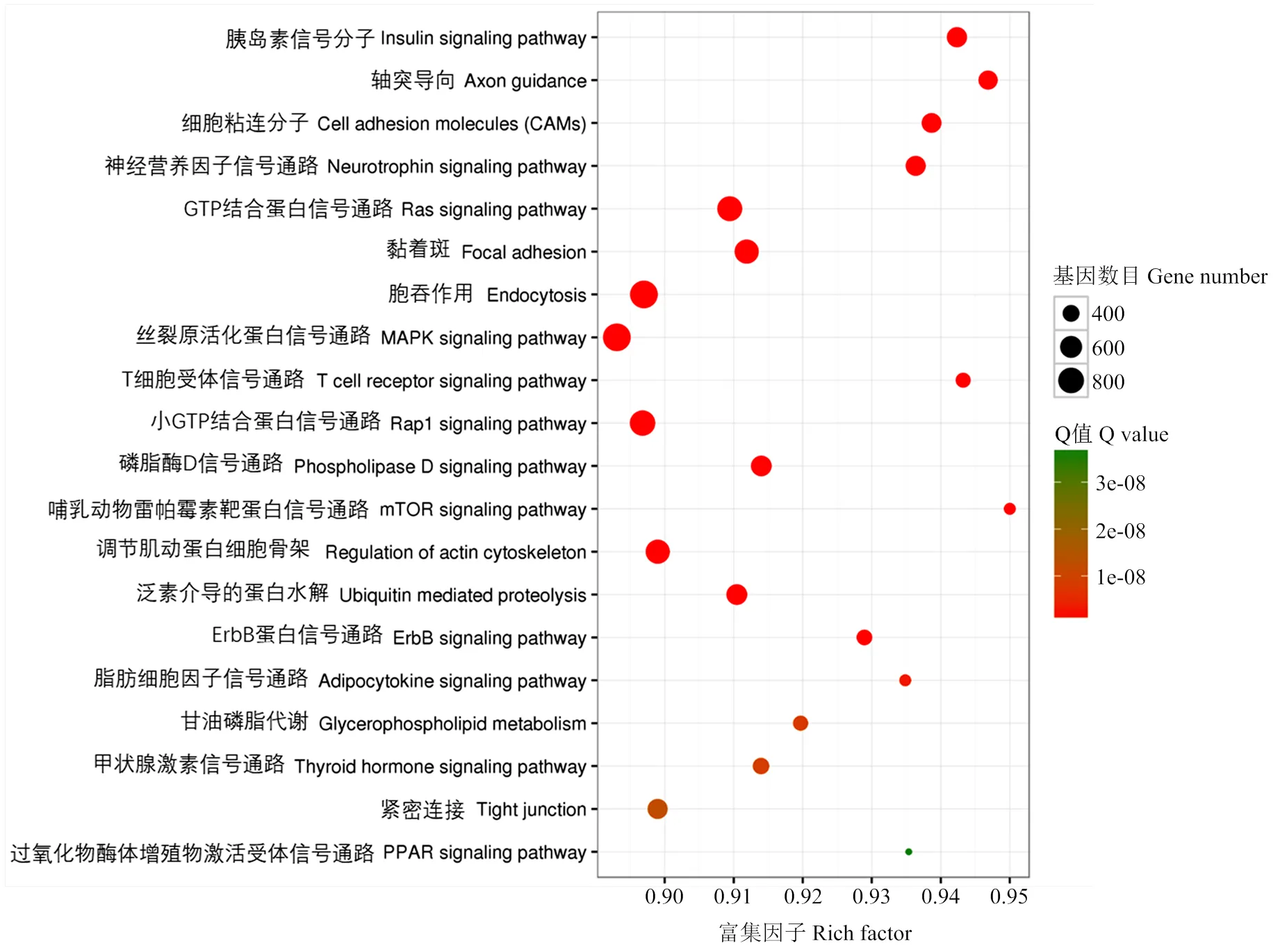
圖4 藏黃牛和宣漢黃牛miRNA靶基因Pathway 通路分析

表5 3個(gè)信號通路涉及的靶基因
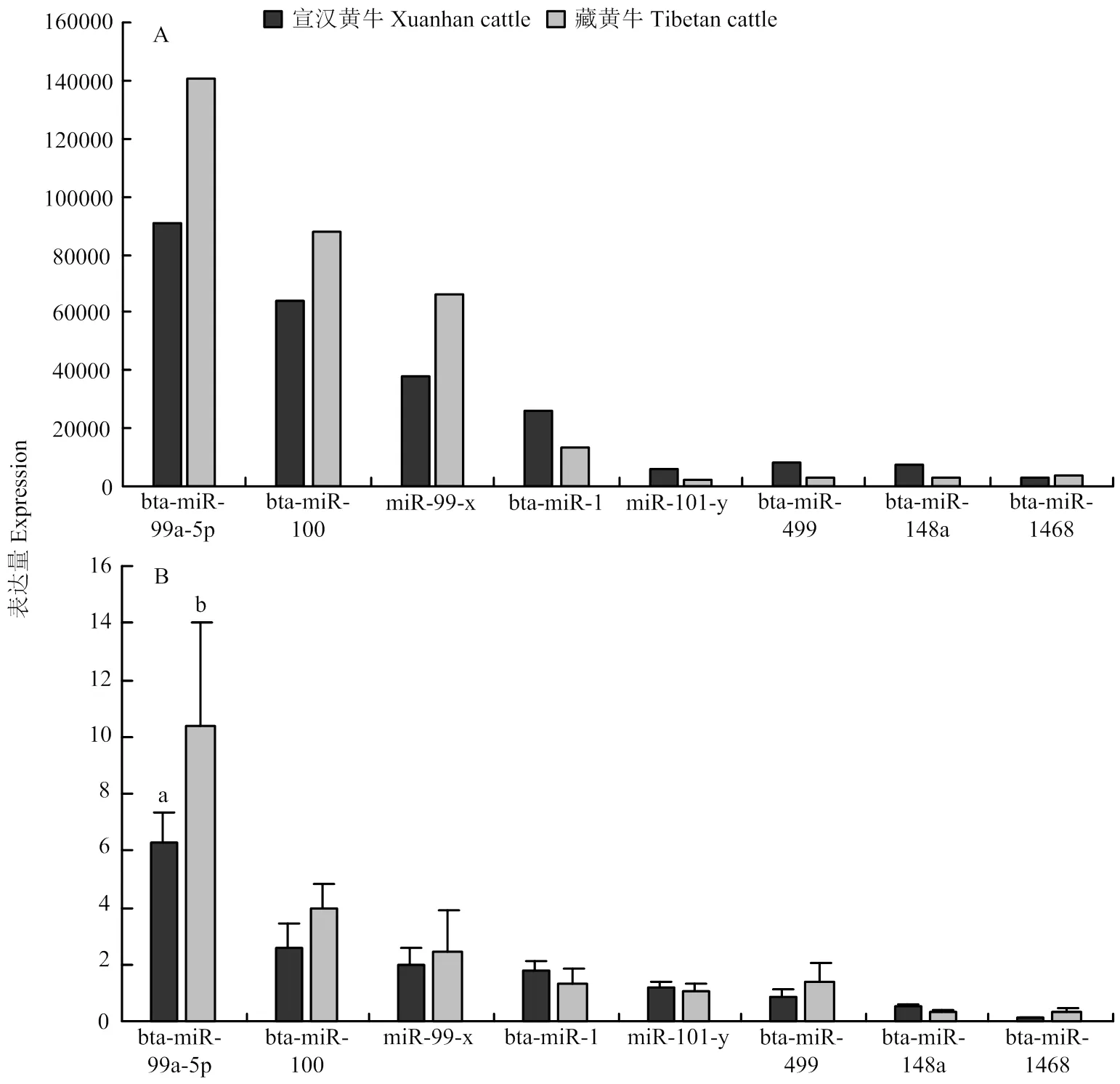
不同字母表示具有顯著性差異(P<0.05) Different letters indicate significant difference (P<0.05)
3 討論
隨著科學(xué)技術(shù)的飛速發(fā)展,高通量測序因?yàn)椴僮骱唵巍⑿矢摺⑼看蟮葍?yōu)點(diǎn)在科學(xué)實(shí)驗(yàn)中得到了廣泛應(yīng)用;它能夠快速地獲得成千上萬條基因序列,以此開發(fā)出了大量的微衛(wèi)星分子標(biāo)記資源[20]。
心臟有左心房、左心室、右心房和右心室4個(gè)腔室,是脊椎動物最重要的器官之一,主要作用是推動血液流動,為組織和器官提供充足的血流量,是機(jī)體各項(xiàng)生命活動的重要?jiǎng)恿碓础2攸S牛長期生活在高海拔地區(qū),為了能夠適應(yīng)惡劣環(huán)境,對低氧、低溫逐漸形成了適應(yīng)性特征,如肺臟重為其體重的1.1%—1.7%,心臟的重量占其體重的0.5%—0.8%[21]。
miRNA是參與轉(zhuǎn)錄后表達(dá)水平調(diào)節(jié)的重要因子,是表型調(diào)控的重要潛在位點(diǎn),而缺氧會影響某些特定組miRNA的表達(dá)[22-23],本研究通過高通量測序分析,去除質(zhì)量值低于20的堿基數(shù)超過1個(gè)或含有N的片段,以及rRNA,scRNA,snoRNA,snRNA,tRNA,得到高質(zhì)量片段,結(jié)果分析表明藏黃牛和宣漢黃牛miRNA的長度均集中分布在21 nt,符合典型的Dicer酶切割成熟的miRNA的長度分布[24]。在RT-qPCR驗(yàn)證試驗(yàn)中,bta-miR-499的表達(dá)趨勢與測序數(shù)據(jù)不一樣,可能是由于樣本量較少和試驗(yàn)方法不同而造成的,但其他miRNA的表達(dá)趨勢和測序結(jié)果中miRNA的表達(dá)趨勢基本相同,表明本研究所得數(shù)據(jù)有較高的可信度。