桂西北喀斯特山區非共生固氮速率隨退耕后生態演替變化特征
2020-08-13 10:24:16孫錫斌張青山肖孔操李德軍
農業現代化研究 2020年4期
關鍵詞:生長
孫錫斌 ,張青山 ,肖孔操 ,李德軍
(1. 中國科學院亞熱帶農業生態研究所亞熱帶農業生態過程重點實驗室,湖南 長沙 410125;2. 中國科學院環江喀斯特生態系統觀測研究站,廣西 環江 547100;3. 中國科學院大學,北京 100049)
氮是大多數陸地自然生態系統的主要限制性營養元素,尤其是在植被恢復的早期階段[1-2]。生物固氮是陸地自然生態系統外源氮輸入的主要途徑[3-4],在生態系統演替過程中具有重要作用。生物固氮分為共生和非共生固氮,但目前對生物固氮速率及控制機理的認識尚很不足,導致生物固氮已成為氮循環主要的不確定性環節,在生態模型中仍不能通過機理過程進行模擬[5]。盡管有研究已經對不同生態系統的生物固氮速率進行了測定,但是目前的研究相當有限,對任何生物區系(Biome)生物固氮速率的測定都遠沒有代表性。Reed等[3]搜集了全球生物固氮速率數據,發現共生與非共生固氮速率的實測數據分別有52個和114個;全球不同生態系統的共生固氮速率介于0.1~160 kg N/(hm2·a),而非共生固氮速率介于0.01~60 kg N/(hm2·a)。但是對于共生與非共生固氮對生態系統生物固氮總通量的相對貢獻尚存在很大爭議,其中一個重要原因是很少有研究同時測定了共生與非共生固氮[6]。傳統觀點認為共生固氮占主導地位,但是近年來的觀點認為非共生固氮同樣重要,甚至在許多生態系統中占主導地位[3,6]。這是因為固氮植物空間分布上不均勻,許多生態系統中固氮植物數量少甚至缺乏[7]。考慮到現有研究的局限性,無疑需要開展更多的研究以獲取代表性數據及揭示生物固氮的控制機理,如揭示生物固氮隨生態演替變化特征。
生物固氮可受多種因素影響,如pH、養分有效性、溫濕度等[3,8-9]。在生態演替過程中,往往伴隨著植物群落、土壤溫濕度、養分有效性等環境因子的變化,這些都會深刻影響到生物固氮速率。研究發現,共生固氮速率往往會隨著生態演替進程而降低[10-11]。如Pearson等[12]發現夏威夷Acacia koa林6年齡階段的共生固氮速率為8 kg N/(hm2·a),而到了成熟林階段共生固氮速率小于2 kg N/(hm2·a)。Augusto等[10]報道了6年齡海岸松林中的荊豆屬植物(Ulex)固氮通量介于8.1~57.4 kg N/(hm2·a),而成熟海岸松林中的荊豆屬植物固氮通量則降至
0.5~5.1 kg N/(hm2·a)。這可能與演替過程中生態系統氮水平升高而固氮植物豐度降低有關[11,13]。相對而言,目前關于非共生固氮速率如何隨生態演替變化的報道較少,且結果不盡一致。例如,DeLuca等[14]發現在北方地區苔蘚非共生固氮速率在生態演替后期顯著降低。Vitousek[15]發現在夏威夷的一個生態演替序列中非共生固氮量(地衣、凋落物和腐木等)從0.3 kg N/(hm2·a)顯著增加到2.8 kg N/(hm2·a)。總之,現有研究對非共生固氮速率隨生態演替變化特征的認識仍很不足。
