取食玉米行為對散放丹頂鶴繁殖期能量攝入及卵殼特征的影響
2020-08-03 06:36:58楊志宏鄒紅菲
野生動物學報 2020年3期
關鍵詞:質量
楊志宏 鄒紅菲
(1.齊齊哈爾大學生命科學與農林學院,齊齊哈爾,161006;2.東北林業(yè)大學野生動物與自然保護地學院,哈爾濱,150040)
扎龍濕地總面積約為21萬hm2(亞洲第一,世界第四),是世界最大的蘆葦(Phragmitesaustralis)濕地(蘆葦沼澤面積約為10萬hm2、明水面積約2萬hm2、草甸草原面積約6萬hm2和農田面積約3萬hm2),有著極為豐富的生物多樣性,同時也是丹頂鶴(Grusjaponensis)等許多鳥類的主要繁殖地[1-2]。扎龍國家級自然保護區(qū)(46°52′—47°32′N,123°47′—123°37′E)是我國最早成立的國家級自然保護區(qū)(1987年4月國務院批準),并被列為中國首批“世界重要濕地名錄”。保護區(qū)成立以來,制定了許多相關的法律法規(guī),生物多樣性得到有效保護。
丹頂鶴,屬鶴形目(Gruiformes),鶴科(Gruidae),為大型涉禽,雜食性鳥類(魚類、蝦、昆蟲、軟體動物、老鼠和蛇、植物的嫩芽和種子等),早成鳥,單配制,有補卵習性,棲息于濕地及沿海灘涂,蘆葦中營巢及繁殖期家域范圍較大,IUCN(2012)和《中國生物多樣性紅色名錄-脊椎動物卷》將其列為瀕危(endangered,EN)物種[3-4]。丹頂鶴的生存與繁殖一直備受世人關注。近期,有文獻報道丹頂鶴種群呈現減少趨勢[2,5],并認為是濕地生物資源減少[6]、棲息地喪失[7]和生境破碎化[8-9]等原因導致丹頂鶴的種群數量迅速下降。目前,每年春季,會有野生、散放和籠養(yǎng)三類繁殖丹頂鶴在扎龍濕地繁殖。保護區(qū)內的農田作為丹頂鶴的補充生境已經引起關注[10],其中農田生境的利用被認為是丹頂鶴對食物資源變化產生的一種適應[11-12]。
在扎龍濕地,丹頂鶴春季在已經播種的農田內掘土取食玉蜀黍(Zeamays,玉米)種子的行為并不常見。2016年春季,首次發(fā)現這對散放繁殖丹頂鶴連續(xù)數日取食播種農田內的玉米種子,直至其首次繁殖失敗后離開農田區(qū)域。那么,它們的取食效率如何?能夠滿足其攝入能量需求嗎?首次繁殖失敗與此有關嗎?本文對此進行相關的調查和分析。
1 研究對象與方法
1.1 農田與散放繁殖丹頂鶴
這塊面積約1 hm2的長方形農田位于扎龍濕地內,屬齊齊哈爾扎龍村村民的生產用地(周邊有蘆葦和雜草伴生)。經調查,此前,這塊農田一直用于播種玉米,2012—2015年農田休耕,2016年春季農田復耕和播種玉米。
扎龍保護區(qū)管理局近年來散放到扎龍濕地的丹頂鶴繁殖鶴對目前已有10余對,本文研究對象為散放繁殖鶴對中的一對(雌鶴環(huán)志編號為:ZL0452051;雄鶴環(huán)志編號為:ZL0452101),2013年春季散放(散放前,籠養(yǎng),經過野化訓飛等有關訓練,人工喂養(yǎng)期間供給主要食物和方式為鯽魚白天足量和玉米粒全天足量)。據調查,該對繁殖丹頂鶴從2013年春季散放開始至2015年春季都是在這塊農田區(qū)域內筑巢和繁殖。2012—2016年繁殖季節(jié),除該對散放繁殖丹頂鶴外,無其他丹頂鶴在此棲息。2016年春季3月開始,這對散放繁殖丹頂鶴一直在這塊農田區(qū)域內棲息,農田播種后,開始取食已播種農田內的玉米種子,大約10 d后,在農田東側附近區(qū)域的蘆葦叢中筑簡易巢并首次產2枚卵,隨即棄卵(卵已破碎)和離開。大約2周后,這對散放繁殖丹頂鶴再次筑巢和產卵于農田南側相距較遠的一片蘆葦叢中,再次筑巢,產卵2枚,孵化33 d后2只丹頂鶴幼鳥出生。
