蘭州市北山不同林地春夏季土壤纖毛蟲群落特征
2020-07-31 09:36:08劉旻霞李全弟蔣曉軒夏素娟南笑寧
生態學報 2020年12期
關鍵詞:物種
劉旻霞, 李全弟, 蔣曉軒, 夏素娟, 南笑寧
西北師范大學地理與環境科學學院, 蘭州 730070
土壤作為生物圈能量交換與物質遷移的重要載體,承載了絕大部分生物的生命活動,是地球生態環境的主要構成部分[1]。土壤原生動物是土壤動物這一巨大生物群落的主要類群,在土壤生態系統的物質循環和能量流動方面扮演著十分重要的角色[2]。土壤纖毛蟲是土壤原生動物的三大組成類群之一,它比其他土壤生物具有更大的生物量和生產力,由于其與外界相接觸的原生質細胞膜比較薄,因此土壤纖毛蟲對外部環境的微小變化的響應非常敏捷,比細菌等原核生物對環境變化的響應程度更高;有大批的土壤纖毛蟲類群的分布是全球性的,這有利于在不同區域之間進行互相對比[3]。土壤纖毛蟲還具有個體較小、繁殖速度快、數量巨大及在惡劣環境條件下可以生存等的特點,其物種多樣性特征和群落結構的變化可以很好地反應出其生存地區環境情況,因此被當作十分重要的土壤環境狀況的指示生物。近年來,有利用底棲纖毛蟲群落的生物性狀分析和功能多樣性指數進行環境狀況評估[4],取得了良好的效果。而甘肅省對土壤纖毛蟲群落的研究多集中于高寒草甸地區放牧及施肥的研究,也有涉及到退耕還林與植被及土壤養分含量變化的一些研究[5-6],針對蘭州市北山不同植被類型的季節變化研究相對較少。
蘭州市是我國西部地區非常典型的生態環境脆弱區,具有黃土高原典型地貌特征,大部分土地覆蓋深厚黃土,溝谷地帶砂礫裸露[7]。長期以來受到自然條件和人為活動的雙重影響,土地承載力逐步下降,水分和養分的流失日益加重,嚴重制約了蘭州城市發展和社會經濟的提高[8]。為了緩解黃土高原地區日益加重的水土及養分流失狀況,我國于1999年提出了退耕還林還草政策。2000年,蘭州市生態造林工程全面開展,城市生態建設取得了明顯成效,植被綠化覆蓋率和土壤恢復得到有效改善。學者們對蘭州市兩山的研究也逐漸開展起來。史志熠[9]對兩山的植被恢復和重建情況做了調查,調查結果顯示人工造林和封山育林對兩山的綠化效果十分顯著。石善斌和席正英[10]對蘭州市兩山的地被植物做了調查,結果顯示蘭州市兩山地被植物具有資源豐富,但分布不均勻,垂直構成比較單調,樹種單一,灌木和草本缺乏自然配置等特點。為了使兩山的綠化更加科學和合理的進行,許宏剛等[11]研究了兩山植物的群落結構、優勢種構成、物種多樣性特性,探究適合在蘭州市南北兩山面山的特定立地條件下生長的植被種類。然而對蘭州市北山相關土壤纖毛蟲群落特征及與生態恢復的關系的研究卻仍舊缺乏。
本研究選擇蘭州市北山的羅九公路綠化區為研究對象,研究受人為影響較重的人工林與荒坡之間的土壤纖毛蟲群落特征、土壤環境因子差異、土壤纖毛蟲群落的分布與環境因子的關聯關系,以及季節變化對這些因素的影響。旨在正確評估蘭州市南北山的生態恢復情況;確定不同綠化植被對生態恢復影響的差別;以各樣地土壤纖毛蟲群落對環境因子狀況的響應,指示不同樣地的環境狀況,尋求對土壤恢復最有利的恢復樣點。本研究試圖回答以下科學問題:(1)土壤纖毛蟲群落出現差異的原因?(2)探究土壤纖毛蟲群落在不同季節的變化特征及影響因素。通過以上問題的研究可以為兩山綠化工程建設提供更加經濟有效的方法,為黃土高原干旱區退化生態系統恢復提供理論依據。
