幾株偽狂犬病病毒的增殖特性與保存條件
2020-08-04 12:30:35陳賽賽鄭亞婷郭容利
江蘇農業科學 2020年11期
陳賽賽 鄭亞婷 郭容利
摘要:高滴度的抗原是制備優質疫苗的首要條件。對6株偽狂犬病病毒在倉鼠腎(baby hamster kidney,簡稱BHK)細胞和豬睪丸細胞(swine testicular,簡稱ST)細胞上的增殖效率進行了比較,同時測定不同保存溫度和凍融條件對病毒滴度的影響。結果表明,幾株偽狂犬病病毒在BHK細胞和ST細胞上增殖效率均很高,可達到生產要求。相較于ST細胞,各毒株在BHK細胞上出現病變時間更早。然而,所有毒株在ST細胞上的病毒滴度都高于同期在BHK細胞上的滴度。強毒株AH02LA株在2種細胞上滴度可達108.0 TCID50/0.1 mL以上,表現為發生病變早,病毒增殖快,峰值到來早的特性。通過強毒株AH02LA株獲得的基因缺失株LA-A株和自然傳代缺失致弱株LA2017株峰值均達108.0 TCID50/0.1 mL,展現出優良疫苗株的潛質。保存條件試驗表明,PRV在-70 ℃保存30 d和4 ℃保存12 d內滴度相對穩定,而-20 ℃保存的病毒滴度下降明顯。多次凍融后,滴度有所下降,相較于-70 ℃,-20 ℃條件下凍融病毒滴度下降更為顯著,建議避免多次凍融。本試驗結果可為PRV疫苗抗原在實際生產中制備與保存提供科學依據。
關鍵詞:偽狂犬病病毒;豬睪丸細胞;倉鼠腎細胞;生長特性;凍融;保存
中圖分類號: S852.65+5 ?文獻標志碼: A ?文章編號:1002-1302(2020)11-0148-05
收稿日期:2020-04-23
基金項目:江蘇省農業科技自主創新資金[編號:CX(18)3020];江蘇省自然科學基金(編號:BK20181243)。
作者簡介:陳賽賽(1993—),男,山西長治人,研究實習員,主要從事豬偽狂犬病疫苗及其防控技術的研究。E-mail:1304652436@qq.com。
通信作者:王繼春,男,江蘇南京人,博士,研究員,主要從事活載體疫苗和基因工程疫苗研究。E-mail:jcwang@263.net。 ?偽狂犬病是由偽狂犬病病毒(pseudorabies virus,簡稱PRV)引起的以家畜發熱、奇癢(豬除外)和腦脊髓炎為主要特征的烈性傳染病,又稱Aujeszky病[1]。該病毒可感染不同日齡的豬,妊娠母豬感染后引起流產、產木乃伊胎或死胎,初生仔豬感染死亡率可達100%,成年豬一般為隱性感染,不表現臨床癥狀,但長期帶毒和排毒。