小麥-華山新麥草異附加系的細胞遺傳學和分子標記輔助鑒定
2020-07-31 10:41:02張德時王斯文王長有王艷珍陳春環吉萬全
麥類作物學報 2020年1期
張德時,王斯文,王長有,2,王艷珍,陳春環,吉萬全,2,張 宏,2
(1.西北農林科技大學農學院,陜西楊凌 712100; 2.農業部作物基因資源與種質創新陜西科學觀測實驗站,陜西楊凌 712100)
小麥(TriticumaestivumL.)作為我國主要糧食作物,發展小麥生產對保障我國糧食生產和安全有重要作用[1]。但遺傳背景日趨狹窄和骨干親本單一[2]的問題制約小麥育種研究,不利于小麥生產。利用遠緣雜交將小麥近緣種作物的優異基因導入小麥,可強化種質資源的原始積累,進而推動創新性品種的培育和生產[1],不斷提高小麥產量和品質[3]。
華山新麥草(Psathyrostachyshuashanica,2n=2x=14,NsNs)主要分布于陜西華山地帶,是珍稀農作物野生親緣種[4-5]。華山新麥草具有抗病、耐寒、耐瘠薄等眾多優異性狀,是小麥抗逆、抗病基因庫[6-7],如華山新麥草中的果聚糖合成酶(6-SFT)可使煙草增強對非生物脅迫的耐受性[8],其含有的高分子量谷蛋白可影響小麥品質[9],將華山新麥草的優良性狀基因引入小麥可改善小麥抗病性和貧瘠土壤的耐受性[7,10-11]。在近年來的研究中,如Du等[12-15]報導的1~7Ns附加系和2Ns(2D)代換系以及王秀娟等[16]研究的代換系在抗條銹病和葉銹病、株高和小穗等農藝性狀上都有優異表現,既為育種工作者提供了更多可選擇的中間育種材料和種質資源,也證明了華山新麥草染色體的導入會給小麥帶來許多優異的農藝性狀。
小麥條銹病是由條銹菌(Pucciniastriiformisf.sp.tritici(Pst))引起的嚴重病害[17],可通過空氣傳播造成小麥20%~50%的產量損失[18]。在小麥抵抗條銹菌侵染的長期過程中,國內許多抗性基因逐漸失去了抗性,如Yr1、Yr2、Yr3、Yr9、Yr10、Yr24和Yr26[19],這與條銹菌小種的頻繁變異有關[19-20]。因此需不斷尋找新抗病基因和抗病品種,有效防控病害和減少作物損失[21]。利用小麥野生近緣種中的豐富基因資源,不斷創制新抗病材料,無疑是小麥育種中重要的基礎性工作 之一。