研究顯示,缺氧誘導(dǎo)因子(HIF)可作為氧穩(wěn)態(tài)的主要調(diào)節(jié)劑,能夠刺激紅細(xì)胞生成;脯氨酰羥化酶蛋白(EGLN1)錯(cuò)義突變編碼的脯氨酰羥化酶2(PHD2)是HIF依賴的負(fù)調(diào)節(jié)劑,在適應(yīng)低氧的情況下可防止紅細(xì)胞增多癥;細(xì)胞過度磷酸化缺陷型的過度表達(dá)能夠干擾MAPK依賴性,從而調(diào)節(jié)HIF的活性[25-26]。本試驗(yàn)的差異表達(dá)分析及生物信息分析結(jié)果表明,bta-miR-499y的靶基因和,miR-101-y的靶基因、bta- miR-101的靶基因,bta-miR-2284p的靶基因均屬于低氧適應(yīng)性基因。推測這些miRNAs可能通過這些低氧適應(yīng)性相關(guān)基因從而參與低氧適應(yīng)性的調(diào)控。
在差異表達(dá)分析中,我們共挖掘了219個(gè)差異表達(dá)miRNAs(48個(gè)顯著上調(diào),171個(gè)顯著下調(diào)),說明miRNA主要通過負(fù)調(diào)控基因的表達(dá)從而參與到機(jī)體生長發(fā)育的過程中。但也有相關(guān)研究表明miRNA的作用機(jī)制也會因細(xì)胞所處的發(fā)育階段或特殊的生理?xiàng)l件而存在差異[27]。有研究發(fā)現(xiàn),miRNA在牦牛和黃牛心臟組織中的差異表達(dá)基因富集到胰島素信號通路、mTOR信號通路和HIF-1信號通路等與低氧適應(yīng)性調(diào)控密切相關(guān)的信號通路中;藏雞適應(yīng)高原環(huán)境也是通過HIF-1信號通路、黑色素生成、DNA修復(fù)等通路實(shí)現(xiàn)[28-29]。這與本研究得到的KEGG信號通路分析結(jié)果基本一致,說明miRNA靶向的差異表達(dá)基因具有較高的保守性。本試驗(yàn)差異表達(dá)miRNAs靶基因功能富集分析表明,富集最多的GO條目涉及細(xì)胞過程和代謝過程,推測差異表達(dá)miRNAs參與心肌細(xì)胞增殖及能量代謝等生理過程。