1 研究區域與方法
1.1 研究區概況
研究區位于蘭市北山羅九公路綠化工程區附近(圖1),地理坐標為36°03′N、103°40′E,海拔1780—1860 m。蘭州市居于內陸深處,溫濕氣流很難到達,雨水稀少,氣候干燥,土壤主要為灰鈣土。植被覆蓋度在20%—50%。主要植被有:側柏(Platycladusorientalis)、刺槐(Robiniapseudoacacia)、新疆楊(Populusalba)、檸條(Caraganakorshinskii)、檉柳(Tamarixchinensis)、紫穗槐(Amorphafruticosa)、沙棗(Elaeagnusangustifolia)及一些經濟果樹。
圖1 研究區位置圖Fig.1 Location map of study areaS1:樣區1 Sample1; S2:樣區2 Sample2
1.2 試驗設計與樣品采集
于2016年4月(春季)、7月(夏季)在2個大樣區(每個樣區約65萬m2)進行野外重復調查采樣,該研究樣區在最近幾年均引入了噴灌、滴灌,所以在設置的樣地和對照地內都能進行人工灌溉。每個樣區設置大小為30 m×30 m的樣地10個,2個大樣區共設置20個30 m×30 m的樣地(表1),同時的調查了樣地坡度、坡向、海拔、植被及土壤溫度等環境因子,同時用直徑為3.5 cm的土鉆,采用“梅花五點法”在每個樣地取0—10 cm土壤5份,每個季節取50份土樣,2季共采100份土樣。去除土壤表面的凋落物以及石塊后,立即裝入自封袋帶回實驗室做后續實驗。
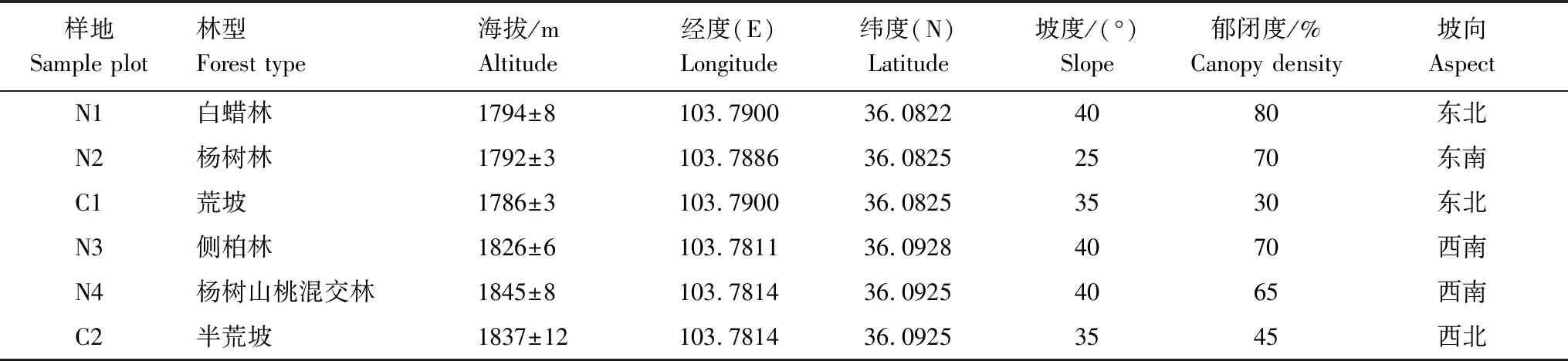
表1 樣地生境概況
1.3 室內處理及方法
1.3.1 土壤理化性質的測定
土壤全氮用半微量凱氏定氮法,土壤全磷用硫酸-高氯酸消煮法,土壤有機碳用硫酸消煮-重鉻酸鉀外加熱法,土壤pH值用水浸提電位法,土壤含水量用烘干法(重復3次),土壤溫度用土溫計測定。
1.3.2 土壤纖毛蟲的鑒定
從每份新鮮土樣中取適量等體積的3份至于培養皿中,采用“非淹沒培養皿法”(non-flooded petridish method)培養鑒定土壤纖毛蟲。即在加有土樣的培養皿中加入土壤浸出液,將土壤充分潤濕,但不被淹沒,在25℃的恒溫下培養土壤纖毛蟲[12-13],在培養后的第2、4、7、11、14、21、30天放置于光學顯微鏡下鑒定土壤纖毛蟲的物種,并記錄每個物種的個體數。土壤纖毛蟲的物種鑒定技術采用“活體觀察法”和“固定染色法”,固定染色技術主要依據Fernandez-Galiano等[14- 16]的蛋白銀染色法。物種鑒定依據文獻[17- 19]。采用Foissner計數法[20]對纖毛蟲進行培養計數,即將已風干30 g土壤樣品置于直徑為5 cm的培養皿中,加入蒸餾水使其略微高出土壤表面,25℃恒溫培養9 d,在第10天時使培養皿傾斜45度5 min,吸取土壤浸出液并用波恩液固定,取1 mL搖勻的上述液體于顯微鏡下計數。