在我國該病已涉及包括香港、臺灣在內幾乎所有地區[2]。美洲和歐洲許多國家通過應用基因缺失弱毒苗免疫,配合鑒別診斷等綜合防控措施,成功將該病從家養豬場根除[3],我國許多豬場也應用類似方法有效控制了該病。但自2011年以來,在我國一些Bartha株疫苗免疫的豬場又有該病的發生[4-6],其特征為母豬產弱仔、死胎,流產,仔豬出現神經癥狀等臨床癥狀,且死亡率較高,其地域包括黑龍江省、吉林省、天津市、河北省、湖北省、四川省、江蘇省、廣東省等,給我國養豬業造成極大的經濟損失[4-6],分析表明疫情由PRV變異株所引起。
疫苗接種是控制豬偽狂犬病最重要的措施,而免疫效果很大程度取決于抗原滴度。目前市場上所售的豬偽狂犬病疫苗在制備抗原時所用的細胞主要是雞胚成纖維細胞,倉鼠腎(baby hamster kidney,BHK)細胞和豬睪丸細胞(swine testicular,簡稱ST)細胞,后2種細胞便于利用微載體培養和懸浮培養提高抗原滴度,是制備豬偽狂犬病疫苗的首選。
本試驗將6株PRV在BHK和ST細胞上的增殖效率進行了比較,并且對不同保存條件下PRV的滴度變化進行了初步研究,以期為篩選疫苗候選株提供依據,同時為豬偽狂犬病疫苗生產工藝的研究提供基礎。
1 材料與方法
1.1 病毒、細胞和主要試劑
病毒:Bartha K61株,購自西班牙海博萊生物大藥廠,產品批次為66HP[7];AH02LA株為筆者所在實驗室分離的PRV變異株[6],保藏號碼為:CGMCC No.10891;B-gD & gCS株是以Bartha K61株為母本,將其gD和gC基因替換為流行變異株AH02LA株的gD和gC基因的重組病毒株[8];LA2017株是以AH02LA株為母本經連續傳代獲得的致弱疫苗株,該毒株缺失了從gI基因第118位核苷酸至28K基因第251位核苷酸的共3 686堿基,包括gI部分基因、gE全部基因、11K全部基因和28K部分基因,保藏號碼為:CGMCC No.18170;LA1206株,是以AH02LA株為母本,人工缺失gE和TK基因,再經連續傳代80代獲得的弱毒疫苗株,保藏號碼為:CGMCC No.14329[9];LA-A株,是以AH02LA株為母本,人工缺失gE而得,用作PRV基因缺失滅活疫苗的種毒[10]。
細胞:BHK細胞和ST細胞,由筆者所在單位細胞工程組提供。
主要試劑:新生牛血清(newborn bovine serum,簡稱NBS)和DMEM培養基為Gibco公司產品。