本課題組前期利用普通小麥7182和華山新麥草的BC1F9代植株獲得三個穩定的小麥-華山新麥草衍生系:H1133、H4122和H1423。本研究利用細胞學鑒定、原位雜交鑒定、分子標記分析、形態學鑒定等方法,分析這3個衍生系中的華山新麥草遺傳物質,評估它們的育種和抗病價值,以期為后續研究提供參考依據。
1 材料與方法
1.1 供試材料
供試材料為普通小麥品系7182(2n=6x=42,AABBDD)、華山新麥草(2n=2x=14,NsNs)以及小麥(7182)-華山新麥草衍生系H1133、H4122和H1423,感病對照品種輝縣紅。以上材料由西北農林科技大學農學院染色體工程實驗室培育和提供。2016-2019年將試驗材料按照家系點播種植于西北農林科技大學試驗地(陜西楊凌),目標單株套袋自交獲得后代種子。條銹菌小種CYR32和CYR33,由西北農林科技大學植物保護學院提供。
1.2 細胞學鑒定
于2018和2019年3-4月份,取田間材料的根尖和幼穗。將根尖在冰水混合物中處理24 h,卡諾固定液Ⅰ(乙醇∶冰醋酸= 3∶1,V/V)處理2 d,1%醋酸洋紅染色,45%醋酸壓片,鏡檢。幼穗用卡諾固定液Ⅱ(乙醇∶氯仿∶冰醋酸= 6∶3∶1,V/V/V)處理3 d,用1%醋酸洋紅染色壓片,鏡檢。
1.3 原位雜交鑒定
染色體制片:將供試材料種子放入墊有2層濕潤濾紙的培養皿中,在23 ℃恒溫黑暗培養箱中進行種子發根處理,待根長至2~4 cm時剪下,N2O處理2 h,90%醋酸冰上固定10 min,置于70%的乙醇中-20 ℃保存。參照Han等[22]方法制備滴片,混合酶液(1%纖維素酶和2%果膠酶)中 37 ℃處理57~59 min(酶解時間因材料而異)。70%乙醇沖洗3次后破碎并離心,除去乙醇,加20 μL冰醋酸,渦旋,取10 μL滴于載玻片上,鏡檢,合格樣品備用。
探針制備:用CTAB法[23]提取華山新麥草基因組DNA,以切口平移法標記華山新麥草基因組DNA探針。參考Tang等[24]的方法,由上海英駿(Invitrogen)生物技術有限公司合成寡核苷酸探針Oligo-pTa535和Oligo-pSc119.2。