1.4 數據處理與分析
1.4.1 多樣性指數計算
(1)多樣性指數(Shannon-Wiener)公式:
(1)
(2)均勻度指數(Pieluo)公式:
E=H′/lnS
(2)
(3)優勢度指數(Simpson)公式:
(3)
(4)豐富度指數(Margalef)公式:
(4)
式中,H為物種的多樣性指數;Pi為第i類群的個體數比,Pi=Ni/N,Ni為第i類群的個體數,N為總個體數;S為所有類群數。
C/P系數:腎形綱土壤纖毛蟲(r對策者)的物種數與異毛綱、旋毛綱和瓶纖綱土壤纖毛蟲(k對策者)物種數總和的比值。C/P系數的數值大小能在一定水平上體現出土壤纖毛蟲所生存的生境的環境惡劣狀況,在環境質量相對較好的土壤生境中C /P≤1,而在無法衡量的環境質量較差的土壤生境中C/P > 1[21]。
1.4.2 相似性指數計算
對各樣點土壤纖毛蟲群落特征的相似性研究用Jaccard相似性指數。公式:q=c/(a+b-c)式中,c用來表示兩個群落中的共有物種數,a和b分別用來表示兩個群落各自獨立的物種數。當q值在0.00—0.25范圍時,為極不相似;當q值在0.25—0.50范圍時,為中等不相似;當q值在0.50—0.75范圍時,為中等相似;當q值在0.75—1.00范圍時,為極相似[22]。
1.4.3 數據統計與分析
在Excel 2010中進行數據的整理與計算,在SPSS 18.0中進行環境因子與土壤纖毛蟲的相關分析與單因素方差分析,運用CANOCO 4.5軟件對土壤纖毛蟲與環境因子進行冗余分析(RDA),運用Sigmaplot 12.0軟件作圖。
2 結果與分析
2.1 不同樣地土壤環境因子的季節變化
圖2顯示,土壤全氮、全磷、有機碳含量在荒坡地顯著低于人工林樣地,而在人工林樣地中,側柏純林又明顯低于其他人工林。春季不同人工林樣地間、人工林與荒坡間的土壤全磷量有顯著差異(P<0.05)。夏季白蠟純磷、側柏純林與荒坡、半荒坡間的全磷含量有顯著差異(P<0.05)。土壤有機碳和土壤全氮在不同樣地之間變化趨勢相同,楊樹純林、楊樹-山桃混交林及半荒坡的土壤有機碳和全氮含量較高,而側柏純林和荒坡較低。夏季土壤溫度和土壤含水量高于春季,但是土壤pH高于春季。夏季荒坡的土壤溫度最低,其次是半荒坡,春季荒坡的土壤溫度在所有樣點中最高。土壤含水量人工林高于荒坡對照。春季的側柏純林、楊樹-山桃林和半荒坡之間土壤含水量差異較大。夏季的白蠟樹林與其他樣點呈極顯著差異(P<0.01),荒坡與側柏純林含水量的差異性顯著(P<0.05),而與其他樣點的差異性達到極顯著(P<0.01)。
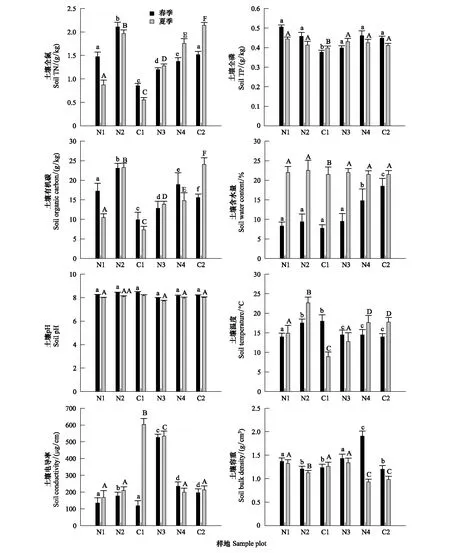
圖2 不同樣地土壤環境因子的季節變化Fig.2 Seasonal variation of soil environmental factors in different plots不同小寫字母表示春季不同樣地間差異顯著(P<0.05);不同大寫字母表示夏季不同樣地間差異顯著(P<0.05)
2.2 土壤纖毛蟲群落結構