我國西南喀斯特山區生態環境十分脆弱,過去由于不合理的人為活動,大面積的喀斯特生態系統遭到破壞,導致生態退化[16]。近20年來,國家先后在西南喀斯特山區實施了一系列重大生態恢復工程,因此,目前大面積的喀斯特生態系統處于退耕后不同恢復階段[17]。前期的研究表明,喀斯特山區退耕后土壤總氮和有效氮能快速累積[2,18],在退耕后自然演替過程中,草叢階段受氮限制,灌叢階段受氮和磷共同限制,而森林階段則表現出明顯的氮飽和特征[2,19]。然而目前我們對退耕后演替過程中土壤氮累積機理的認識尚很不足。由于非共生固氮是生態系統外源氮輸入的重要途徑[3],我們猜測非共生固氮對退耕后演替過程中土壤氮累積具有重要意義。然而,由于缺乏研究,我們尚不清楚西南喀斯特山區退耕后演替過程中非共生固氮速率如何變化。以往研究針對生態演替過程中土壤固氮菌群落數量與多樣性方面作了探討,例如,魏媛等[20]研究發現喀斯特山區生態演替過程中土壤固氮菌豐度持續上升,認為隨著生態系統的正向演替,土壤從大氣中獲取氮素的能力也在不斷增強。鐘熙敏等[21]發現生態演替過程中土壤固氮菌種群數量增加,群落多樣性也更加豐富。然而,固氮菌群落的變化并不能直接反映生物固氮速率的變化特征。
本研究基于桂西北喀斯特山區一條退耕后的演替序列(包括草叢、灌叢和次生林),研究了土壤、凋落物和苔蘚的非共生固氮速率隨演替變化特征,并分析了主控因素。由于前期研究表明氮狀況隨演替進程增加[2,18],而有效氮水平對非共生固氮速率具有抑制作用[14],我們猜測土壤和凋落物的非共生固氮速率隨演替進程而下降(猜測1)。苔蘚的養分主要來源于大氣沉降,考慮到不同演替階段距離較小,大氣氮等養分沉降通量理論上應類似,因此,我們猜測苔蘚的非共生固氮速率隨演替無明顯的變化特征(猜測2)。總之,本研究的主要目標包括:1)定量桂西北喀斯特山區典型生態系統非共生固氮速率;2)揭示桂西北喀斯特山區不同生態系統組分的非共生固氮速率隨演替變化特征;3)探討控制不同生態系統組分的非共生固氮速率隨演替變化的主要因素。
1 材料與方法
1.1 試驗地點
研究區位于廣西壯族自治區環江縣下南鄉的古周 村(107°56′~107°57′ E,24°54′~24°55′ N),屬 于典型的喀斯特峰叢洼地,海拔375~816 m。該地區的植被屬于亞熱帶濕潤常綠闊葉林,氣候為亞熱帶季風氣候。年平均氣溫為15.0~18.5 ℃,其中最低溫出現在1月,約3.4~8.7 ℃,最高溫出現在7月,約23.0~26.7 ℃。年平均降水量為1 530~1 820 mm,但降水季節分配不均,其中4月到8月為雨季,9月到次年3月為旱季。土壤為石灰土。
1.2 試驗設計