mTOR信號通路屬于磷脂酰肌醇-3激酶,其通過MTOR C1(MTOR complex1, MTOR C1)和MTOR C2(MTOR complex2,MTOR C2)實(shí)現(xiàn)對細(xì)胞生長、增殖等生理功能的調(diào)控[30]。MTOR C1對心臟有保護(hù)作用;而MTOR C2能夠保護(hù)心臟結(jié)構(gòu)和功能[31-32]。前人研究表明激活PI3K-AKT-mTOR信號通路能促進(jìn)細(xì)胞生長[33],它誘導(dǎo)激活的miR-199a[34]和miR-21[35]能通過靶向GLUT1(胰島素依賴性葡萄糖轉(zhuǎn)運(yùn)蛋白)調(diào)節(jié)葡萄糖代謝。HIF依賴性代謝適應(yīng)有助于活細(xì)胞保持完整性,維持能量供應(yīng),并在低氧條件下維持其功能;缺氧情況下,能夠啟動缺氧誘導(dǎo)因子-1a()的表達(dá),并進(jìn)一步誘導(dǎo)(血管內(nèi)皮生長因子)和(絲裂原活化蛋白激酶)等靶基因的表達(dá),促進(jìn)血管生成及發(fā)育、呼吸系統(tǒng)發(fā)育和心肌細(xì)胞發(fā)育等細(xì)胞組織生理變化[36-39]。有研究發(fā)現(xiàn),miR-20a能通過靶向HIF-la來調(diào)節(jié)VEGF的表達(dá)[40]。bta-miR-101主要富集到胰島素信號通路和mTOR信號通路,推測bta-miR-101能夠促進(jìn)細(xì)胞生長;bta-miR-499和bta-miR-126-3p等miRNA同時(shí)作用于胰島素信號通路、mTOR信號通路和HIF-1信號通路,由此推測,bta-miR-499和bta-miR-126-3p等miRNA可能通過HIF-1信號通路增加氧氣的運(yùn)輸量,為機(jī)體提供更多氧氣以適應(yīng)高海拔環(huán)境。同時(shí),也有研究發(fā)現(xiàn),miR-101可以在缺氧情況下減少心肌細(xì)胞凋亡[41];miR-499能夠抑制心肌梗死后心肌纖維化并且改善心臟推動血液循環(huán)的功能[42];miR-126-3p促進(jìn)表達(dá)[43]。