蘭州市北山共鑒定出土壤纖毛蟲10綱21目39科62屬157種(表2)。其中旋毛綱有5目12科25屬48種,占總種類數30.57%;裂口綱有1目4科10屬28種,占總種類數17.83%;腎形綱2目3科5屬21種,占總種類數13.38%;寡膜綱4目6科6屬19種,占總種類數12.1%。春季有土壤纖毛蟲10綱21目34科44屬80種。旋毛綱有5目10科16屬24種,占總種類數30.00%;裂口綱有1目4科7屬14種,占總種類數17.50%;腎形綱2目2科3屬11種,占總種類數13.75%;寡膜綱4目5科5屬8種,占總種類數10.00%。夏季有土壤纖毛蟲10綱21目38科54屬104種。旋毛綱有5目11科20屬33種,占總種類數31.73%;裂口綱有1目4科8屬18種,占總種類數17.31%;腎形綱2目3科4屬13種,占總種類數12.50%;寡膜綱4目6科6屬12種,占總種類數11.54%。夏季的物種數明顯多于春季,尤其是前管目、嗜污目及固著目的數量大大增加,由罕見類變為常見類。紡錘全列蟲(Holostichakessleri)春季在人工林N1、N2、N3及N4為常見種,而夏季只有N3中作為偶見種出現,其他樣地均沒有;扭曲管葉蟲(T.sigmoides)、巴維利亞斜管蟲(C.barvariensis)、鉤刺斜管蟲(Chilodonellauncinata)在夏季的N1、N2;N1、N2、N3;N4中為常見種,在春季的人工林中沒有出現。大喙纖蟲(Loxodesmagnus)、速體蟲(Tachysomapellionellum)及似織毛蟲(Histriculussimiles)春季分別在N1、N2、N4;N1、N3; N2、N3、N4中是常見種,在夏季的人工林中數量減少,變為偶見種。
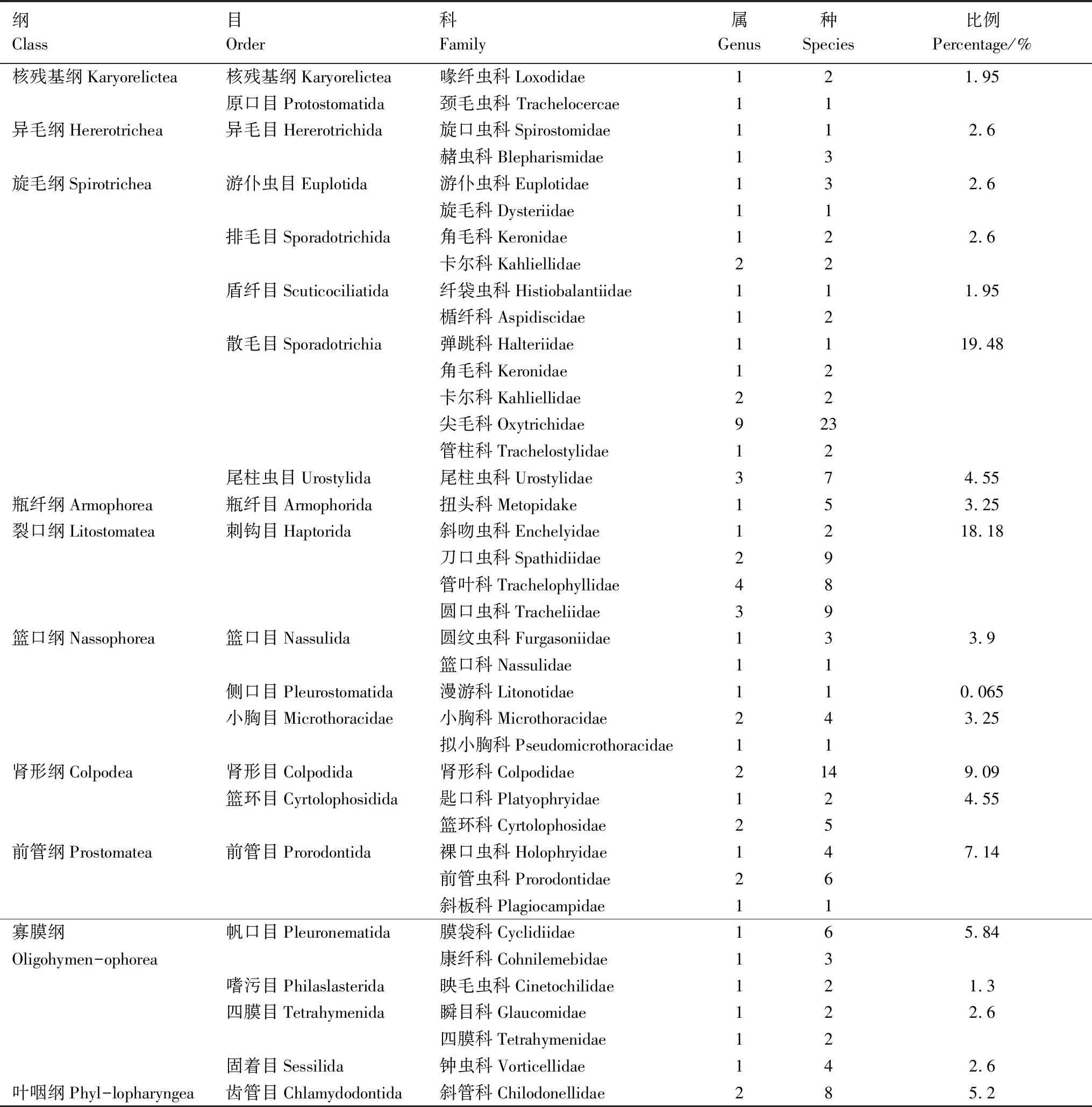
表2 土壤纖毛蟲群落結構
2.3 土壤纖毛蟲多樣性和C/P系數變化
圖3顯示,纖毛蟲物種數、豐度、香農指數及Margalef豐富度指數均是夏季高于春季;均勻度指數及優勢度指數則是春季高于夏季。在不同樣地中,人工林的土壤纖毛蟲物種數、豐度及多樣性指數均高于荒坡對照樣點;而均勻度指數和優勢度指數則是荒坡高于人工林。春季楊樹林與對照樣地荒坡的物種分布存在極顯著差異(P<0.01);夏季不同人工林之間、人工林和荒坡對照間有極顯著差異(P<0.01);可見人工林對土壤恢復狀況良好,使得土壤纖毛蟲物種數分布相比荒坡和半荒坡有明顯的優勢。春季白蠟純林和側柏純林的土壤纖毛蟲豐度最高,夏季楊樹純林和側柏純林的豐度最高。Margalef豐富度指數人工林的土壤纖毛蟲物種多樣性指數明顯高于對照樣地荒坡和半荒坡。
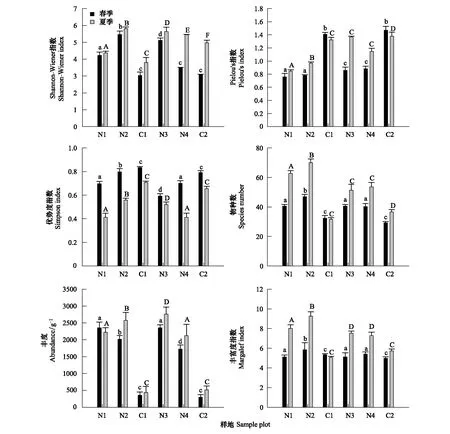
圖3 不同樣地土壤纖毛蟲多樣性Fig.3 Diversity of soil ciliates in different plots不同小寫字母表示春季不同樣地間差異顯著(P<0.05);不同大寫字母表示夏季不同樣地間差異顯著(P<0.05)