本試驗采用空間代替時間的方法選取退耕后不同演替階段的草叢、灌叢和次生林,每個階段設置3個20 m×20 m樣地,樣地之間距離大于20 m,不同演替階段的最大直線距離小于3 km。草叢的主要優勢種是斑茅(Saccharum arundinaceum),此外還有白茅(Imperata cylindrical(L.)Beauv.)、狗尾草(Setaria viridis(L.)Beauv.)等。灌叢的主要優勢種是黃荊(Vitex negundo(L.)(VIT)),其他物種有伊桐(Itoa orientalisHemsl.)、毛桐(Mallotus barbatus(Wall.ex Baill.)Muell.-Arg.)、香葉木(Spermadictyon suaveolensRoxb.)和雞仔木(Sinoadina racemose(Sieb.et Zucc.) Ridsd.)。次生林的主要物種為香椿(Toona sinensis),其他物種還有五角楓(Acer elegantulum(Fang et P. L.)Chiu)、斜葉榕(Ficus tinctoriaForst subsp. gibbosa(Bl.) Corner)和楤木(Aralia chinensis(L.))。所有樣方中均無固氮植物分布。
1.3 樣品采集
分別于2016年的非生長季和生長季在樣方內采集表層土壤(0~10 cm)、凋落物和苔蘚樣品。在每個樣方內,通過直徑為2 mm的土鉆按照“S”型取樣法(8~10個點)采集土壤樣品(剔除植物根系等雜物后充分混勻);分別通過20 cm×20 cm和10 cm×10 cm的正方形金屬框,根據所需樣品量采集一定面積的凋落物和苔蘚樣品。土壤樣品一部分用于原位測定非共生固氮速率,另一部分帶回實驗室過孔徑為1 mm篩后測定土壤銨態氮和硝態氮含量,最后一部分在室內風干后過孔徑為1 mm或0.15 mm篩測定土壤理化指標。凋落物和苔蘚樣品一份用于原位測定非共生固氮速率,另一部分帶回實驗室65 ℃烘干粉碎后測定理化指標。
1.4 非共生固氮速率的測定
采用乙炔還原法在原位條件下測定土壤、凋落物和苔蘚的乙烯產生速率[22]。采集一定質量的樣品(其中土壤20 g、凋落物7 g、苔蘚2 g)裝入密閉性良好的330 mL具塞的玻璃培養瓶中,蓋好瓶塞,用注射器抽出瓶中體積10%的頂空氣體,隨后往培養瓶中注入同等體積的高純乙炔(已通過98%的濃硫酸和5 mol/L NaOH溶液進行純化),將培養瓶置于樹蔭下避免陽光直射,在原位條件下培養24 h后用注射器抽取瓶中30 mL頂空氣體注入到已預先抽好真空的12 mL玻璃儲氣瓶(Labco Exetainer,Labco Limited,UK)中,帶回實驗室使用氣相色譜儀(GC-FID,Agilent GC 7890A,Agilent,USA)測定氣體樣品中的乙烯濃度。同步利用不注射乙炔的培養瓶和不裝入樣品的培養瓶分別檢測樣品本身和乙炔本身的乙烯產生情況。基于乙烯濃度和時間回歸分析獲得的斜率計算乙烯產生速率(nmol C2H4/(g·d))。隨后,利用我們前期針對喀斯特生態系統土壤、凋落物和苔蘚分別測定的乙烯產生速率與固氮速率之間的轉換系數[23],將本研究測得的乙烯產生速率轉換為固氮速率(ng N/(g·d))。