本研究結(jié)果表明,有12個(gè)靶基因(、、、、、、、、、、、)共同參與胰島素信號通路、HIF-1信號通路和mTOR信號通路,推測差異表達(dá)miRNAs可能通過這些靶基因參與藏黃牛的低氧適應(yīng)性機(jī)制的調(diào)控。
4 結(jié)論
本研究通過對藏黃牛和宣漢黃牛miRNA高通量測序分析,篩選出了219個(gè)差異表達(dá)miRNAs,其靶基因富集到胰島素信號通路、mTOR信號通路和HIF-1信號通路等對心臟的結(jié)構(gòu)保護(hù)、能量產(chǎn)生及儲存具有重要作用的通路中,推測這些差異表達(dá)基因在藏黃牛適應(yīng)低氧環(huán)境的過程中具有重要調(diào)控作用。本研究為進(jìn)一步探討高原動物低氧適應(yīng)機(jī)制提供了基礎(chǔ)數(shù)據(jù)。
[1] TEKIN D, DURSUN A D, XI L. Hypoxia inducible factor 1 (HIF-1) and cardioprotection., 2010, 31(9): 1085-1094.
[2] SEMENZA G L. Hypoxia-inducible factors in physiology and medicine., 2012,148(3):399-408.
[3] BARTEL D P. MicroRNAs: genomics, biogenesis, mechanism, and function., 2004, 116(2):281-297.
[4] HE M, LU Y, XU S, MAO L, ZHANG L, DUAN W, LIU C, PI H, ZHANG Y, ZHONG M, YU Z, ZHOU Z. MiRNA-210 modulates a nickel-induced cellular energy metabolism shift by repressing the iron-sulfur cluster assembly proteins ISCU1/2 in Neuro-2a cells., 2014, 5(2):e1090.