各樣地的土壤纖毛蟲群落C/P系數春季(0.38)>夏季(0.33)。不同樣地土壤纖毛蟲群落C/P系數隨季節變化差異較大,其C/P系數的動態變化為:春季蒿子-灌叢半荒坡(0.55)>楊樹-山桃林(0.53)>側柏-檸條林(0.45)>荒坡(0.44)>白蠟樹林(0.43)>楊樹林(0.36),夏季蒿子-灌叢半荒坡(0.64)>側柏-檸條林(0.46)>白蠟樹林(0.41)>楊樹林(0.37)>楊樹-山桃林(0.36)>荒坡(0.33)。
2.4 土壤纖毛蟲群落相似性特征
表3顯示,春季不同樣地間的相似性系數范圍為0.39—0.58,在中等相似和中等不相似之間,人工林與荒坡之間的相似性除白蠟林與荒坡、楊樹-山桃林以及楊樹與側柏林、混交林為中等相似外,其他均為中等不相似;夏季不同樣地間的相似性系數在0.40—0.79之間,在中等不相似到極相似之間,人工林與荒坡之間表現為中等不相似,其他樣點之間均表現為中等相似,白蠟樹林與楊樹林表現為極相似。
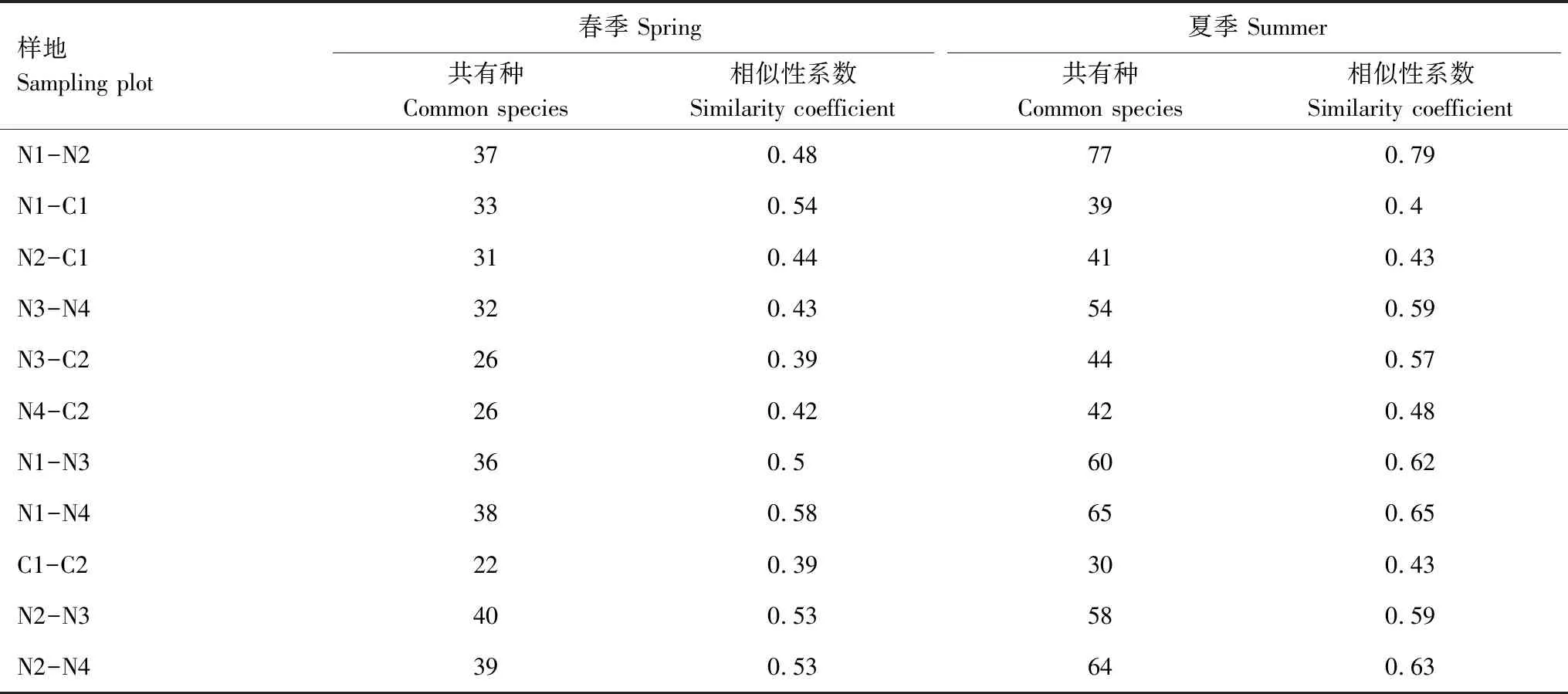
表3 土壤纖毛蟲群落相似性
圖4表明,春季不同樣地土壤纖毛蟲種類組成在歐式距離1.0處楊樹純林和側柏純林首先聚為一類,在歐氏距離6.0處與楊樹-山桃混交林聚為一類,在歐氏距離13.0處與白蠟純林聚為一類,荒坡與半荒坡在歐氏距離3.0處聚為一類后與人工林在25.0處聚為一類。圖4顯示,夏季側柏純林和楊樹-山桃林混交林在歐氏距離2.0處聚為一類、荒坡和半荒坡在歐氏距離7.0處聚為一類,接著在歐氏距離10.0處聚為一類,最后與在歐氏距離1.5處聚為一類后與白蠟樹林和楊樹林在歐氏距離25.0處聚為一類。表明荒坡的生態恢復較差,半荒坡的土壤恢復狀況較荒坡好,其次是側柏林和楊樹-山桃混交林,白蠟樹林和楊樹林的土壤恢復情況最好。
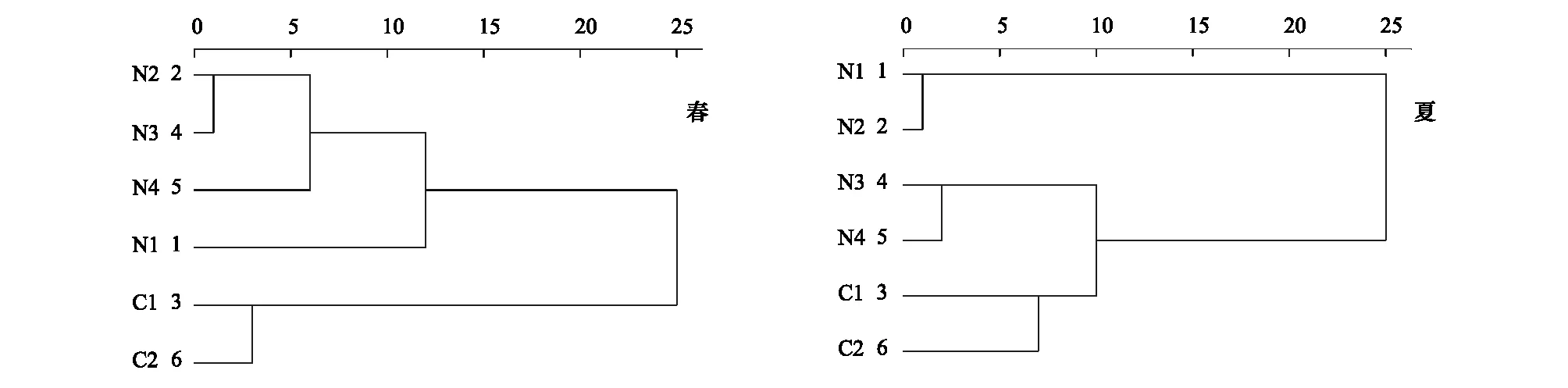
圖4 土壤纖毛蟲的聚類分析Fig.4 Cluster analysis of soil ciliates
2.5 土壤理化性質與土壤纖毛蟲的相關關系
本研究采用冗余分析(RDA)方法,圖中實線箭頭代表土壤纖毛蟲群落,虛線箭頭代表各環境因子;實線與虛線之間的夾角代表土壤纖毛蟲與各環境因子之間的相關性。圖5顯示,春季大多數土壤纖毛蟲與土壤電導率、土壤溫度、有機碳等呈正相關關系,與土壤含水量呈負相關關系,且大多是纖毛蟲都分布在人工林樣地,荒坡和半荒坡對照樣地分布數量較少。斜板科和斜吻蟲科土壤纖毛蟲受環境因子的影響最小。圖5表明,夏季土壤纖毛蟲土壤全氮、有機碳及電導率正相關,與其他環境因子呈負相關關系,除尾柱蟲科、籃口科及裸口蟲科的少數種外,極大多數土壤纖毛蟲同樣分布在人工林樣地。這些均表明經過多年的人工林培育,其土壤環境明顯優于荒坡和半荒坡。表4為各物種RDA第一、二排序軸與環境因子的相關系數,由表4可以看出,環境因子對土壤纖毛蟲的前2軸的累積解釋量占特征值的總和分別為62.90%和74.70%,表明排序軸可有效反映蘭州北山土壤及不同樣地環境因子的梯度變化特征。
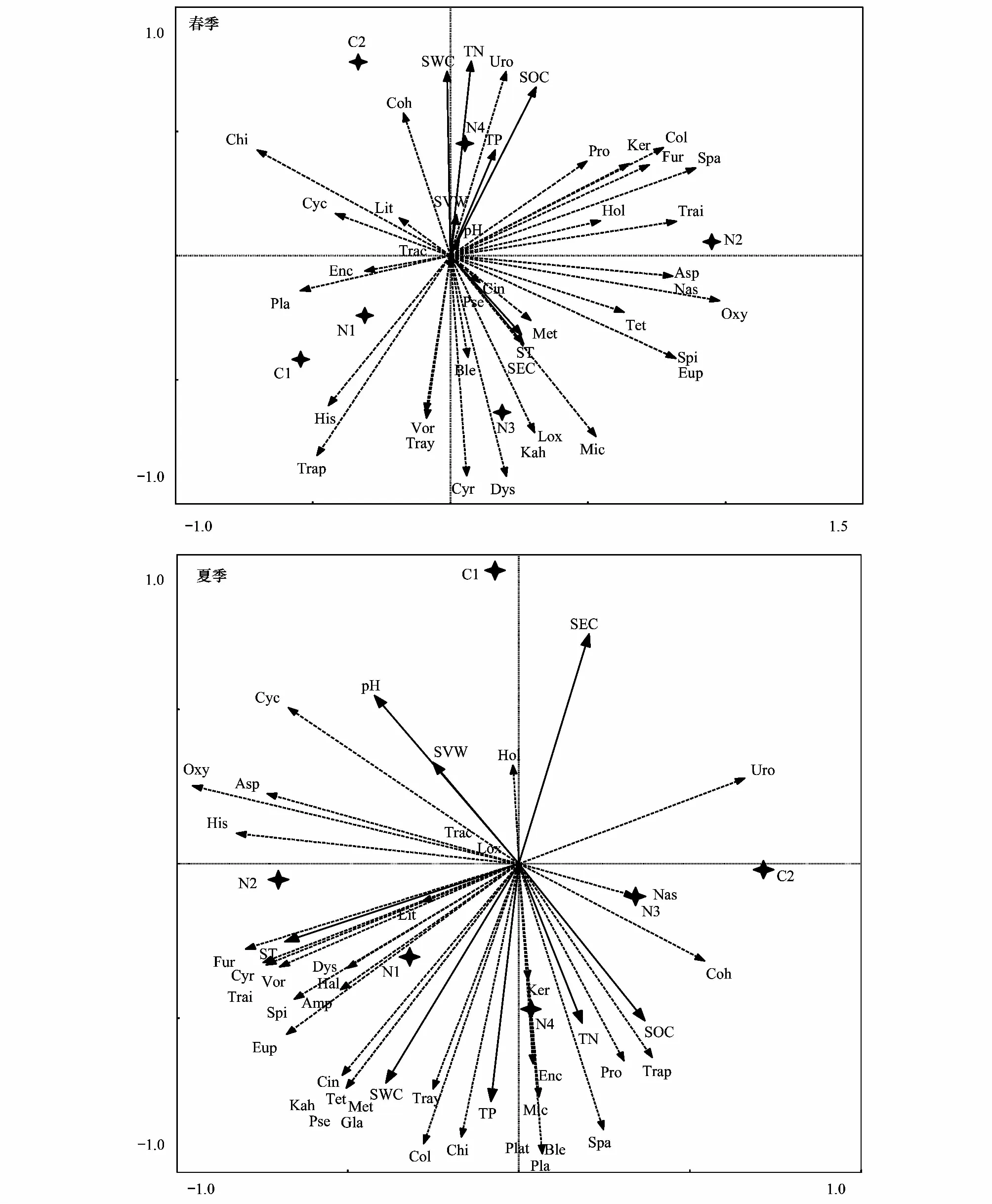