參照Reed等[24]的方法基于單位面積樣品的儲量和固氮速率估算非共生固氮年通量(kg N/(hm2·a))。單位面積土壤、凋落物和苔蘚樣品的儲量通過質量-面積轉換系數來估算,其中凋落物和苔蘚的質量-面積轉化系數分別通過20 cm×20 cm、10 cm×10 cm的正方形金屬框中凋落物和苔蘚干重來計算,土壤則結合土壤容重和巖石出露率計算其質量-面積轉換系數。
1.5 土壤、凋落物和苔蘚理化性質的分析
土壤、凋落物和苔蘚的含水率采用烘干法測定,土 壤pH用pH計(FE20K,Mettler-Toledo,Switzerland)測定(液土比為2.5:1)。土壤有機碳采用重鉻酸鉀氧化-硫酸外加熱法(油浴)測定(GB 8834—1988),土壤有效磷(AP)采用0.5 mol/L NaHCO3浸提-鉬銻抗比色法測定,土壤總磷采用H2SO4、HClO4消煮-鉬銻抗比色法測定,土壤全氮(TN)采用半微量開氏法-流動注射儀(Fiastar 5000)測定,土壤銨態氮(NH4+-N)和硝態氮(NO3--N)采用2 mol/L KCl溶液浸提-流動注射分析儀(AA3型)測定,凋落物和苔蘚中的碳和氮通過碳氮元素分析儀(EA 3000,EuroVector,Italy)測定,凋落物和苔蘚樣品的全磷采用H2SO4-H2O2消煮-鉬銻抗比色法測定[25]。
1.6 數據分析
運用SPSS 16軟件(SPSS Inc.,Chicago,IL,USA)對數據進行統計分析。所有數據均進行正態性和方差同質性檢驗。采用多因素方差分析檢驗演替階段、生態系統組分和季節以及它們的交互作用對非共生固氮速率的影響。采用單因素方差分析檢驗不同演替階段之間非共生固氮速率、非共生固氮年通量和理化性質的差異是否顯著。采用皮爾遜相關分析確定固氮速率與理化指標之間的相關性,采用多元線性回歸分析確定非共生固氮速率的最強解釋因子。
2 結果與分析
2.1 非共生固氮速率隨生態演替的變化特征
多因素方差分析顯示生態系統組分、演替階段、季節及其交互作用對非共生固氮速率均有顯著影響(表1,p< 0.05)。在非生長季,土壤和凋落物非共生固氮速率均表現為次生林最高,灌叢最低,隨生態演替呈先降低后增加的趨勢(圖1a,圖1b);苔蘚固氮速率在草叢和灌叢階段相當,次生林階段最高(圖1c)。在生長季,土壤和苔蘚的非共生固氮速率在草叢和灌叢階段沒有顯著差異,在次生林階段最高(圖1a,圖1c)。但凋落物的非共生固氮速率草叢階段最高,在灌叢階段最低(圖1b)。