原位雜交:參考Lukaszewski等[25]和Yang等[26]的方法,以華山新麥草基因組DNA為探針對材料進行基因組原位雜交(genomicinsituhybridization,GISH)分析;參考Tang等[24]的方法,以寡核苷酸序列Oligo-pTa535(紅色)和Oligo-pSc119.2(綠色)為探針對材料進行熒光原位雜交(fluorescenceinsituhybridization,FISH)分析;以1:1的比例混合FISH探針與GISH探針,按照GISH程序對材料進行FISH-GISH同步雜交分析。用OLYMPUS BX-53熒光顯微鏡進行鏡檢,利用DP800 CCD(OLYMPUS)系統進行圖像采集和合成,用軟件Adobe Photoshop CS6進行圖片處理。
1.4 EST和PLUG標記分析
依據網站http://wheat.pw.usda.gov/SNP/new/pcr_primers.shtml、GrainGenes公布的引物信息和部分發表文獻[12-15,27]中的引物信息,選取位于小麥7個部分同源群上的72對EST-STS引物和88對PLUG(PCR-based landmark unique gene)引物(由北京奧科鼎盛生物技術有限公司合成)對普通小麥7182、華山新麥草、H1133、H4122和H1423進行PCR擴增分析。PCR擴增體系和程序參考Zhang等[28]的方法。
1.5 形態學鑒定和抗病性鑒定
在試驗材料成熟期進行形態學鑒定,統計株高、分蘗、穗長、小穗數和小穗粒數等農藝性狀,記錄標準參照李立會等[29]方法。小麥-華山新麥草衍生系與普通小麥親本之間的差異顯著性用T測驗方法進行統計分析。
在試驗材料幼苗期時,于西北農林科技大學試驗田對誘發行輝縣紅接種條銹菌小種CYR32和CYR33的混合菌種。當感病對照輝縣紅充分發病后,用0~4級分級標準[30]觀察并記錄7182、華山新麥草、H1133、H4122和H1423的條銹病反應型,0級為免疫,0;級為近免疫,1級為高抗,2級為中抗,3級為中感,4級為高感。
2 結果與分析
2.1 細胞學鑒定結果
觀察試驗材料的花粉母細胞,發現在減數第一次分裂中期,H1133(圖1a)、H4122(圖1b)和H1423(圖1c)的染色體構型均為22個二價體;觀察試驗材料的根尖體細胞,發現H1133(圖1d)、H4122(圖1e)和H1423(圖1f)的染色體數均為44條。各材料的細胞統計數均達到30以上,可以初步判定H1133、H4122和H1423具有細胞學穩定性,且攜帶44條染色體并能穩定配對。