圖5 土壤纖毛蟲與環境因子的冗余分析Fig.5 Redundancy analysis of soil ciliates and environmental factorsLox:喙纖蟲科Loxodidae;Trac:頸毛蟲科 Trachelocercae;Spi:旋口蟲科Spirostomidae;Ble:赭蟲科 Blepharismidae;Eup:游仆蟲科Euplotidae Ehrenberg;Dys:旋毛科Dysteriidae Claparéde;Ker:角毛科Keronidae;Kah:卡爾科Kahliellidae;Amp:小雙蟲科Amphisiellidae;Hal:彈跳科Halteriidae;Asp:楯纖科Aspidiscidae; His:纖袋蟲科Histiobalantiidae;Oxy:尖毛科Oxytrichidae;Tray:管柱科Trachelostylidae;Uro:尾柱蟲科Urostylidae;Met:扭頭科Metopidake;Enc:斜吻蟲科Enchelyidae;Spa:刀口蟲科Spathidiidae;Trap:管葉科Trachelophyllidae;Trai:圓口蟲科Tracheliidae;Fur:圓紋蟲科Furgasoniidae;Nas:籃口科Nassulidae;Lit:漫游科Litonotidae;Mic:小胸科Microthoracidae;Pse:擬小胸科Pseudomicrothoracidae;Col:腎形科Colpodidae;Plat:匙口科Platyophryidae;Cyr:籃環科Cyrtolophosidae;Hol:裸口蟲科Holophryidae;Pro:前管蟲科Prorodontidae;Pla:斜板科Plagiocampidae;Cyc:膜袋科Cyclidiidae;Coh:康纖科Cohnilemebidae;Cin:映毛蟲科Cinetochilidae;Gla:瞬目科Glaucomidae;Tet:四膜科Tetrahymenidae;Vor:鐘蟲科Vorticellidae;Chi :斜管科Chilodonellidae;TN- Total nitrogen:土壤全氮;TP- Total phosphorus:土壤全磷;SOC- Soil organic carbon:土壤有機碳;SWC:土壤含水量;SEC:土壤電導率;SVW:土壤容重;pH:pH值;ST:土壤溫度
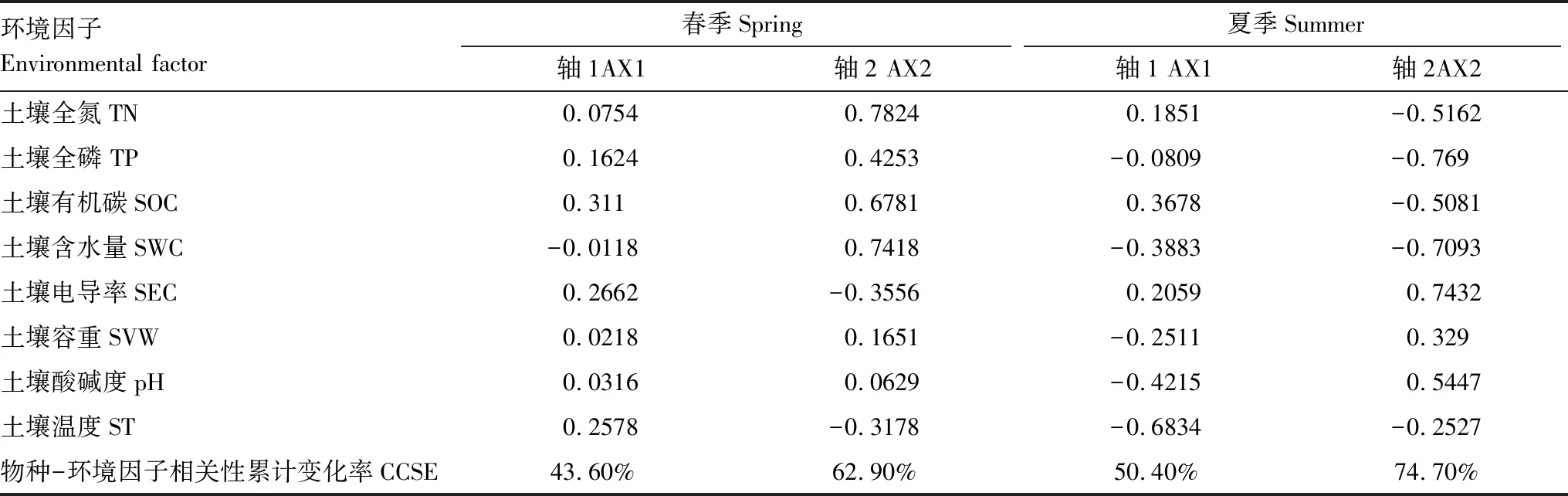
表4 排序軸與土壤因子的RDA分析
CCSE: 物種-環境因子相關累積變化率 Cumulative change rate of correlation between species and environmental factors;TN:土壤全氮 Total nitrogen;TP:土壤全磷 Total phosphorus;SOC:土壤有機碳 Soil organic carbon;SWC:土壤含水量 Soil water content;SEC:土壤電導率 Soil electrical conductivity;SVW:土壤容重 Soil volume-weight; pH: 土壤酸堿度 Soil pH;ST:土壤溫度 Soil temperature
3 討論