全年總體來看,土壤的非共生固氮速率在次生林階段最高((2.99 ± 0.10) ng N/(g·d)),比草叢階段增加了50.19%;在灌叢階段最低((1.31 ± 0.17) ng N/(g·d)),比草叢階段下降了34.17%(圖1a)。凋落物的非共生固氮速率在草叢階段最高((3 914.90± 255.33) ng N/(g·d)),而在灌叢階段最低((2.62 ±
0.23) ng N/(g·d)),灌叢和次生林凋落物的非共生固氮速率分別比草叢階段下降了99.93%和96.92%(圖1b)。次生林苔蘚的非共生固氮速率((669.45± 60.08) ng N/(g·d))分別比草叢和灌叢階段增加了
301.57%和329.31%(圖1c)。

表1 生態系統組分、演替階段、季節及其交互作用對非共生固氮速率的影響Table 1 Effects of ecosystem compartment, succession stage, season and their interactions on ANF rate
2.2 非共生固氮年通量隨生態演替的變化特征
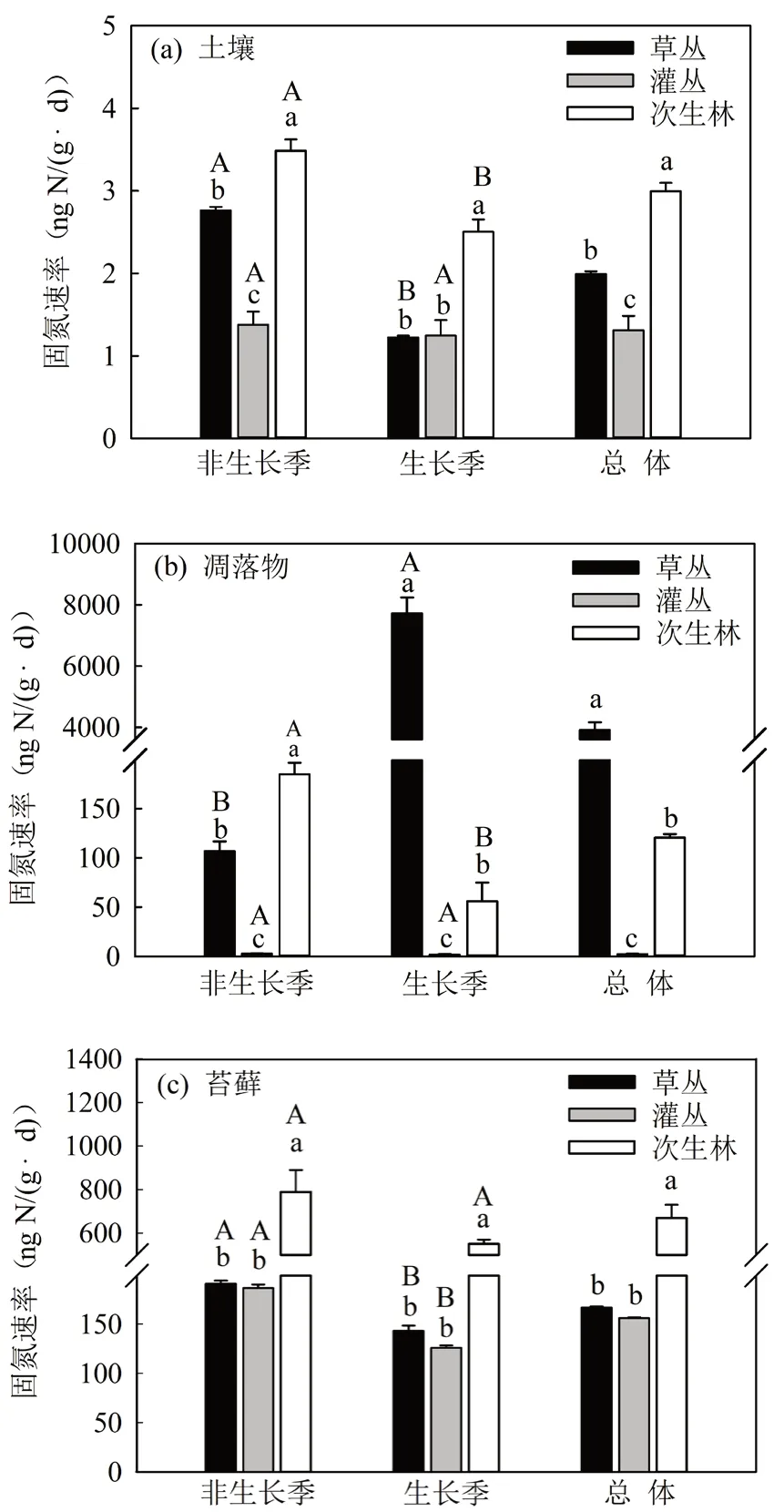
圖1 土壤、凋落物和苔蘚非共生固氮速率隨生態演替變化特征Fig. 1 Changes of ANF rate in soil, litter and moss with ecological succession

圖2 非共生固氮年通量隨生態演替變化特征Fig. 2 Changes of annual ANF rate with ecological succession
不同生態系統組分的非共生固氮年通量隨生態演替變化特征與各自的全年總體固氮速率變化特征類似(圖1,圖2)。土壤和苔蘚的非共生固氮年通量隨演替進程先小幅下降,隨后顯著上升,而凋落物的非共生固氮年通量隨演替進程顯著下降(圖2)。基于上述三個組分得到的生態系統非共生固氮年通量隨生態演替呈現先下降而后增加的特征,草叢、灌叢和次生林的非共生固氮年通量分別為(1.21± 0.06) kg N/(hm2·a),(0.23 ± 0.03) kg N/(hm2·a)和(0.65± 0.04) kg N/(hm2·a)(圖2)。
不同組分對生態系統非共生固氮年通量的相對貢獻隨演替變化特征存在差異(圖3)。土壤對生態系統非共生固氮年通量的相對貢獻以灌叢最高(80.71%)而草叢最低(21.89%);凋落物的相對貢獻則以草叢最高(70.71%)而灌叢最低(0.28%);苔蘚的相對貢獻隨演替而增加,從草叢的7.40%增加到次生林的39.87%。不同組分對生態系統非共生固氮年通量的相對貢獻在不同演替階段也存在差異。在草叢階段,相對貢獻以凋落物最高而苔蘚最低;在灌叢和次生林階段,相對貢獻則以土壤最高而凋落物最低。
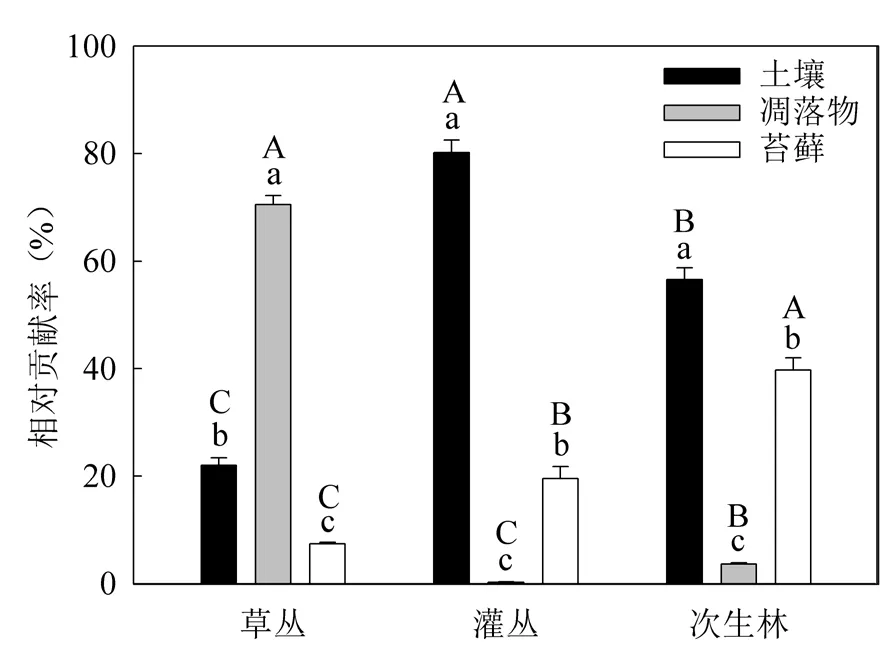