1.2 研究方法
1.2.1 取食行為觀察、日取食玉米質量和攝入能量的估算
用觀察法對其取食行為特征和取食地域選擇進行研究。利用DV錄像來記錄和獲取其取食頻率、每次取食持續(xù)時間和停歇時間等數據參數。DV錄像時間:從2016年5月13日上午開始至5月17日下午結束,共5 d,每天8:30—9:00、9:30—10:00、10:30—11:00和14:00—14:30、15:00—15:30、16:00—16:30共計6個時段進行錄像,每時段30 min,每天錄制時長3 h,總錄制時長15 h。每時段30 min中包括遠距離錄制雌雄丹頂鶴(互動行為)、拉近距離分別錄制雌鶴和雄鶴(掘食頻率和掘食行為差異)3部分,每部分約10 min左右,DV錄像人員距離丹頂鶴約50—80 m。數據處理方法:實驗室內重放錄像,對丹頂鶴掘土取食頻率、每次取食持續(xù)時間和停歇時間的規(guī)律性等信息進行采集和確認。通過實地用直尺測量平均取食的土壤深度和橫向土壤間距來估算其日均取食和攝入玉米種子的質量和能量。據了解,2016年5月初開始農戶采用播種機種植玉米,壟間行距為60 cm,播種方式為穴播,1粒(測得平均每顆玉米種子質量約0.4 g)/每株(精播),株距為15 cm。農田開始播種至5月8日無法證實丹頂鶴取食玉米種子,5月9—11日丹頂鶴取食可以確定。實驗是從2016年5月12日清晨開始至5月18日傍晚結束,共7 d。取食效率的測量方法:在農田被取食區(qū)域內,先用直尺量取和標記一個單位壟長(150 cm),再在其壟上用鐵鍬反復翻土和尋找剩余的玉米粒,將播種與剩余玉米粒數的差值記為該單位壟長取食玉米種子粒數,之后將測量過的壟做好標記,避免重復測量。7 d內按此方法共計測量36次。取食效率、日取食玉米種子質量和攝入能量的估算方法如下。
取食效率(%)=A1/A2×100(其中,A1為單位壟長取食玉米種子粒數,A2為單位壟長播種機播種粒數,計算36次平均值并記為取食效率)。
日均取食玉米種子質量(kg)=(L1/L2)×η× 0.4× 10-3,其中,L1為日均取食壟的總長度(×102cm),L2為播種株距(cm),η為取食效率(%),0.4為平均每粒種子質量約為0.4 g。
攝入能量(kJ/d)=m×q,其中,m為干玉米種子質量(kg/d),q為干玉米種子熱值(kJ/kg)。用氧弾熱量計(GR-3500型,廣東)測定干玉米種子熱值。
1.2.2 籠養(yǎng)繁殖鶴對攝入總能量
籠養(yǎng)繁殖鶴對攝入總能量的實驗從2016年5月12日清晨開始至5月19日清晨結束,共記7 d。選擇10對籠養(yǎng)丹頂鶴繁殖鶴對(食物是鯽魚和玉米,鯽魚定時8:00—16:00足量,玉米全天足量),測定每對日平均攝食玉米和鯽魚的攝食量,用氧弾熱量計(GR-3500型,廣東)測定食物熱值和計算攝入總能量。測量和計算方法如下:
攝食量(kg/d)=投食量(kg/d)-剩余食物量(kg/d)
干物質(kg)=物質總質量(kg)-物質中水質量(kg)
攝入能(kJ/d)=攝入干食物質量(kg/d)×干食物熱值(kJ/kg)