蘭州市屬于干旱區,全年降雨較少,季節溫度變化較大。而溫度是調節陸地生態系統生物化學過程的重要環境因子,碳和氮循環的主要過程,包括植物光合與呼吸作用、凋落物的積累與分解作用、土壤呼吸與 CO2釋放,土壤動物的取食活動與代謝等都受土壤溫度的調節[23]。土壤水分是土壤原生動物生存、繁殖和分布的主要限制因子[24],大多數土壤生物生活于土壤孔隙中,其活動依賴于水分的可獲得性[25-26]。研究區不同樣地間的土壤理化因子差異明顯(圖2),表現為人工林樣地的養分含量普遍高于荒坡對照樣地,這表明蘭州市經過多年的退耕還林改造,土壤環境條件已經得到了大大的改善。而季節變化中,夏季降水偏多,溫度高,導致土壤含水量較多、土壤溫度較高,但土壤pH減小,這與土壤含水量增大,土壤偏酸性的研究結果一致[27]。
北山是蘭州市兩山綠化的重要地區,人為的有利干擾(比如植樹造林)對土壤纖毛蟲群落分布有較顯著影響,受人工長期對環境的調控使其土壤環境條件更適宜土壤纖毛蟲生存。本次研究中在該樣區共計鑒定出土壤纖毛蟲10綱21目39科62屬157種(表2),與甘南瑪曲縣的沼澤化草甸[4]8綱17目30 科39屬 105種相比,明顯可以看出蘭州市綠化工程區的土壤纖毛蟲物種數較高,這說明該綠化工程區的環境有一定的獨特性,且土壤改良也在向好的方向轉變。春夏兩季土壤纖毛蟲的優勢種均為膨脹腎形蟲和盤狀腎形蟲,這與腎形目對較貧瘠環境的適應性有關,腎形目能在人為干擾較多、植被覆蓋率較低的土壤環境中占優勢,說明蘭州市南北兩山經過多年的植被恢復與土壤環境的改善,仍然沒有恢復到理想狀態,生態環境的改善工作依然任重而道遠。另有研究表明,造成原生動物的種群出現差異的主要原因是溫度和降水[28]。當地面溫度在14.3到24.3℃之間時是適宜土壤動物生存的最佳溫度[29]。寧應之[5]等的研究結果表明,土壤原生動物的豐度一年中有1個或2個高峰,大都出現在夏季。本研究中土壤纖毛蟲的豐度、物種數及多樣性指數均是夏季高于春季(圖3),主要原因是本研究地的夏季土壤環境比較適宜大多數土壤纖毛蟲的生存,因此土壤纖毛蟲的物種數量較大。另一方面,植物也是生態系統初級生產者,植被的存在形式以及其發育狀況決定著生態系統的組成成分和結構特征[30]。植被恢復能促進該地區棲息和繁衍的土壤生物種類增多,土壤動物的數量也會明顯增加[31]。本研究中人工林樣點的土壤纖毛蟲的物種數、豐度和多樣性均高于荒坡、半荒坡對照樣地,這與對退耕還林區土壤動物的動態變化的研究結果一致[32]。荒坡植被覆蓋率低(表1),惡劣環境的沖擊影響較重,這導致對環境條件需求較高的土壤纖毛蟲類群無法生存或大量繁殖,而人工林樣地中受人們維護的影響,植被和土壤逐漸恢復,能夠為土壤纖毛蟲直接或間接的食物來源,提供生存繁殖所需要的營養條件,因此土壤纖毛蟲的多樣性、豐度及物種數較荒坡高。異毛綱、旋毛綱、瓶纖綱土壤纖毛蟲生存為K-對策,腎形綱土壤纖毛蟲生存為R-對策。C/P系數越大,則說明腎形綱在該生境中越占據較高的優勢,即生存為R-對策占據優勢,該生境環境質量越差[33]。蘭州市羅九公路綠化工程區各樣點中蒿子-灌叢半荒坡的C/P系數在春夏兩季均較高,腎形綱的種類較多,這與腎形綱對較貧瘠環境的適應性有關,腎形綱能在人為破壞較重、植被覆蓋率較低的土壤環境中占優勢。這說明蒿子灌叢半荒坡的土壤環境質量較差,楊樹林、側柏林等生境的土壤質量恢復程度較高。兩季人工林樣點的土壤纖毛蟲多樣性指數較高均是楊樹林和側柏林,這也表明了楊樹林和側柏林對土壤的恢復效果較好。