[5] CONTI A, ROMEO S G, CAMA A, LATORRE D, BARRESI V, PEZZINO G, TOMASELLO C, CARDALI S, ANGILERI F F, POLITO F, FERLAZZO G, DIGIORGIO R, GERMANO A, AGUENNOUZ M. MiRNA expression profiling in human gliomas: upregulated miR-363 increases cell survival and proliferation., 2016, 37(10):14035-14048.
[6] PANDEY R K, SUNDAR S, PRAJAPAATI V K. Differential expression of miRNA regulates T Cell differentiation and plasticity during visceral leishmaniasis infection., 2016, 7(25):206-215.
[7] FULLAONDO A, LEE S Y. Identification of putative miRNA involved in Drosophila melanogaster immune response., 2012, 36(2):267-273.
[8] FU X, MENG Z, LIANG W, TIAN Y, WANG X, HAN W, LOU G, WANG X, LOU F, YEN Y, YU H, JOVE R, HUANG, W. MiR-26a enhances miRNA biogenesis by targeting Lin28B and Zcchc11 to suppress tumor growth and metastasis., 2014, 33(34): 4296-4306.
[9] CHENG Y, XIANG G, MENG Y, DONG R. MiRNA-183-5p promotes cell proliferation and inhibits apoptosis in human breast cancer by targeting the PDCD4., 2016, 16(3): 225-233.
[10] Hessam S, Sand M, Skrygan M, GAMBICHLER T, BECHARA F G. Expression of miRNA-155, miRNA-223, miRNA-31, miRNA-21, miRNA-125b, and miRNA-146a in the Inflammatory Pathway of hidradenitis suppurativa., 2017, 40(2): 464-472.
[11] MAL C, AFTABUDDIN M, KUNDU S. IIKmTA: Inter and intra kingdom miRNA-target analyzer., 2018, 10(3):538-543.
[12] JONAS S, LZAURRALDE E. Towards a molecular understanding of microRNA-mediated gene silencing., 2015, 16(7):421-433.
[13] FENG B, CHAKRABARTI S. MiR-320 regulates glucose-induced gene expression in diabetes., 2012, 6(7):1-7.
[14] CIMMINO A, CALIN GA, FABBRI M, LORIO M V, FERRACIN M, SHIMIZU M, WOJCIK S E, AQEILAN R I, ZUPO S, DONO M, RASSENTI L, ALDER H, VOLINIA S, LIU C G, KIPPS T J, NEGRINI M, CROCE C M. MiR-15 and miR-16 induce apoptosis by targeting BCL2., 2005, 102(39):13944-13949.
[15] NALLAMSHETTY S, CHAN S Y, LOSCALZO J. Hypoxia: a master regulator of microRNA biogenesis and activity., 2013, 64:20-30.
[16] YAO M, WANG X, TANG Y, ZHANG W, Cui B, Liu Q, Xing L. Dicer mediating the expression of miR-143 and miR-155 regulates hexokinase II associated cellular response to hypoxia., 2014, 307(11): 829-837.
[17] XU X, LIU C, BAO J. Hypoxia-induced hsa-miR-101 promotes glycolysis by targeting TIGAR mRNA in clear cell renal cell carcinoma., 2017, 15(3):1373–1378.
[18] ZHANG G, YIN S, MAO J, LIANG F, ZHAO C, LI P, ZHOU G, CHEN S, TANG Z. Integrated analysis of mRNA-seq and miRNA-seq in the liver ofin response to hypoxia., 2016, 10(6):22907.
[19] 張陽陽. miR-378在牛前體脂肪細(xì)胞分化的作用與機(jī)制[D]. 長春: 吉林大學(xué), 2014.
ZHANG Y Y. Effect and mechanism of bovine miR-378 in preadipocyte differention[D]. Changchun: Jilin University, 2014. (in Chinese)
[20] LI H J, LIU M, YE S, YANG F. De novo assembly, gene annotation, and molecular marker development using Illumina paired-end transcriptome sequencing in the clam., 2017, 39 (6): 675-685.
[21] 郭勝祥,劉永年. 高原適應(yīng)動物牦牛與普通黃牛肺血管反應(yīng)性的比較研究. 中國病理生理雜志, 1995, 11(3):230-233.