1.2 病毒的培養與收獲
取PRV各毒株,按0.01感染復數(multiplicity of infection,簡稱MOI)接種長滿單層的BHK或ST細胞,吸附1 h后,吸去病毒液,應用無菌PBS液洗滌3次,加2% NBS的DMEM培養液37 ℃培養,待細胞病變達80%后,將上清與細胞混合物收集,在-70 ℃和37 ℃凍融,經8 000 r/min離心,取上清分裝置-70 ℃保存。
1.3 病毒含量的測定
BHK和ST細胞傳至96孔板長滿單層,將PRV各毒株倍比稀釋至10-1~10-9,每一稀釋度接種8孔,每孔100 μL,對照組加100 μL細胞營養液,37 ℃ 孵育1 h后加入2% NBS的DMEM培養液100 μL,每日觀察細胞病變,觀察5 d,記錄細胞病變情況,按Reed-Muench法計算細胞半數感染量(median tissue culture infectious dose,簡稱TCID50)。
1.4 病毒蝕斑形成特性
BHK和ST細胞傳至六孔板長滿單層,接種病毒稀釋液,每孔100 μL,含50~100 TCID50,對照組加100 μL細胞營養液,37 ℃孵育1 h后加入2% NBS的DMEM培養液100 μL,每隔3~5 h觀察細胞病變情況。
1.5 生長曲線測定
將PRV各病毒株分別以0.01 MOI接種長滿單層的6孔板上的BHK或ST細胞,37 ℃孵育1 h,吸去上清,PBS洗滌3次,添加含2% NBS的DMEM培養液,每孔2 mL,37 ℃培養,分別于接種后6、12、24、36、48、72 h測定培養物中病毒含量,統計所有時間點的病毒滴度,繪制生長曲線。
1.6 不同溫度和多次凍融對病毒滴度的影響
將相同滴度的Bartha K61株和AH02LA株分別放置在37、4、-20、-70 ℃一段時間后,檢測其病毒的滴度,如“1.3”節所述。并且在-20 ℃和-70 ℃條件下,短時間內(72 h完成7次凍融)反復凍融,檢測其病毒的滴度,如“1.3”節所述。
2 結果與分析
2.1 病毒含量
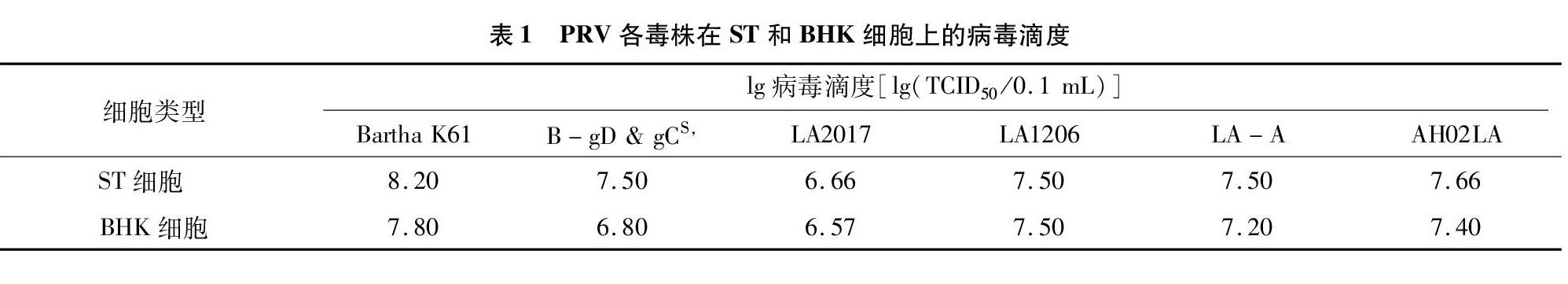
從表1可以看出,各毒株在ST和BHK細胞上適應性良好,滴度達106.57 TCID50/0.1 mL以上,可滿足生產需要。
2.2 病毒蝕斑特性
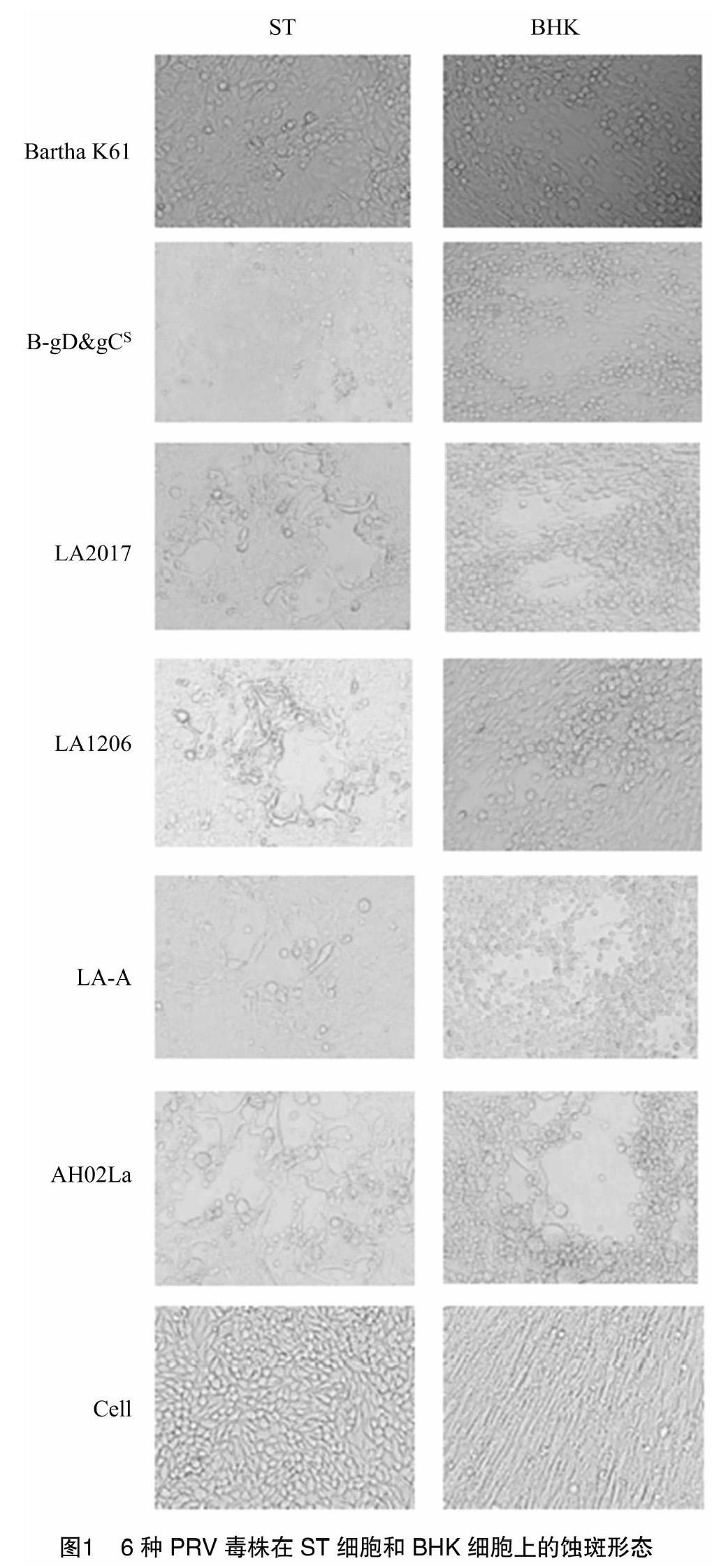