圖1 H1133、H4122和H1423的花粉母細胞(a-c)和根尖體細胞(d-f)鏡檢結果
2.2 原位雜交鑒定結果
以華山新麥草基因組DNA為GISH探針(綠),以Oligo-pTa535(紅)和Oligo-pSc119.2(綠)為FISH探針對H1133、H4122和H1423根尖體細胞進行同步原位雜交鑒定,將結果與Tang等[29]的中國春核型圖對比。發現H1133(圖2a)、H4122(圖2b)和H1423(圖2c)均含有普通小麥的42條染色體和2條華山新麥Ns染色體。
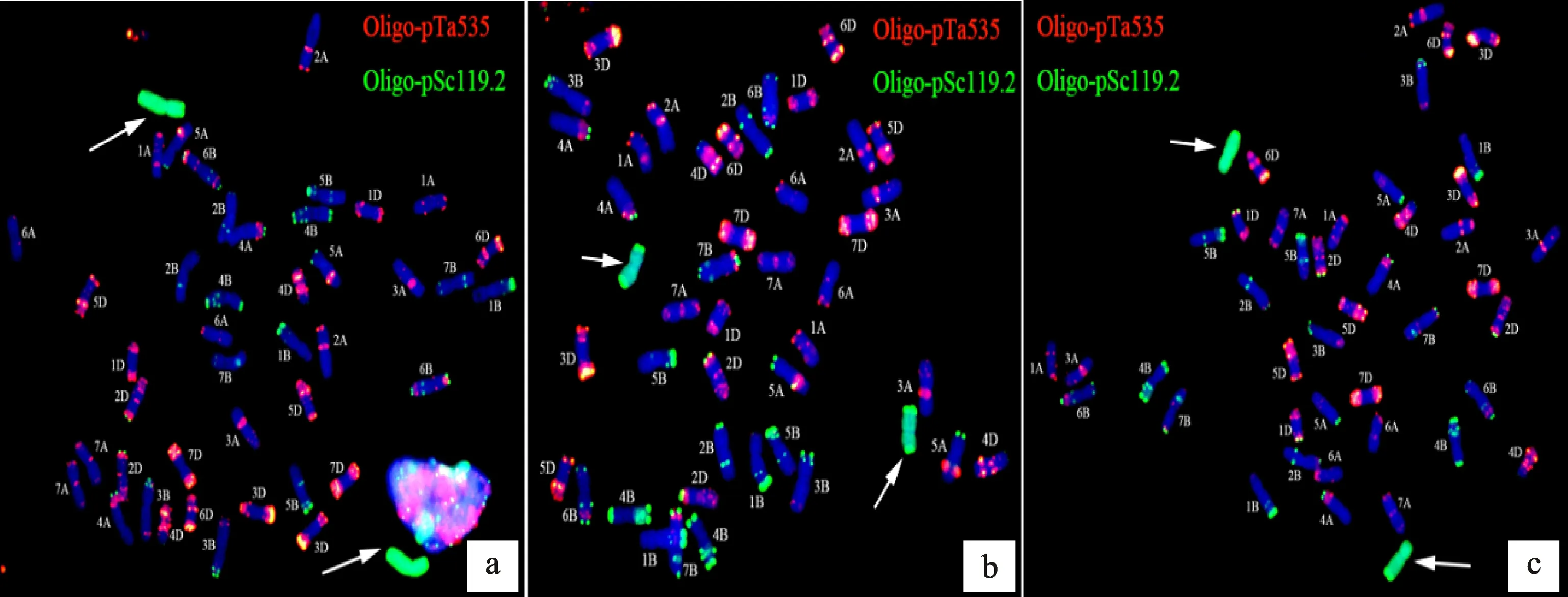
以華山新麥草基因組DNA(綠色)為GISH探針,以Oligo-pTa535(紅色)、Oligo-pSc119.2(綠色)為FISH探針對根尖體細胞進行同步原位雜交,箭頭代表Ns染色體。
2.3 EST和PLUG標記分析結果
為明確各衍生系含華山新麥草Ns染色體所屬同源群,利用72對EST-STS引物和88對PLUG引物分別對H1133、H4122和H1423的Ns染色體進行部分同源群歸屬驗證。將篩選的特異性標記進行分類統計(表1),發現第三同源群中的CD454742(圖3a)、BF473348(圖3b)、CD454575(圖3c)、TNAC1326(圖3d)、TNAC1248(圖3e)可以特異性擴增H1133中華山新麥草特異條帶;第五同源群中BE499166(圖4a)、TNAC1588(圖4b)、TNAC1503(圖4c)可以特異性擴增H4122中華山新麥草特異條帶;第七同源群中BE637663(圖5a)、BG274576(圖5b)、BE591127(圖5c)、TNAC1957(圖5d)、TNAC1821(圖5e)可以擴增出H1423中華山新麥草特異條帶。這說明H1133、H4122和H1423分別攜帶華山新麥草3Ns、5Ns和7Ns染色體。結合細胞學和原位雜交結果,認為H1133、H4122和H1423為3Ns、5Ns和7Ns異附加系。

表1 第三部分、第五部分和第七部分同源群的EST和PLUG標記Table 1 EST and PLUG markers mapped on homeologous group 3,group 5 and group 7