圖3 土壤、凋落物和苔蘚對不同生態系統非共生固氮年通量的相對貢獻Fig. 3 The relative contributions of soil, litter and moss to annual ANF rate in different ecosystems
2.3 土壤、凋落物和苔蘚理化性質及其與非共生固氮速率的相關性
土壤理化性質隨生態演替的變化特征因指標而異(表2)。土壤有機碳和全氮含量在草叢階段最低,但在灌叢和次生林二個階段之間無顯著差異。土壤全磷含量和C∶P比值在三個演替階段之間無顯著差異。土壤C∶N比草叢階段最高,而次生林階段最低;土壤N∶P比隨演替的變化特征則與C∶N比相反。生長季和非生長季土壤含水率隨生態演替變化特征相似,均是次生林階段最高,且草叢和次生林在非生長季土壤含水率高于生長季。對于土壤無機氮含量,在非生長季時以次生林階段最高,而在草叢階段最低;生長季時三個階段之間無明顯差異;非生長季時的含量顯著高于生長季。對于土壤有效磷含量,生長季和非生長季均以次生林階段最低,但季節之間無顯著差異。
凋落物單位面積質量因季節而異,在生長季以草叢最高,次生林最低,而在非生長季以灌叢最高,草叢最低;且二個季節之間質量也存在顯著差異(表3)。凋落物含水率二個季節均以次生林最高而灌叢最低,且非生長季含水率高于生長季。凋落物碳含量在不同演替階段或不同季節之間均無顯著差異。凋落物氮和磷含量均表現為次生林最高,而草叢最低;草叢和灌叢凋落物氮和磷含量在生長季更高。二個季節凋落物C∶N和N∶P比值隨演替變化特征類似,C∶N比以草叢階段最高,而N∶P比在不同階段之間無顯著差異;草叢和灌叢凋落物C∶N比在非生長季更高。
二個季節苔蘚單位面積質量隨演替變化特征類似,草叢階段最高而灌叢階段最低;次生林苔蘚單位面積質量在非生長季更高,而其他二個階段則以生長季更高(表3)。生長季苔蘚的含水率在不同演替階段之間無顯著差異,而非生長季則是灌叢階段的含水率低于其他二個階段;非生長季的含水率顯著高于生長季。苔蘚碳含量在生長季存在演替階段之間的差異,即次生林高于其他二個階段,但生長季苔蘚氮和磷含量在演替階段之間無顯著差異。非生長季時苔蘚碳含量在演替階段之間無顯著差異,但氮和磷含量均以次生林最高而草叢最低。C∶N比值在生長季時各階段之間無顯著差異,但非生長季時以草叢階段較高。N∶P比均是次生林高于其他二個階段。
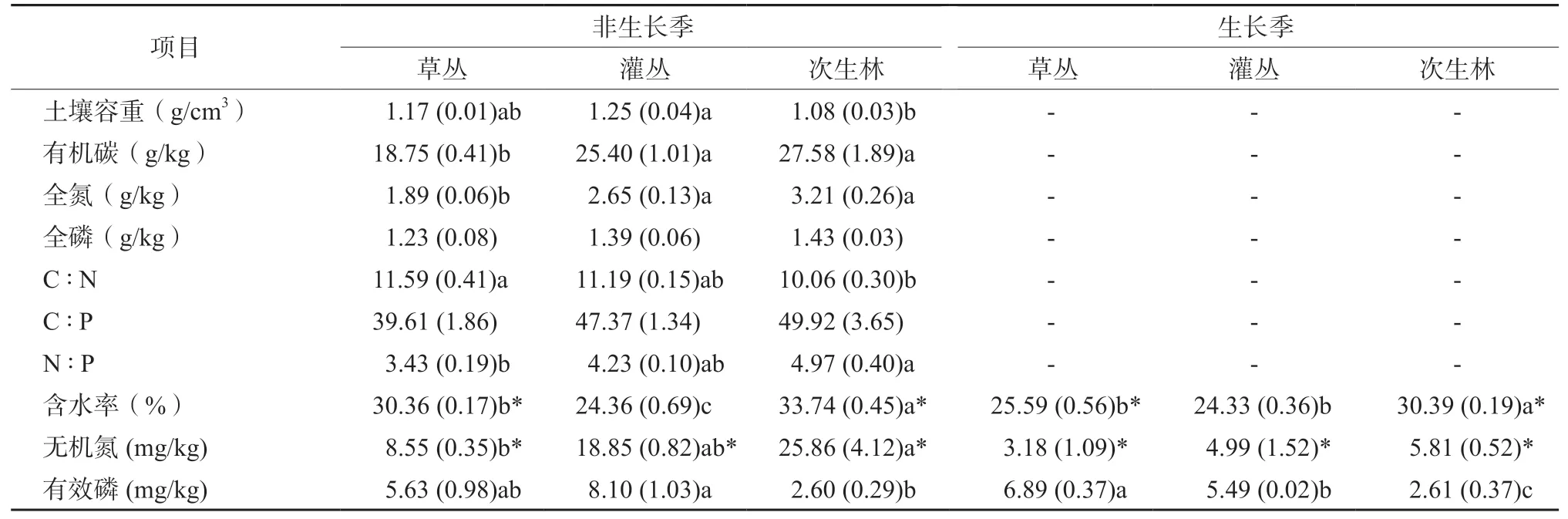
表2 三個不同演替階段土壤理化性質Table 2 Physicochemical properties for soil in grassland, shrubland and forest

表3 三個不同演替階段的凋落物和苔蘚理化性質Table 3 Physicochemical properties for litter and moss in grassland, shrubland and forest