6種PRV毒株在ST和BHK細胞上病變明顯。接種相同劑量的病毒,病變發生的進度不完全一致,Bartha K61和B-gD & gCS在ST細胞上的病變時間明顯滯后于其他毒株,且病變形態表現為細胞堆積,顆粒增多。AH02LA強毒株以及由AH02LA株衍生出來的LA2017株、LA1206株和LA-A株,在ST細胞上產生病變的形態基本一致,空斑明顯,出現多數大小不一的融合細胞,AH02LA強毒株病變進展最快,大小空斑連成一片,融合細胞多且大。AH02LA強毒株在BHK細胞上也出現明顯的融合細胞,而其他毒株在BHK上的融合細胞不明顯,主要出現空斑和細胞變圓等現象(圖1)。相對于ST細胞,所有PRV在BHK細胞上出現病變的時間更早。
2.3 生長動力學測定
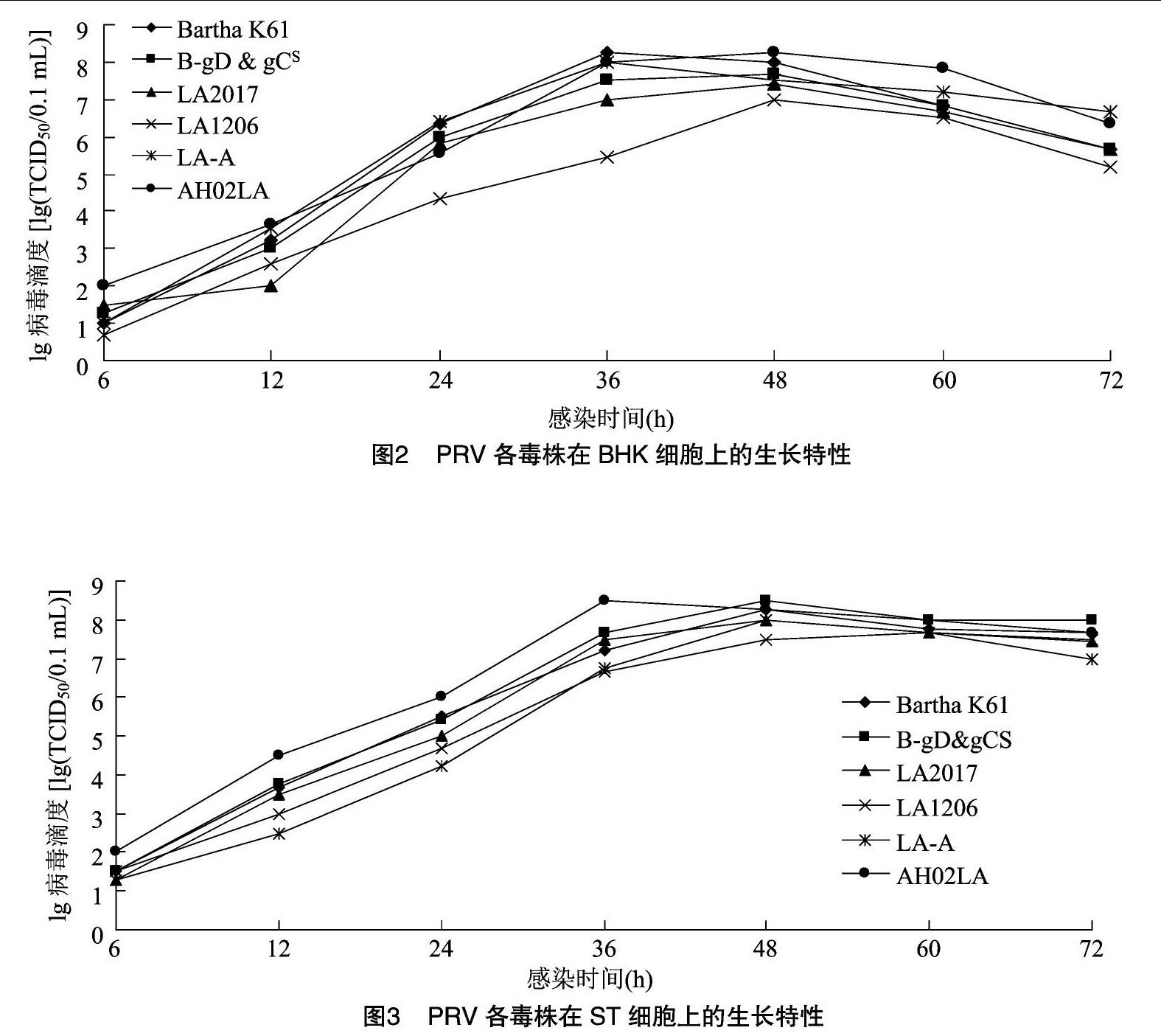
從圖2、圖3可以看出,各毒株滴度峰值出現在36~48 h之間。對于BHK細胞,大多數毒株的峰值出現在36 h。而在ST細胞上,除AH02LA毒株的峰值出現在36 h外,其余毒株的峰值都出現在48 h。Bartha K61株、基因缺失株LA-A以及由強毒株AH02LA自然傳代致弱的LA2017株其滴度峰值都能達到108.0 TCID50/0.1 mL,這十分有利于疫苗的研發。綜上所述,強毒株在2種細胞上增殖滴度均能達到108.0 TCID50/0.1 mL以上,表現為發生病變早,病毒增殖快,峰值到來早。各毒株在ST細胞上的滴度峰值均大于同期在BHK細胞上的值。
2.4 不同保存溫度對病毒滴度的影響
將相同滴度(106.0 TCID50/0.1 mL)的Bartha K61株和AH02LA株樣品分別放置在 37、4、-20、
-70 ℃,一段時間后檢測病毒滴度。結果發現病毒液在37 ℃保存3~12 d,其滴度下降趨勢十分明顯。-70 ℃保存30 d,滴度變化不大;4 ℃保存3~12 d,滴度基本無變化;而-20 ℃保存7 d,病毒滴度下降非常明顯(表2)。說明Bartha K61株和AH02LA株在 -70 ℃ 和4 ℃保存12 d內穩定性較好。
2.5 多次凍融對病毒滴度的影響
將相同滴度的Bartha K61株和AH02LA株樣品分別放置在-20、-70 ℃, 短時間內(72 h完成7次)反復凍融,檢測滴度變化。結果表明,Bartha K61株與AH02LA株-20、-70 ℃凍融多次后滴度有降低趨勢。-20 ℃凍融7次后滴度下降接近90%,-70 ℃ 條件下,3次凍融滴度下降較低。多次凍融后,效價有所下降,-20 ℃條件下凍融比 -70 ℃ 條件下凍融滴度下降趨勢明顯(表3)。