GUO S X, LIU Y N. Comparative study on pulmonary vascular reactivity of plateau adapted animal Yak and common cattle., 1995, 11(3):230-233. (in Chinese)
[22] 金澄艷, 呂曉陽, 高雯, 王悅, 陳煒昊, 盛水興, 陳玲, 林杰, 孫偉. 湖羊羔皮毛囊候選miRNA在不同花紋間的表達(dá)與毛囊發(fā)育特性關(guān)聯(lián)的研究. 中國農(nóng)業(yè)科學(xué), 2018, 51(14): 2814-2824.
JIN C Y, Lü X Y, GAO W, WANG Y, CHEN W H, SHEN S X, CHEN L, LIN J, SUN W. Study on the relationship between the expression of candidate miRNAs and the developmental characteristics in different patterns in Hu Sheep Lambskin., 2018, 51(14):2814-2824. (in Chinese)
[23] NALLAMSHETTY S, CHAN S Y, LOSCALZO J. Hypoxia: a master regulator of microRNA biogenesis and activity., 2013, 64:20-30.
[24] BURNSIDE J, OUYANG M, ANDERSON A, BERNBERG E, LU C, MEYERS B C, GREEN P J, MARKIS M, ISAACS G, HUANG E, MORGAN R W. Deep sequencing of chicken microRNAs., 2008, 9(1):185.
[25] LORENZO F R, HUFF C, MYLLYMAKI M, OLENCHOCK B, SWIERCZEK S, TASHI T, GORDEUK V, WUREN T, RI-LI G, MCCLAIN D A, KHAN T M, KOUL P A, GUCHHAIT P, SALAMA M E, XING J, SEMENZA G L, LIBERZON E, WILSON A, SIMONSON T S, JORDE L B, KAELIN W G J, KOIVUNEN P, PRCHAL J T. A genetic mechanism for Tibetan high-altitude adaptation., 2014, 46(9): 951-956.
[26] QU Y, ZHAO H, HAN N, ZHOU G, SONG G, GAO B, TIAN S, ZHANG J, ZHANG R, MENG X, ZHANG Y, ZHANG Y, ZHU X, WANG W, LAMBERT D, ERICSON P G, SUBRAMANIAN S, YEUNG C, ZHU H, JIANG Z, LI R, LEI F. Ground tit genome reveals avian adaptation to living at high altitudes in the Tibetan plateau., 2013, 4:2071.
[27] VASUDEVAN S, TONG Y, STEITZ J A. Switching from repression to activation: microRNAs can up-regulate translation., 2007, 318(5858):1931-1934.
[28] 賀大芳. 牦牛和黃牛心臟、肺臟組織microRNA轉(zhuǎn)錄組的鑒定與差異表達(dá)分析[D]. 成都:四川農(nóng)業(yè)大學(xué), 2017.
HE D F. Identification and differential expression analysis of microRNAs in heart and lung tissues between yak and cattle[D]. Chengdu: Sichuan Agricultural University, 2017. (in Chinese)
[29] ZHANG Q, GOU W, WANG X, ZHANG Y, MA J, ZHANG H, ZHANG Y, ZHANG H. Genome resequencing identifies unique adaptations of Tibetan Chickens to hypoxia and high-dose ultraviolet radiation in high-altitude environments., 2016, 8(3):765-776.
[30] 劉杰, 李景東. 哺乳動物雷帕霉素靶蛋白信號通路在心臟發(fā)育和重構(gòu)中作用的研究進(jìn)展. 心血管病學(xué)進(jìn)展, 2018(06): 911-915.
LIU J, LI J D. Research progress of mTOR signaling pathway effect in cardiac development and reconstruction., 2018(06):911-915. (in Chinese)
[31] SCIARRETTA S, ZHAI P, MAEJIMA Y, DEL R D P, NAGARAJAN N,YEE D, LIU T, MAGNUSON M A, VOLPE M, FRATI G, LI H, SADOSHIMA J. mTORC2 regulates cardiac response to stress by inhibiting MST1., 2015, 11(1):125-136.
[32] VERMA P, SHARMA A, SODHI M, THAKUR K, KATERIA R S, NIRANJAN S K, BHARTI V K, KUMAR P, GIRI A, KALIA S, MUKESH M. Transcriptome analysis of circulating pbmcs to understand mechanism of high altitude adaptation in native cattle of ladakh region., 2018, 8(1):7681.