多元線性逐步回歸分析結果表明,土壤的非共生固氮速率在生長季和非生長季的最強解釋因子均為含水率,分別解釋了非共生固氮速率變異的83.2%和94.9%(表4)。對于凋落物的非共生固氮速率,在生長季和非生長季的最強解釋因子分別為C∶N比和含水率,分別解釋了變異的86.9%和73.5%。對于苔蘚的非共生固氮速率,在生長季和非生長季的最強解釋因子分別是有機碳和全氮,分別解釋了變異的84.3%和46.8%。此外,皮爾遜相關分析顯示生長季時凋落物的非共生固氮速率與全氮呈顯著負相關(R2=0.67,p<0.01)。
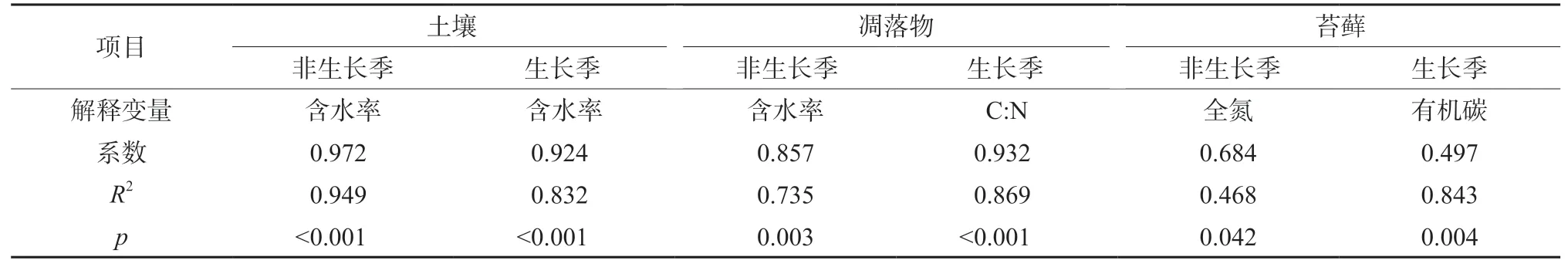
表4 多元線性逐步回歸顯示理化指標對非共生固氮速率的解釋度Table 4 Results of stepwise multiple linear regression analyses showing the dependence of asymbiotic N2 fixation rate on physicochemical variables
3 討論
本研究結果表明,桂西北喀斯特山區典型草叢、灌叢和次生林的非共生固氮年通量相比其他熱帶或亞熱帶地區較低。例如,有文獻報道的熱帶或亞熱帶草叢、灌叢和次生林非共生固氮速率分別介于0.1~21.0、0.63~0.97和0.1~60.0 kg N/(hm2·a)[3,26]。這可能與我國西南地區大氣氮沉降量以及土壤氮有效性相對較高,非共生固氮速率受到了抑制有關[18,27-28]。
我們發現喀斯特山區非共生固氮年通量隨生態演替呈先降低后升高的趨勢,有別于其他研究發現的非共生固氮速率隨生態演替進程而降低[4,29-30]或升高[28]的現象。在本研究中非共生固氮年通量在草叢階段較高,其中凋落物非共生固氮對草叢非共生固氮年通量貢獻最大(70.71%),可能與草叢凋落物中氮含量低而C∶N比高有關,這兩個條件在前期研究中均被證明對非共生固氮具有促進作用[3,31]。灌叢階段非共生固氮年通量較低,這可能與灌叢階段土壤、凋落物含水率均較低而氮含量較高有關。以往大量研究表明,在干旱和富氮環境條件下土壤和凋落物非共生固氮速率受到明顯抑制[32-33]。相比于灌叢階段,次生林階段非共生固氮年通量略有增加。在以往的研究中也有類似發現[34],可能是因為次生林中土壤含水率以及苔蘚碳氮含量較高促進了土壤和苔蘚的非共生固氮速率。
不同生態系統組分非共生固氮速率和年通量隨生態演替變化的趨勢不同。首先,土壤的非共生固氮速率和年通量隨生態演替呈先下降后上升的趨勢,主要與土壤含水率隨生態演替的變化有關。其次,凋落物非共生固氮速率和年通量在演替中后期下降,這與Pérez等[35]的研究結果一致。在本研究中凋落物C∶N比在演替過程中不斷降低,可能是導致凋落物非共生固氮速率和年通量在演替中后期降低的主要原因。最后,苔蘚非共生固氮速率和年通量在演替早期和中期無顯著差異,在后期上升,以往的研究中也有類似的結果,且非共生固氮年通量與我們的研究處于同一水平(1~2 kg N/(hm2·a))[36-37]。