小麥 TaGS-D1與 TaFlo2-A1等位變異對粒重的影響
2020-07-30 10:02:32嚴勇亮付婧璇耿洪偉
麥類作物學報 2020年5期
張 芳 ,任 毅,嚴勇亮,付婧璇,耿洪偉
(1.新疆農業大學農學院,農業生物技術重點實驗室,新疆烏魯木齊 830052;2.新疆農業科學院,農作物品種資源研究所,新疆烏魯木齊 830091)
千粒重是小麥產量構成三要素之一,提高千粒重是小麥增產的有效途徑和重要育種目標[1]。小麥千粒重是受微效多基因控制的數量性狀,利用可靠的分子標記可提高對小麥粒重選擇的準確性和有效性,有利于對高千粒重優異基因型在小麥育種中的充分應用[2]。產量三要素中千粒重是最穩定,具有59%~80%的廣義遺傳率[3]。胡延吉等[4]認為,粒重的增加是提高小麥產量的主導因素。馮素偉等[5]發現,在穗粒數的貢獻相對穩定時,提高千粒重則成為小麥增產的關鍵因素。由此可見,粒重的遺傳改良是對小麥高產非常重要[6]。國內外的研究表明,過去的幾十年中,小麥穗數和穗粒數的改良相對緩慢,且對產量提升貢獻較小,而千粒重的提高對小麥產量的提升貢獻較大[7-11]。因此,在小麥品種選育過程中,注重千粒重的選擇是提高小麥產量的重要途徑,也成為小麥分子標記輔助育種的一個重要內容[12]。
近年來,小麥粒重相關基因定位取得了很大進展[13-14]。Cui等[15]在5B、6A和7B染色體上發現3個與粒重相關的主效QTL,分別解釋表型變異的11.28%~16.06%、5.64%~18.69%和 6.76%~21.16%。Jia等[16]利用來自RIL群體南大2419/望水白的230個家系,將粒重主效QTL定位于3A、4B、4D和5A染色體上。Huang等[17]在2D、4B和5A染色體上發現了3個粒重主效QTL。與此同時,作為育種應用的理想標記,功能標記被用于小麥粒重基因鑒定和克隆[18-19]。Zhang等[20]克隆了普通小麥7DS染色體上的籽粒大小基因TaGS-D1的全長編碼序列,并基于40 bp 的插入/缺失(Indel),開發了與千粒重顯著相關(P<0.01)的共顯性功能標記GS7D。該位點的兩個等位基因TaGS-D1a和TaGS-D1b分別與高千粒重和低千粒重相關,對應的擴增片段長度分別為562和522 bp。利用GS7D標記對51份新疆冬小麥進行檢測發現,該標記能較好地區分TaGS-D1a和TaGS-D1b等位基因[21]。Flo2是廣泛存在植物中的基因家族,小麥TaFlo2基因與粒重緊密相關,其表達量對小麥籽粒的淀粉含量和蛋白質合成有影響,從而影響小麥的粒重。Sajjad等[22]克隆了普通小麥2A染色體上的粒重基因TaFlo2-A1的全長編碼序列,并根據其等位基因TaFlo2-A1a和TaFlo2-A1b在啟動子區存在8 bp的插入/缺失(Indel),開發了一對共顯性功能標記TaFlo2-Indel8。TaFlo2-Indel8標記可在具有等位基因TaFlo2-A1a(與低千粒重相關)和TaFlo2-A1b(與高千粒重相關)的材料中分別擴增出153和 145 bp的片段。