2.4 形態學鑒定和抗條銹病鑒定結果
為了評價各附加系在育種和抗病研究中的價值,對H1133、H4122和H1423的農藝性狀如株高、分蘗、穗長、小穗和小花等進行了觀察和統計(表2)。比較發現H1133、H4122和H1423的平均株高比親本7182低約10 cm。H1133和H4122的平均分蘗比7182多1~2個,而H1423少2個。H4122和H1423的平均穗長約11 cm,較親本7182略有增加,T測驗結果差異顯著 (P<0.05)。植株性狀對照如圖6所示,各附加系的穗型變為紡錘型。
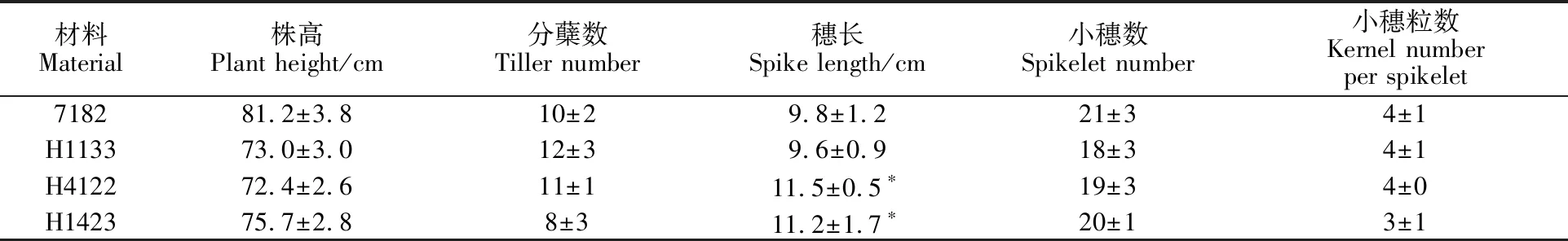
表2 H1133、H4122、H1423和7182的農藝性狀Table 2 Agronomic traits of H1133,H4122,H1423 and 7182
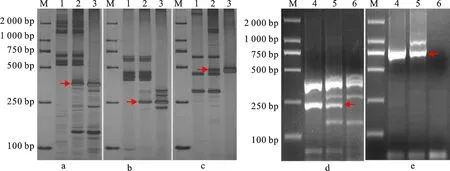
M:DL2000;1:7182;2:H1133;3:華山新麥草;4:華山新麥草;5:H1133;6:7182;a:CD454742標記;b:BF473348標記;c:CD454575標記;d:TNAC1326標記;e:TNAC1248標記;箭頭所指為華山新麥草特異帶。
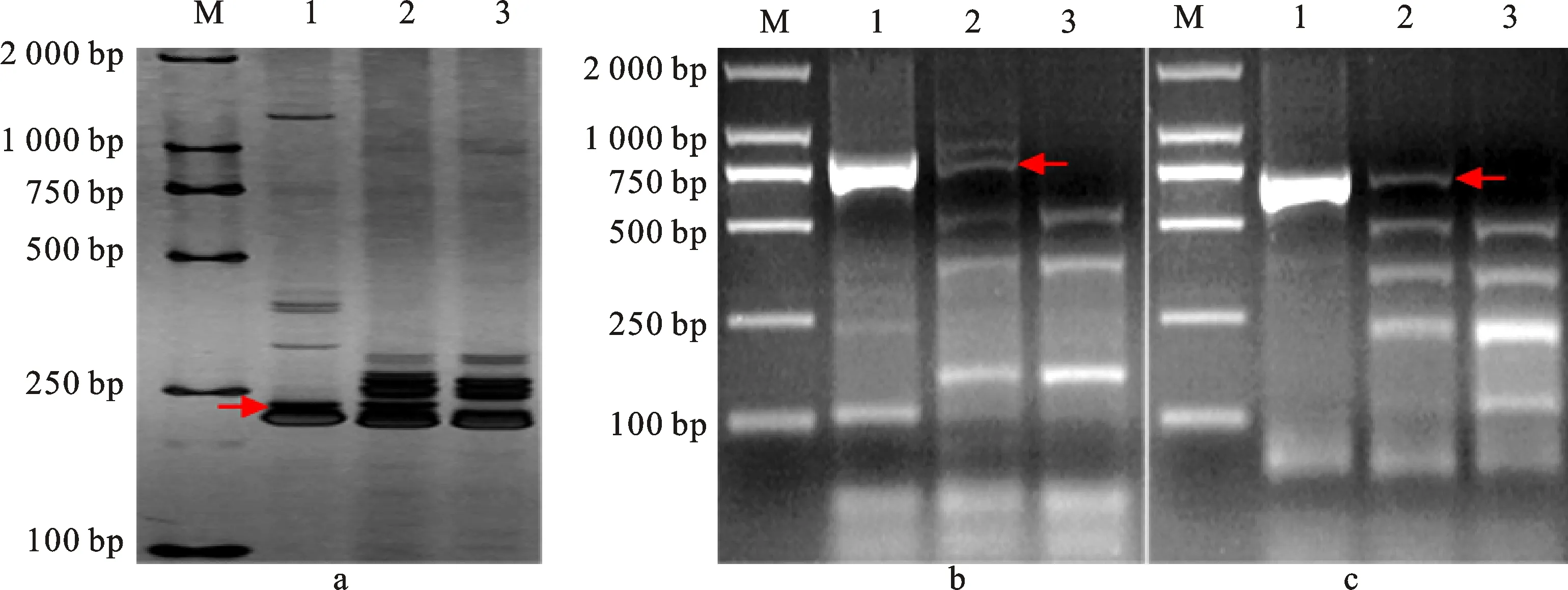
M:DL2000;1:華山新麥草;2:H4122;3:7182;a:BE499166標記;b:TNAC1588標記;c:TNAC1503標記;箭頭所指為華山新麥草特異帶。
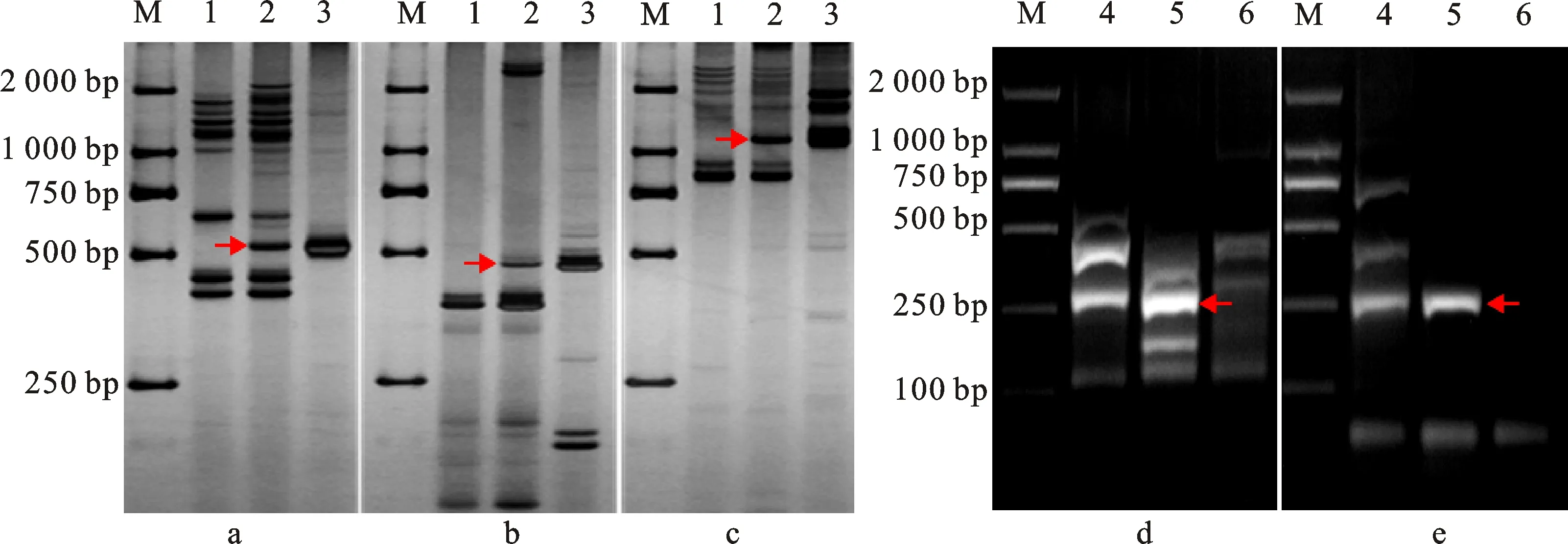
M:DL2000;1:7182;2:H1423;3:華山新麥草;4:華山新麥草;5:H1423;6:7182;a:BE637663標記;b:BG274576標記;c:BE591127標記;d:TNAC1957標記;e:TNAC1821標記;箭頭所指為華山新麥草特異帶。
成株期抗條銹病鑒定結果(圖7)表明:H1133、7182和輝縣紅表現為感病,華山新麥草、H4122和 H1423表現為抗病。根據材料系譜推測H4122和H1423的抗性來源為華山新麥草。

1:7182;2:H1133;3:H4122;4:H1423.