攝入總能量(kJ/d)=攝入鯽魚能量(kJ/d)+攝入玉米能量(kJ/d)
1.2.3 卵殘殼厚度、密度和鈣元素含量的檢測和比較
卵殘殼取樣和處理方法:散放繁殖丹頂鶴,首次產卵和再次產卵的取樣方法相同。先用毛刷將收集的首次產的2枚卵的殘殼做表面清潔處理后,在2枚卵殘殼上選擇和截取包括卵殼上部、中部和下部各3塊面積相接近的樣品,共12塊,1塊/樣本數,之后,再將收集的再次產的2枚卵殘殼做上述相同處理,即樣本數n首次產卵=n再次產卵=12;籠養(yǎng)繁殖丹頂鶴,取12枚籠養(yǎng)鶴卵卵殘殼,其中每枚卵殘殼分別取其上部、中部和下部各1塊面積相接近的樣品,并將這3塊樣品測量值的平均值記為該枚卵的參數值,1枚卵/樣本數,即樣本數n籠養(yǎng)鶴卵=12。采用電子千分尺對卵殼厚度進行測量(mm)。根據公式ρ=m/v計算卵殼密度(電子天平稱量卵殼樣品質量(g)和排水法求得卵殼樣品的體積(cm3)。利用XPS光電子能譜儀檢測卵殼鈣元素含量(ppm)。
1.3 統(tǒng)計分析
利用SPSS 16.0進行統(tǒng)計分析。散放與籠養(yǎng)丹頂鶴繁殖鶴對攝食量和攝入能量的比較均采用獨立樣本t-檢驗(independent-samples)。散放繁殖丹頂鶴首次和再次產卵與籠養(yǎng)繁殖丹頂鶴的卵殘殼物理指標的比較采用單因素方差分析(one-way ANOVA)。文中數據均以平均值±標準誤(mean±SE)表示,P<0.05為差異顯著,P<0.01為差異極顯著。
2 結果
2.1 取食行為特征與取食效率
行為特征:丹頂鶴用喙(微張)插入壟上土壤中深度約10—15 cm后向兩側(雄鶴)或后方(雌鶴)掘土,壟上每次掘土間距為2—3 cm,掘土頻率為30—50次/min(包含吞咽食物時間),當玉米種子(橙黃色)和地下土(黑色)一同被掘出地表后,啄食玉米種子,之后繼續(xù)沿著壟的方向直行取食。每次取食持續(xù)時間和停歇時間都具隨機性。無干擾時可連續(xù)性取食距離可達幾米至十幾米不等,每次停歇(抬頭觀望和時間較短)后一般會換壟和開始反向取食。雌雄丹頂鶴自行取食,相距最近時可達1—2 m和最遠相距約50 m左右。經測算,取食玉米種子效率為91.22%。另外發(fā)現,丹頂鶴取食區(qū)域都是在農田內地勢相對低洼或地表土層較濕潤的地段內進行,對于取食過的區(qū)域未發(fā)現再次取食。觀測期間,這對丹頂鶴除每日會有幾次到農田邊上的水域飲水和短暫的停留外,白天大部分時間都在取食玉米種子。
2.2 攝入能量
7 d內,這對散放繁殖丹頂鶴日取食玉米質量(0.32±0.03)kg為籠養(yǎng)繁殖丹頂鶴鶴對日攝食玉米質量(0.08±0.02)kg的3.94倍(t=105.018,df=14,P=0.000)(圖1)。7 d內,其日取食玉米攝入的能量(4 324.98±398.90)kJ是籠養(yǎng)繁殖鶴對攝入玉米能量(1 103.31±188.26)kJ的3.92倍,但僅占籠養(yǎng)鶴對攝入總能量(6 384.73±266.89)kJ的67.74%(t=1 032.868,df=14,P=0.000;t=-56.461,df=14,P=0.000)(圖2)。
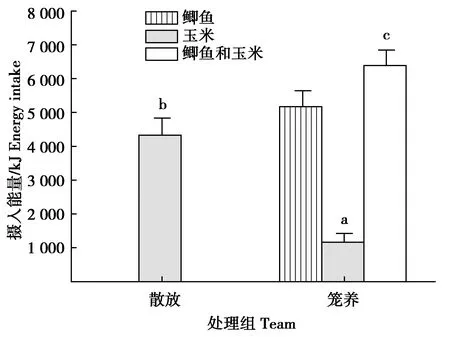