3 討論與結論
PRV可在多種細胞上增殖,如豬腎細胞、非洲綠猴腎細胞、ST細胞、BHK細胞、牛腎細胞和雞成纖維細胞[11-15]。基于對PRV的敏感性和增殖效率
以及規模化生產等因素,通常選取BHK細胞與ST細胞作為最終的生產用細胞。本試驗將6株PRV在BHK和ST細胞上的增殖效率進行了比較。試驗中所有毒株均能在BHK和ST細胞上產生明顯病變,強弱毒株病變的形態存在明顯差異,強毒株表現出巨融合細胞的出現以及病變時間超前等特性,本結論與程曉霞等報道的結果[13]一致。觀察體外生長特性發現病毒增殖的整個過程經歷增殖期和高峰期,而后出現下滑期。對于BHK細胞,多數毒株的峰值出現在36 h,而在ST細胞上峰值出現在48 h(強毒株AH02LA除外),說明PRV在BHK細胞上增殖速度較快。此外,研究發現基因缺失株 LA-A 株、由強毒株AH02LA自然傳代致弱的LA2017株和經典的弱毒株Bartha K61株在BHK和ST細胞上的滴度峰值都能達到108.0 TCID50/0.1 mL以上,這十分有利于PRV疫苗抗原的制備。在實際的生產中,ST細胞可用于活疫苗與滅活疫苗的生產,本研究發現PRV在BHK細胞上出現病變的時間較早,病毒含量較高,在節省時間的同時,又能獲得較高的抗原濃度,因此,BHK細胞也可作為PRV增殖的理想細胞,考慮到其致瘤性的隱患,一般用于滅活疫苗抗原的制備。
此外,在PRV抗原實際的生產中,在抗原收獲后需要進行短時間保存,并且需要凍融離心去除細胞碎片,因此,本試驗研究了抗原在-70、-20、4、37 ℃ 溫度條件下短期保存對病毒滴度的影響。結果表明,病毒不宜-20 ℃保存,-70 ℃是理想的保存溫度,但是實際生產中抗原產量較大,-70 ℃保存需要消耗大量能源,因此可選擇4 ℃短期保存(不超過12 d)。病毒凍融試驗結果顯示,PRV抗原-20、-70 ℃條件下凍融多次后滴度均有所下降,為維持抗原穩定,應避免多次凍融,必要時可選擇-70 ℃條件下凍融3次以內。
綜上所述,本研究從實際生產出發,開展的PRV生長特性以及不同保存溫度和反復凍融對病毒滴度的影響,其目的是提高抗原滴度,保護抗原活力,為疫苗的生產提供科學依據。
參考文獻:
[1]殷 震,劉景華. 動物病毒學[M]. 2版.北京:科學出版社,1997:700-713.