[33] VERMA P, SHARMA A, SODHI M, THAKUR K, BHARTI V K, KUMAR P, GIRI A, KALIA S, SWAMI S K, MUKESH M.Overexpression of genes associated with hypoxia in cattle adapted to trans himalayan region of Ladakh., 2018, 42(9):1141-1148.
[34] TAGUCHI A, YANAGISAWA K, TANAKA M, CAO K, MATSUYAMA Y, GOTO H, TAKAHASHI T. Identification of hypoxia-inducible factor-1 alpha as a novel target for miR-17-92 microRNA cluster., 2008, 68(14):5540-5545.
[35] GIATROMANOLAKI A, BAI M, MARGARITIS D, BOURANTAS K L, KOUKOURAKIS M I, SIVRIDIS E, GATTER K C. Hypoxia and activated VEGF/receptor pathway in multiple myeloma., 2010, 30(7):2831-2836.
[36] BEFANI C D, VLACHOSTERRGIOS P J, HATZIDAKI E, PATRIKIDOU A, BONANOU S, SIMOS G, PAPANDREOU C N, LIAKOS P. Bortezomib represses HIF-1α protein expression and nuclear accumulation by inhibiting both PI3K/Akt/TOR and MAPK pathways in prostate cancer cells., 2012, 90(1):45-54.
[37] MAJMUNDARR A J, WONG W J, SIMON M C. Hypoxia-inducible factors and the response to hypoxic stress., 2010, 40(2):294-309.
[38] TAGUCHI A, YANAGISAWA K, TANAK A M, CAO K, MATSUYAMA Y, GOTO H, TAKAHASHI T. Identification of hypoxia-inducible factor-1 alpha as a novel target for miR-17-92 microRNA cluster., 2008, 68(14):5540-5545.
[39] CASCIO S, D'ANDREA A, FERLA R, SURMACZ E, GULOTTA E, AMODEO V, BAZAN V, GEBBIA N, RUSSO A. MiR-20b modulates VEGF expression by targeting HIF-1 alpha and STAT3 in MCF-7 breast cancer cells.2010, 224(1): 242-249.
[40] 岳瑩, 呂風(fēng)華, 陳玉磊, 王卓, 司澳洋. miR-499對缺氧/復(fù)氧誘導(dǎo)的心肌細(xì)胞凋亡的影響. 鄭州大學(xué)學(xué)報(bào)(醫(yī)學(xué)版), 2018, 53(04): 503-507.
YUE Y, Lü F H, CHEN Y L, WANG Z, SI A Y. Effect of miR-499 on apoptosis of primary cardiomyocytes induced by anoxia-reoxygenation., 2018, 53(04): 503-507. (in Chinese)
[41] 趙欣. MicroRNA-101a通過靶向調(diào)控心臟成纖維細(xì)胞TGFβRI的表達(dá)抑制缺氧誘導(dǎo)的心肌纖維化[D]. 武漢:華中科技大學(xué), 2015.
ZHAO X. MicroRNA-101a inhibits hypoxia-induced myocardial fibrosis by targeting regulation of cardiac fibroblast TGFβRI expression[D]. Wuhan:Huazhong University of Science and Technology, 2015. (in Chinese)
[42] XI T Y, JIN F, ZHU Y, WANG J, TANG L, WANG Y, LIEBESKKIND D S, HE Z.MicroRNA-126-3p attenuates blood-brain barrier disruption, cerebral edema and neuronal injury following intracerebral hemorrhage by regulating PIK3R2 and Akt., 2017, 494(1-2):144-151.