圖2 散放與籠養(yǎng)繁殖丹頂鶴攝入能量的比較Fig.2 Comparison of energy intake between released and caged red-crowned cranes pair in breeding period
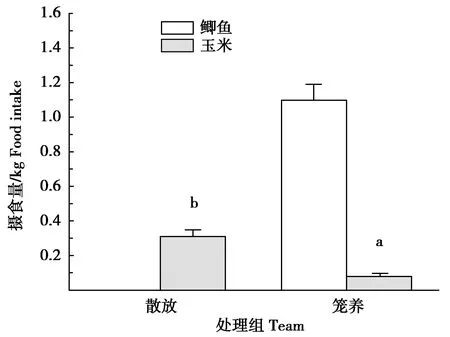
圖1 散放與籠養(yǎng)繁殖丹頂鶴攝食量的比較Fig.1 Comparison of food intake between released and caged red-crowned cranes pair in their breeding period
2.3 卵殼厚度、密度和鈣元素含量
卵殼厚度,首次產卵顯著薄于再次產卵和籠養(yǎng)鶴卵(F(2,33)=11.215,P=0.002),僅為再次產卵和籠養(yǎng)鶴卵的75%和70%,但再次產卵與籠養(yǎng)鶴卵組間差異不顯著(P>0.05)。卵殼密度,首次產卵顯著小于再次產卵和籠養(yǎng)鶴卵(F(2,33)=5.849,P=0.010),僅為再次產卵和籠養(yǎng)鶴卵的73.33%和88.00%,但首次產卵與籠養(yǎng)鶴卵組間差異不顯著(P>0.05)。卵殼中鈣元素含量,首次產卵最少和再次產卵最多,組間差異均極顯著(F(2,33)=28.629,P=0.000)(表1)。
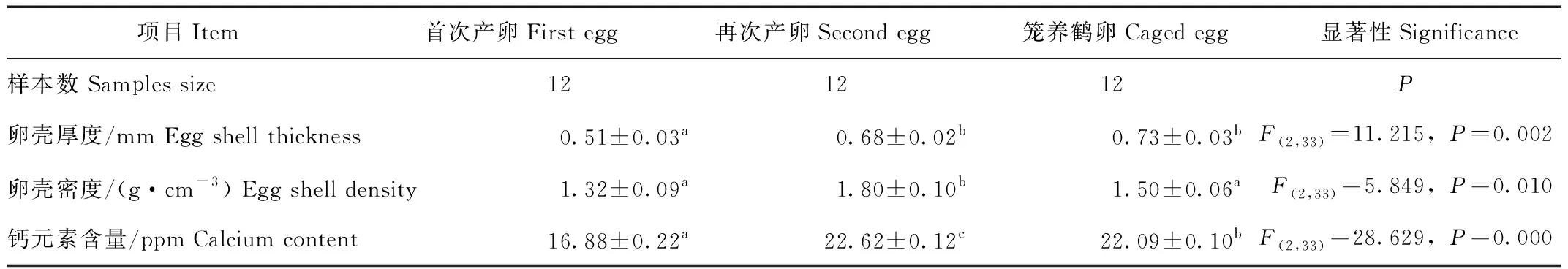
表1 散放與籠養(yǎng)丹頂鶴卵殼物理參數的比較Tab.1 Comparison physical parameters of egg shells between released and caged red-crowned cranes
3 討論
3.1 取食玉米種子的行為發(fā)生
鳥類不同的覓食對策不僅能夠反映出鳥類與其特定生境之間的契合,還可以呈現出鳥類對環(huán)境的適應模式和生理能力[13]。鳥類能夠充分利用自身特有的物種進化優(yōu)勢和選擇最適的能量收支對策來適應微生境變化[14-15]。理論上,扎龍濕地有著豐富的自然資源和物種多樣性,自然生境中丹頂鶴可獲得的食物種類和數量都應該優(yōu)于農田內取食玉米種子。那么,這對散放繁殖丹頂鶴選擇滯留農田和適應春耕期農田微生境變化的分析如下:首先,丹頂鶴具備用喙(長約15 cm)取食玉米種子的生理基礎(喙、頸、腿3長的生物學特點)。散放繁殖丹頂鶴,在取食玉米種子期間,不僅成功生存,還筑巢和產卵2枚,這說明丹頂鶴的這種取食對策能夠滿足其攝入能量需求。其次,與其繁殖習性(除了自然環(huán)境發(fā)生較大變化時繁殖丹頂鶴各年間的巢址都會相對固定[16])有關。據調查,此前連續(xù)3年,這對散放繁殖丹頂鶴都在該農田附近區(qū)域內進行筑巢和繁殖。最后,應該與鳥類間繁殖領域的競爭壓力有關。繁殖季節(jié),隨著時間的推移,重新選擇巢址和建立繁殖領域,理論上可能要同時應對種內甚至種間其他鳥類繁殖領域間逐漸升級的競爭壓力。例如,保護區(qū)內觀察發(fā)現,僅2對(2只/對)繁殖的同種的鷗(種內互助),采用交替(掠飛和休息)不間斷(掠飛)地伴有持續(xù)鳴叫聲的低空(從緊貼丹頂鶴頭部兩側或后方)急速反復穿插掠飛方法,丹頂鶴頻繁縮頭(每只/每次掠飛丹頂鶴都會縮頭一次)而又無可奈何,僅約30 min即可將1對繁殖丹頂鶴驅離其家域地帶(激怒反應)。所以,本研究認為丹頂鶴具備用喙掘土取食的生理基礎,掘土取食能夠滿足其攝入能量需求,固定領域內繁殖的生物學習性和繁殖季節(jié)存在繁殖領域的較大競爭壓力,這些因素的交互作用和影響導致這對散放繁殖丹頂鶴選擇滯留農田,并通過掘土取食來適應農田春耕和播種的微生境變化。