Jaccard相似性系數評價不同生境之間的相似性與差異性[22]。本研究顯示(表3),春季不同樣點間的相似性系數的范圍為0.39—0.58,人工林樣點與對照點的相似性系數在0.39—0.54之間,相似性系數最高的為白蠟純林和楊樹-山桃混交林(0.58),說明春季土壤纖毛蟲群落受各樣點的影響差異較大,白蠟林和楊樹-山桃林的植被覆蓋度較高,土壤表面凋落層較厚,因而土壤環境差異小;夏季不同樣點間的相似性系數在0.40—0.79之間,人工林樣點與對照樣點的相似性系數范圍為0.40—0.57,人工林樣點之間的相似性系數較高,說明夏季土壤纖毛蟲群落受人工林的影響差異不大。馬正學等[34]對甘肅太統-崆峒山自然保護區的研究中,油松林、沙棘灌叢、薔薇灌叢和荒坡之間的相似性系數在0.275—0.292之間,遠低于蘭州市北山綠化區白蠟樹林、楊樹林和荒坡之間的相似性系數,表明崆峒山森林自然保護區的土壤環境質量明顯較高,與對照荒坡之間差異較大。這也同樣證明了蘭州市綠化工程區的人工林樣地的土壤恢復程度還未達到最好的程度。聚類分析結果(圖4)表明,兩季中均是人工林之間先聚為一類,最后與對照樣地聚為一類。說明人工林的土壤質量和荒坡還是有明顯差異。不同人工林之間的土壤纖毛蟲群落存在一定的差異性,這是因為人工林之間的樹種、蓋度以及坡向的差異(表1),引起不同季節的溫度、光照、土壤含水量及養分等的差異,這與劉旻霞等[35]在甘南高寒草甸坡向梯度的研究結果一致。
土壤動物與其生存的環境之間有著復雜的相互關系,動物能夠過通過其生命活動過程促進營養元素在凋落物和土壤系統中的循環,尤其在土壤營養元素特別是氮、磷元素的循環中起著重要作用[36-37]。張雪萍等[38]的研究結果顯示,土壤動物的種類、數量和生物量與土壤有機質、全氮和全磷的含量正相關。在本研究中,春季有機碳、土壤溫度及電導率等與土壤纖毛蟲正相關,與其他環境因子負相關(圖5);夏季是動植物的快速生長繁殖的時期,對土壤營養元素的競爭激烈,研究顯示土壤纖毛蟲物種分布與土壤有機碳和電導率等正相關,與其他環境因子呈負相關關系(圖5)。縱觀春夏兩季,土壤有機碳對土壤纖毛蟲群落的影響較大。主要原因是土壤有機碳是指示土壤健康的關鍵指標,其不僅是植物養分循環的中心,而且影響水分關系和土壤被侵蝕的潛力,是表征土壤結構的關鍵因子[38];另一方面,是因為研究區不同人工林之間的植被種類不同,土壤表面上的凋落物也不同,不同凋落物分解的難易程度就會出現差別[39],加之春夏兩季的土溫及含水量差異較大,其纖毛蟲的物種分布就不同。
綜上所述,蘭州市北山不同植被類型下土壤纖毛蟲的分布不同,且這些分布與環境因子的季節變化各有不同,最終使得土壤纖毛蟲群落出現差異。探究土壤纖毛蟲群落在不同季節、不同生境的變化特征及影響因素,可為黃土高原干旱區退化生態系統恢復提供理論依據。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