Comparative Analysis of miRNA Expression Profiles in the Hearts of Tibetan Cattle and Xuanhan Cattle
CHEN LuLu, WANG Hui, WANG JiKun, WANG JiaBo, CHAI ZhiXin, CHEN ZhiHua, ZHONG JinCheng
(Institute of Tibetan Plateau Research, Southwest Minzu University/Key Laboratory of Qinghai-Tibetan Plateau Animal Genetic Resource Reservation and Utilization, Sichuan Province and Ministry of Education, Chengdu 610041)
【Objective】As a kind of non-coding RNA, miRNA is widely involved in various life activities of the organism. This study was aimed to explore the differential expression profiles of miRNA in the heart tissues between Tibetan cattle and Xuanhan cattle, so as to provide the basic data for further study on molecular mechanism of hypoxia adaptation in Tibetan cattle. 【Method】Each three healthy Tibetan and Xuanhan cattle were randomly selected for heart tissue sampling. RNA was extracted from tissues using the Trizol method. An 18 to 30nt fragment was selected by agarose gel electrophoresis, and 3' connector and 5' liner was ligated and then the fragment was enlarged. After gel electrophoresis, three Tibetan cattle and Xuanhan cattle libraries were established, respectively. High-throughput sequencing was performed by using the Illumina HiSeq4000 sequencing platform. The sequence was then filtered and the differentially expressed miRNA of Tibetan cattle and Xuanhan cattle were screened by comparing GenBank and Rfam databases. Functional annotation and signal pathway enrichment analysis of differentially expressed miRNA in Tibetan cattle and Xuanhan cattle. Finally, in order to verify the accuracy of the sequencing data, 8 miRNAs were randomly selected and the expression level of miRNA was detected by RT-qPCR. 【Result】The results showed that Tibetan cattle and Xuanhan cattle had high-quality reads of 17 463 446 and 13 662 812, respectively, while the clean reads were 16 552 296 and 12 055 304, respectively. The highest enrichment of high-quality nucleic acid sequences in Tibetan cattle and Xuanhan cattle were 21 nt, which were 37.5% and 32.1%, respectively. A total of 219 differential expressed miRNAs (48 up-regulated and 171 down-regulated) were obtained. There were 22 terms in the GO function annotation that significantly enriched in the molecular function of differentially expressed miRNAs target genes, such as GO: 0005488 (binding), GO: 0005515 (protein binding) and GO: 0043167 (ion binding). GO: 0005623 (cell), GO: 0044464 (cell component) and GO: 0005622 (cell) were among the 20 terms, which were significantly enriched in the cellular components. While there were 13 terms, which were significantly enriched in biological processes, such as GO: 0035556 (intracellular signal transduction), GO: 0032774 (RNA biosynthesis process) and GO: 0006351 (transcription, DNA templated). Analysis of KEGG signaling pathways revealed that miRNA target genes were significantly enriched to 232 signaling pathways, including the insulin signaling pathway (139 target genes), the mTOR signaling pathway (38 target genes) and the HIF-1 signaling pathway (92 target genes). Among them, 12 miRNA target genes worked together on these three signaling pathways. These results suggested that the differentially expressed miRNAs might participate in the regulation of hypoxia adaptation in Tibetan cattle through these three signaling pathways. Eight miRNAs were randomly selected for RT-qPCR, and the expression profiles were consistent with the sequencing data, indicating that the high-throughput sequencing data was reliable. 【Conclusion】Taken together, the expression profiles of miRNAs in the heart tissues of Tibetan and Xuanhan cattle were obtained in the present study, which laid a foundation for further research on the hypoxia adaptation mechanism of Tibetan cattle.
Tibetan cattle; Xuanhan cattle; miRNA; heart; high-throughput sequencing; hypoxia adaptability
2019-04-30;
2020-02-19
國家肉牛牦牛產(chǎn)業(yè)技術(shù)體系項(xiàng)目(CARS-37)、青藏高原生態(tài)畜牧業(yè)協(xié)同創(chuàng)新中心開放基金資助(QZGYXT02)
陳露露,E-mail:1556370692@qq.com。通信作者鐘金城,E-mail:zhongjincheng518@126.com。通信作者陳智華,E-mail:czh@swun.cn
(責(zé)任編輯 林鑒非)
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價(jià)·高一版(2020年6期)2020-11-02 02:45:24
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國生殖健康(2019年3期)2019-02-01 06:12:26
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
鑿巖機(jī)械氣動工具(2016年3期)2016-03-01 04:00:25
海軍航空大學(xué)學(xué)報(bào)(2015年3期)2015-11-11 17:20:00
鄭州大學(xué)學(xué)報(bào)(醫(yī)學(xué)版)(2015年2期)2015-02-27 14:50:46