適宜覓食生境對鳥類的生存或繁衍意義重大[1,17]。丹頂鶴的生境選擇具有明顯的多尺度利用特征[11],微生境特征的改變會影響丹頂鶴的生境選擇[1],并且丹頂鶴對于棲息生境的整體分布和繁殖微生境質量變化也具有一定的適應和應答能力[17]。春季,扎龍濕地自然生境中有野生或散放兩類繁殖丹頂鶴,卻僅有這對散放繁殖丹頂鶴在農田內掘土取食。這對散放繁殖丹頂鶴滯留農田和適應農田微生境變化(外因)應該不是為了生存,而是要在此地繁殖(內因),即掘土取食的覓食對策與其繁殖需求有關。這對散放繁殖丹頂鶴的繁殖期與農田春耕(翻耕和播種)期在起始時間上存在交集,此前連續(xù)3年這對散放繁殖丹頂鶴在該農田區(qū)域內繁殖,其繁殖期家域與該春耕農田在地理位置上有重疊。為了繁殖,這對散放繁殖丹頂鶴選擇滯留農田和適應農田春耕的微生境變化。農田內掘土取食應該不是最佳的覓食對策,而是應對繁殖微生境變化的適應對策,這很有可能會對其生存或繁殖產生影響。
3.2 影響3.2.1 生存
食物可獲得性決定和影響著動物個體的能量收支收益[1,6,8,13]。調查發(fā)現,這對散放繁殖丹頂鶴在春耕農田內把玉米種子作為主要食物來源,掘土取食,輕易、精準和高效地取食玉米種子,持續(xù)數日并成功生存。這表明:觀察發(fā)現取食區(qū)域都是地表土壤濕潤的地帶,因此不會造成其喙的損傷,并且它們已獲取了生存所需的能量。然而,結果發(fā)現這對散放繁殖丹頂鶴日取食玉米種子的能量僅占籠養(yǎng)鶴對攝入總能量的67.74%,這也意味著僅取食玉米種子并不能滿足它們的攝入能量需求,而且,本文認為存在喜鵲(Picapica)、環(huán)頸雉(Phasianuscolohicus)和鼠類等盜食農田土壤中玉米種子的可能性,所以丹頂鶴取食玉米種子真實值極有可能會低于本文的估算結果。楊志宏等[13]認為在分析動物的食物可獲得性時不能脫離其特定的生態(tài)位,既要考慮食物的質量、數量、營養(yǎng)和適口性等方面,還要考慮到食物的可攝食時間、動物處理食物的能力及其攝入能量需求等方面。丹頂鶴是雜食性鳥類[6],因此不排除這對散放繁殖丹頂鶴在取食玉米種子的同時,還應該取食了土壤動物(如蚯蚓等)、青蛙、老鼠以及植物根莖等其他食物。總之,本文認為雖然此間這對散放繁殖丹頂鶴通過農田內的掘土取食獲得了維持生存所需的能量,但是也存在半饑餓或營養(yǎng)不良狀態(tài)的可能性。
3.2.2 繁殖
食物可獲得性不僅與環(huán)境變化密切相關,也會對鳥類的行為、覓食對策、能量收支收益、生存或繁殖等產生重要影響[18-20]。散放繁殖丹頂鶴2次繁殖,首次農田區(qū)域內產卵,尚未開始孵化,卵破碎和繁殖失敗,此后,離開農田區(qū)域再次產卵、孵化和成功繁殖。散放繁殖丹頂鶴通過掘土取食成功生存,且首次產卵(2枚),這表明此期間它們具有進行繁殖的生理基礎和能力以及具備了繁殖所需的基本物質條件。但是,首次繁殖微生境與再次繁殖的自然生境不同,且與此前3年繁殖的休耕農田微生境也不同,農田復耕使其繁殖家域的微生境發(fā)生了大的變化,選擇滯留農田(繼續(xù)占領繁殖家域)和掘土取食(覓食對策)是其生態(tài)適應對策,可獲得食物的種類和數量均隨之發(fā)生較大變化。結合此前3年這對繁殖丹頂鶴的繁殖記錄(均成功繁育后代)和這2次繁殖結果,本文認為首次繁殖失敗不應該與其自身的繁殖能力有關,而應該與其2次繁殖所處的微生境差異、食物可獲得性和覓食對策差異有關,存在攝入能量或營養(yǎng)不足的可能性,這很可能是導致其初次繁殖失敗的真正原因。