可能與次生林中苔蘚碳氮含量較高有關。我們發現含水率是影響土壤和非生長季凋落物非共生固氮的關鍵因子。以往的研究也表明了含水率對非共生固氮的促進作用[32,38]。例如,我們前期的研究也發現喀斯特森林土壤和凋落物非共生固氮速率隨土壤含水率的增加而增加[32]。這一方面可能是因為固氮酶對氧氣十分敏感,適當的水分條件可以為微生物創造厭氧條件,防止固氮酶被氧氣破壞[8,33]。另一方面可能是因為含水率增加提升了土壤或凋落物固氮功能基因(nifH)的相對豐度[39]。與非生長季期間不同,我們發現C∶N比對凋落物生長季期間非共生固氮速率的影響程度最大。由于生物固氮過程需要消耗大量的能量用于打破N2中的化學鍵以及保持固氮酶活性[40],而凋落物中較高的C∶N比可以為固氮微生物提供更多固氮碳和能量,因此對非共生固氮具有促進作用。這與Zheng等[31]的研究一致。此外,我們還發現氮對凋落物和苔蘚非共生固氮速率有顯著影響,且存在不同效應。其中,在生長季期間凋落物的非共生固氮速率與氮含量呈負相關(R2=0.669,p<0.01)。以往許多研究也表明了環境氮水平的升高對非共生固氮的抑制作用[41-42],我們前期的研究也表明喀斯特森林中凋落物非共生固氮速率隨氮添加濃度的增加而降低[23]。這是因為生物固氮極其消耗固氮微生物自身養分(碳、氮、磷等)和能量[30],固氮微生物在富氮環境中傾向于降低固氮速率而直接吸收利用環境中的氮[33,43]。研究表明,高氮條件會降低固氮菌和固氮基因的豐度[44],增加固氮基因擴增的困難性[45]。而苔蘚在非生長季期間非共生固氮速率與氮含量呈顯著正相關。Wang等[23]的研究也報導了喀斯特山區苔蘚非共生固氮速率對環境氮狀況的響應有別于土壤和凋落物。這與苔蘚獲取氮的方式有關,因為苔蘚不從土壤中獲取生長需要的氮,需要通過固氮來滿足自身氮素需求[46]。由于固氮酶的合成需要消耗一定量的氮,苔蘚氮含量更高意味著可以合成更多的固氮酶[33],因而對非共生固氮具有促進作用。意外的是,本研究中并未發現生態演替過程中磷對非共生固氮速率的影響,這與以往研究報導的磷對非共生固氮具有促進作用的結果不同[47-48]。可能是由于喀斯特山區非共生固氮速率受含水率、氮等的影響更大,導致磷的促進效應不明顯。
4 結論
本文研究了桂西北喀斯特山區土壤、凋落物和苔蘚的非共生固氮速率隨退耕后生態演替的變化特征,發現從演替早期(草叢)到后期階段(次生林),非共生固氮年通量呈現先下降后上升的變化趨勢,且土壤、凋落物和苔蘚非共生固氮速率和年通量隨生態演替的變化趨勢不一,主要受生態演替過程中土壤、凋落物和苔蘚含水量、有機碳、全氮和C∶N比變化的影響。研究結果有助于深入認識喀斯特生態系統氮循環特征,并為生態恢復過程中氮累積提供了一種機理解釋。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