1:華山新麥草(IT0);2:H1133(IT3);3:H4122(IT0;);4:H1423(IT0);5:7182(IT3);6:輝縣紅(IT3)。
2.5 探針的特異性檢驗結果
結合細胞學技術和分子技術,鑒定獲得了3Ns異附加系H1133、5Ns異附加系H4122和7Ns異附加系H1423。為了進一步研究這些導入的Ns染色體的特性,以華山新麥草基因組DNA(綠)、Oligo-pTa535(紅)和Oligo-pSc119.2(綠)為探針,對各材料進行先FISH后GISH的鑒定。對比發現,FISH探針在H1133(圖8a)、H4122(圖8b)和H1423(圖8c)中的Ns染色體上無特異信號,這說明Oligo-pTa535(紅)和Oligo-pSc119.2(綠)無法有效區分3Ns、5Ns和7Ns染色體。
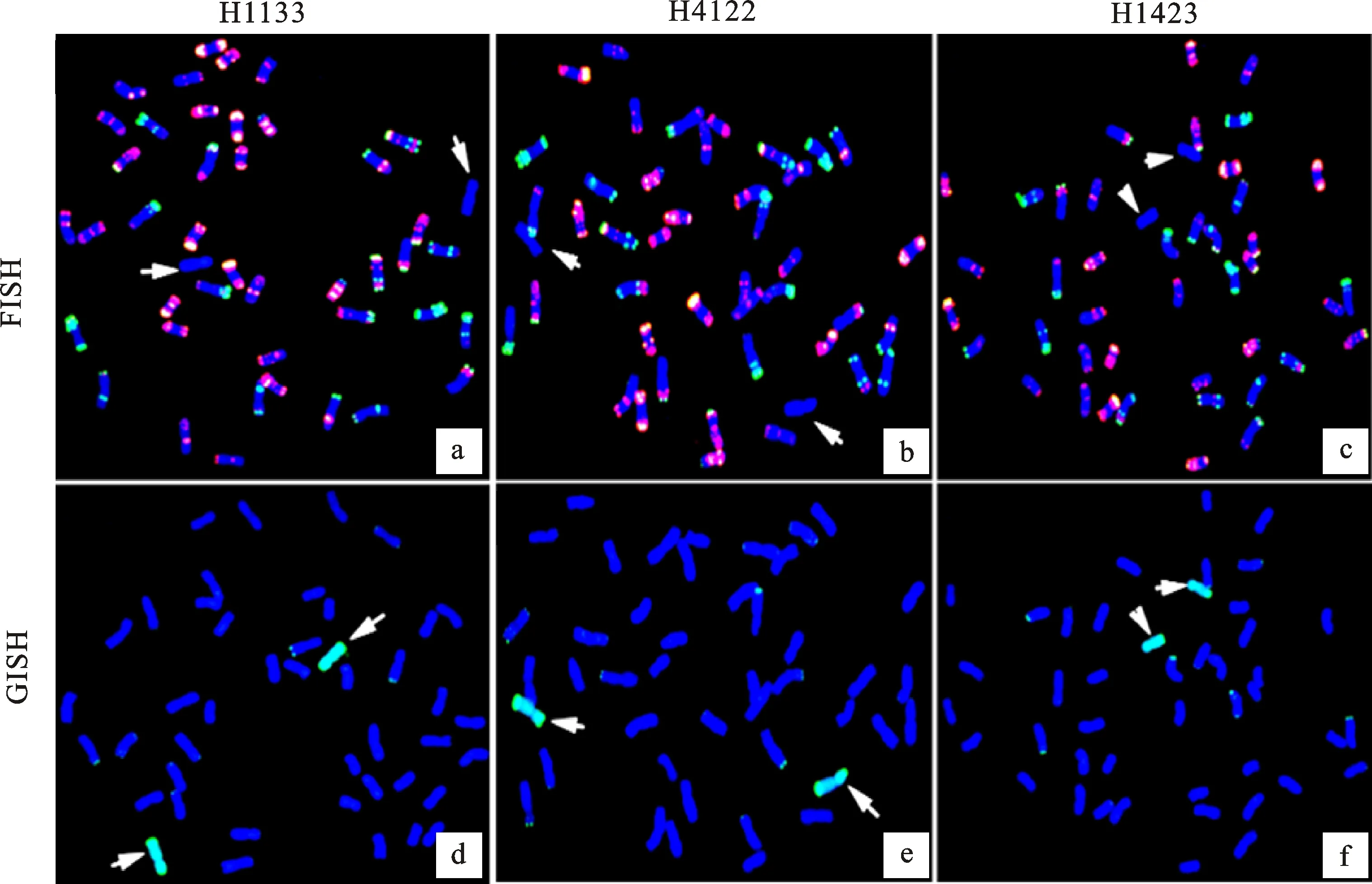
箭頭指示Ns染色體。
3 討 論
導入外源染色體的材料在染色體互作研究中有重要意義[31],可使植株表型產生多種變化,如華山新麥草Ns染色體的導入使H1133、H4122和H1423株高降低、小穗增多,而矮稈、多花等性狀都能直接影響小麥的產量[32-33]。利用遠緣雜交技術將近緣種作物攜帶的優異基因轉入小麥,是拓寬小麥遺傳背景的有效方法。但附加系中整條染色體的導入常引入不利性狀的基因,這意味著H1133、H4122和H1423需進一步改良為易位或滲入系,才能提高其利用價值,如Kang等[34]創制的易位系k-13-835-3具有多籽粒和抗條銹病的優良性狀。