首次繁殖失敗是以卵破碎告終。卵破碎有兩種可能性:其一,農田復耕后,丹頂鶴巢址隱蔽效果較差,易受驚擾,不排除親代鶴爪意外踏碎卵的可能性(但是能使卵殼質量和硬度都達標的2枚卵都破碎的可能性很小);其二,2枚卵構成的營養(yǎng)元素不足和未達到孵化標準,親代棄卵后,卵破碎。無論前者還是后者,都與農田春耕有關,也與丹頂鶴選擇滯留農田和農田內掘土取食的覓食對策有關。兩種可能性,本文傾向于后者,理由是:一方面,數據顯示,這對散放繁殖丹頂鶴日取食玉米種子的質量是同期籠養(yǎng)繁殖丹頂鶴攝入玉米粒質量的3.92倍,占籠養(yǎng)繁殖丹頂鶴攝入總能量的67.74%,但它們攝入的玉米粒質量真實值很可能少于估算值。相關研究發(fā)現,春季繁殖期籠養(yǎng)繁殖丹頂鶴的攝入能量需求全年最低,攝食鯽魚(Carassiusauratus)和玉米的比例卻是全年最高,這表明繁殖季節(jié)繁殖丹頂鶴有著較低的攝入能量需求和更高的攝入營養(yǎng)需求[18]。鯽魚和玉米均為高能量食物,相比鯽魚中含有較多的蛋白質和鈣元素,玉米粒中含有較多的糖類物質和維生素。從籠養(yǎng)繁殖丹頂鶴攝食比例來看,繁殖季節(jié)繁殖丹頂鶴應該有著較高的攝入蛋白質和鈣元素等營養(yǎng)物質的攝入需求。雌鶴產卵2枚(約250 g/枚),蛋白質(卵清蛋白)和鈣元素(卵殼)等營養(yǎng)物質是構成卵的重要組成成分,雌鶴攝入足夠的營養(yǎng)物質是其生產2枚達標卵的物質基礎和前提。然而,這對散放繁殖丹頂鶴的攝食比例卻與籠養(yǎng)繁殖丹頂鶴相反(除主要取食玉米種子以外,可能隨機取食少量其他食物,蛋白質攝入量極有可能不足)。因此,本文認為這對散放繁殖丹頂鶴掘土取食期間存在營養(yǎng)不良和首次產卵未能達標的可能性。另一方面,數據顯示,首次產卵卵殼厚度、密度和鈣元素含量均顯著低于再次產卵,卵殼厚度和鈣元素含量同樣顯著低于籠養(yǎng)鶴卵,這就表明首次產卵卵殼極有可能未達標。綜上所述,無論后者卵殼的厚度和硬度未達標使其卵破碎,還是前者卵內蛋白質等營養(yǎng)成分不足親代丹頂鶴選擇棄卵,首次繁殖失敗以卵破碎告終,卵破碎又極有可能是其產卵前蛋白質或鈣元素等營養(yǎng)物質攝入不足所致,營養(yǎng)物質攝入不足與其掘土取食農田內玉米種子的覓食對策有關。所以,本文認為這對散放繁殖丹頂鶴農田內掘土取食期間攝入營養(yǎng)物質不足,導致其首次繁殖失敗,應該更具有可能性。
從生態(tài)學意義上來理解,掘土取食的行為發(fā)生應該是其應對繁殖家域內微生境改變的一種生態(tài)適應。這對散放繁殖丹頂鶴前后共筑巢和繁殖2次,首次巢址位點選擇在農田邊緣區(qū)域和繁殖失敗,再次的巢址位點已經遠離農田區(qū)域和繁殖成功,這可以理解為是其應對繁殖家域內微生境變化的兩種繁殖對策(適應與逃離)。
總之,這對散放繁殖丹頂鶴通過掘土取食農田玉米種子,成功生存,卻未能繁殖成功。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