[2]鄧仕偉,汪 勇,薛春芳. 我國偽狂犬病流行現狀及新特點[J]. 動物醫學進展,2006,27(9):105-107.
[3]Ketusing N,Reeves A,Portacci K,et al. Evaluation of strategies for the eradication of pseudorabies virus (Aujeszkys disease) in commercial swine farms in Chiang-Mai and Lampoon provinces,Thailand,using a simulation disease spread model[J]. Transboundary and Emerging Diseases,2014,61(2):169-176.
[4]彭金美,安同慶,趙鴻遠,等. 豬偽狂犬病病毒新流行株的分離鑒定及抗原差異性分析[J]. 中國預防獸醫學報,2013,35(1):1-4.
[5]An T Q,Peng J M,Tian Z J,et al. Pseudorabies virus variant in Bartha-K61-vaccinated pigs,China,2012[J]. Emerging Infectious Diseases,2013,19(11):1749-1755.
[6]喬永峰,顧一奇,柳 暢,等. 豬偽狂犬病病毒AH02LA株的分離鑒定及其對仔豬致病性研究[J]. 畜牧與獸醫,2016,48(12):36-41.
[7]王繼春,曾容愚,Torrents D,等. 豬偽狂犬病活疫苗(Bartha K61株)對變異株的保護效力[J]. 畜牧與獸醫,2015,47(12):1-4.
[8]Zhang C J,Liu Y M,Chen S,et al. A gD & gC-substituted pseudorabies virus vaccine strain provides complete clinical protection and is helpful to prevent virus shedding against challenge by a Chinese pseudorabies variant[J]. BMC Veterinary Research,2019,15:2.
[9]劉婭梅,喬永峰,顧一奇,等. 豬源偽狂犬病病毒變異株AH02LA株細菌人工染色體的構建與鑒定[J]. 中國預防獸醫學報,2018,40(10):880-885.
[10]Wang J C,Guo R L,Qiao Y F,et al. An inactivated gE-deleted pseudorabies vaccine provides complete clinical protection and reduces virus shedding against challenge by a Chinese pseudorabies variant[J]. BMC Veterinary Research,2016,12:277.
[11]李小靜,唐滿華. 豬偽狂犬病病毒在不同細胞上的增殖研究[J]. 吉林畜牧獸醫,2015,36(6):14-16.
[12]王 巖,楊明凡,崔保安,等. PK15、Vero、BHK-21、CEF細胞增殖豬偽狂犬病病毒的比較[J]. 安徽農業科學,2007,35(18):5432-5445.
[13]程曉霞,陳少鶯,胡奇林,等. 豬偽狂犬病病毒在幾種細胞中增殖情況的比較[J]. 福建畜牧獸醫,2006,28(6):37-38.
[14]王樹成,趙祥平,董志珍,等. 用PK15、VERO和MDBK三種細胞增殖豬偽狂犬病病毒的比較[J]. 畜牧與獸醫,1996,28(2):80.
[15]彭麗英,倪建平,張婉華,等. 豬偽狂犬病病毒在不同細胞中增殖的研究[J]. 上海農業學報,2011,27(4):67-69.任義軍,孫大明,俞曉鵬,等. 哺乳期麋鹿的生長發育與疾病發生的關系[J]. 江蘇農業科學,2020,48(11):153-157.