而前人的研究給我們指引了一個新方向,如Zhang等[35]利用60Co-γ對附加系進行輻射處理,將攜帶種子貯藏蛋白基因的簇毛麥1V染色體片段導入中國春,提高了面包品質;Zhang等[36]利用同樣方法將冰草的6P染色體片段導入小麥,創制的易位系顯著提高了小麥千粒重和穗長。因此,利用輻射處理不同的華山新麥草Ns附加系材料,可獲得大量的擁有優異農藝性狀的易位系育種材料。總之,華山新麥草是小麥育種研究中的重要基因資源庫,多樣的附加系材料更豐富了小麥的育種資源,為進一步改良和利用華山新麥草中的優異基因奠定了基礎,但后續研究工作仍需我們長期的探索。
在本研究中,探針Oligo-pTa535和Oligo-pSc119.2的組合無法有效區分3Ns、5Ns和7Ns染色體,間接證明了Ns組染色體與中國春染色體組有著不同的寡聚核酸偏好性。王丹蕊等[37]利用寡核苷酸探針套鑒別長穗偃麥草和中間偃麥草染色體的研究,為本研究進一步利用多樣化的探針組合或新的寡核苷酸探針對不同Ns染色體的快速鑒別提供了借鑒和參考。此外,基因組測序和分析技術的完善為開發新的寡聚核酸探針提供了契機,而本研究創制的附加系材料為通過基因組學開發探針提供了基礎材料。在抗條銹病鑒定中,H4122和H1423對條銹病小種CYR32和CYR33混合菌種田間表現為抗病。H1133的田間表現與Du 等[13]的3Ns附加系抗條銹病的鑒定結果存在差異,可能是小種差異或材料遺傳背景差異導致的;Du 等[14]的7Ns附加系研究中未提及有關條銹病抗性的研究,所以本試驗對H1423的研究是對前人成果的補充;馬東方等[40]在華山新麥草易位系H9015-17的5DL上發現了一個新的抗性基因YrHua1,H4122中的抗性基因是否是其等位基因或華山新麥草的新抗病位點,則需要進一步分析研究。總之,普通小麥-華山新麥草附加系H1133、H4122和H1423為育種者提供了更多的選擇機會。