雖然目前有許多小麥粒重相關基因的分子標記被開發和利用[23-26],但TaGS-D1和TaFlo2-A1為粒重基因研究的主要對象。這些研究多是對單個基因效應進行分析,而對不同粒重基因組合對千粒重影響尚不清楚。基于此,本研究利用粒重功能標記GS7D和TaFlo2-Indel8對298份國內外冬小麥品種(系)進行檢測,對各等位基因、基因組合與千粒重的相關性及其在不同小麥類型中分布規律進行分析,以期探究不同粒重基因型對千粒重的作用,為小麥粒重分子聚合育種提供參考依據。
1 材料與方法
1.1 供試材料
供試的298份冬小麥品種(系)來自12個國家,包括65份國外品種(系)及233份國內四個主要冬麥區的歷史品種(系)和當前主栽品種(表2)。其中,國外冬小麥品種(系)分別來自法國(21份)、意大利(9份)、美國(7份)、阿根廷(7份)、羅馬尼亞(6份)、俄羅斯(5份)、日本(4份)、匈牙利(2份)、澳大利亞(1份)、德國(1份)、英國(1份)、土耳其(1份);國內冬小麥品種(系)分別來自黃淮冬麥區(145份)、北部冬麥區(55份)、西南冬麥區(22份)和長江中下游區冬麥區(11份)。所有小麥品種(系)均于2016-2017年和2017-2018年度在新疆農科院瑪納斯試驗站種植。隨機區組設計,2個重復,行長2 m,行距25 cm,3行區,每行播量為100粒,田間管理同大田生產,正常成熟后及時收獲,人工脫粒。
1.2 千粒重測定
采用杭州萬深檢測科技有限公司生產的SC-G型千粒重儀測定小麥千粒重,3次重復,計算平均值。
1.3 分子標記檢測
1.3.1 基因組DNA提取
為避免單粒種子偶然性錯誤,每個品種(系)選取3 粒粒型均勻的種子,粉碎后放入2.0 mL離心管中,參照Lagudah等[27]方法提取基因組DNA,并用1%瓊脂糖凝膠電泳技術檢測DNA。
1.3.2 PCR擴增與檢測
利用Zhang等[20]開發的共顯性標記GS7D檢測小麥7DS染色體上粒重基因TaGS-D1等位基因;利用Sajjad等[22]開發的共顯性標記TaFlo2-Indel8檢測小麥2AL染色體上粒重基因TaFlo2-A1等位基因。引物(表1)均由上海生物工程技術服務有限公司合成。
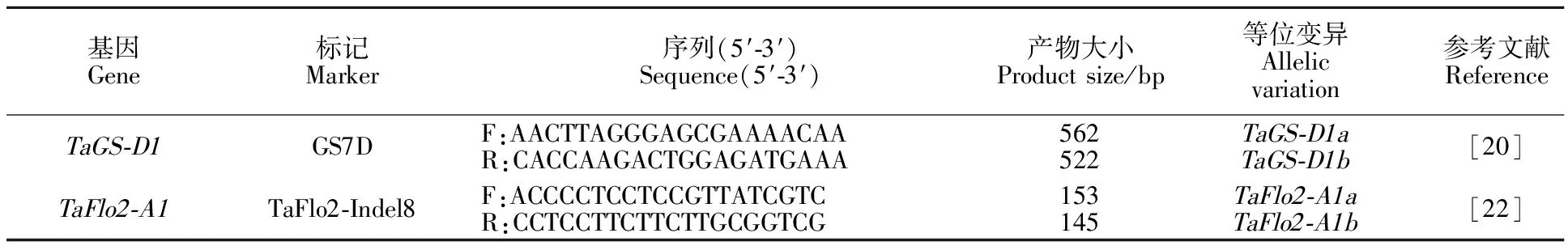
表1 目標基因分子標記信息
PCR體系15 μL,含1 μL的DNA為模板,2×Es Taq MasterMix(Dye)7.5 μL(康為世紀),上、下游引物(10 μmol·L-1)各0.5 μL,用ddH2O補充至15 μL。PCR反應程序為94 ℃ 5 min;94 ℃ 1 min,55~59 ℃ 45 s,72 ℃ 2 min,共35個循環;72 ℃ 10 min。其中,標記GS7D的PCR產物在4.0%瓊脂糖凝膠(加入核酸染料)上進行電泳分離,在VILBER LOURMAT凝膠成像系統紫外燈掃描成像并保存到計算機;標記TaFlo2-Indel8的擴增產物用6.0%的變形聚丙酰胺凝膠電泳分離,銀染后記錄結果。
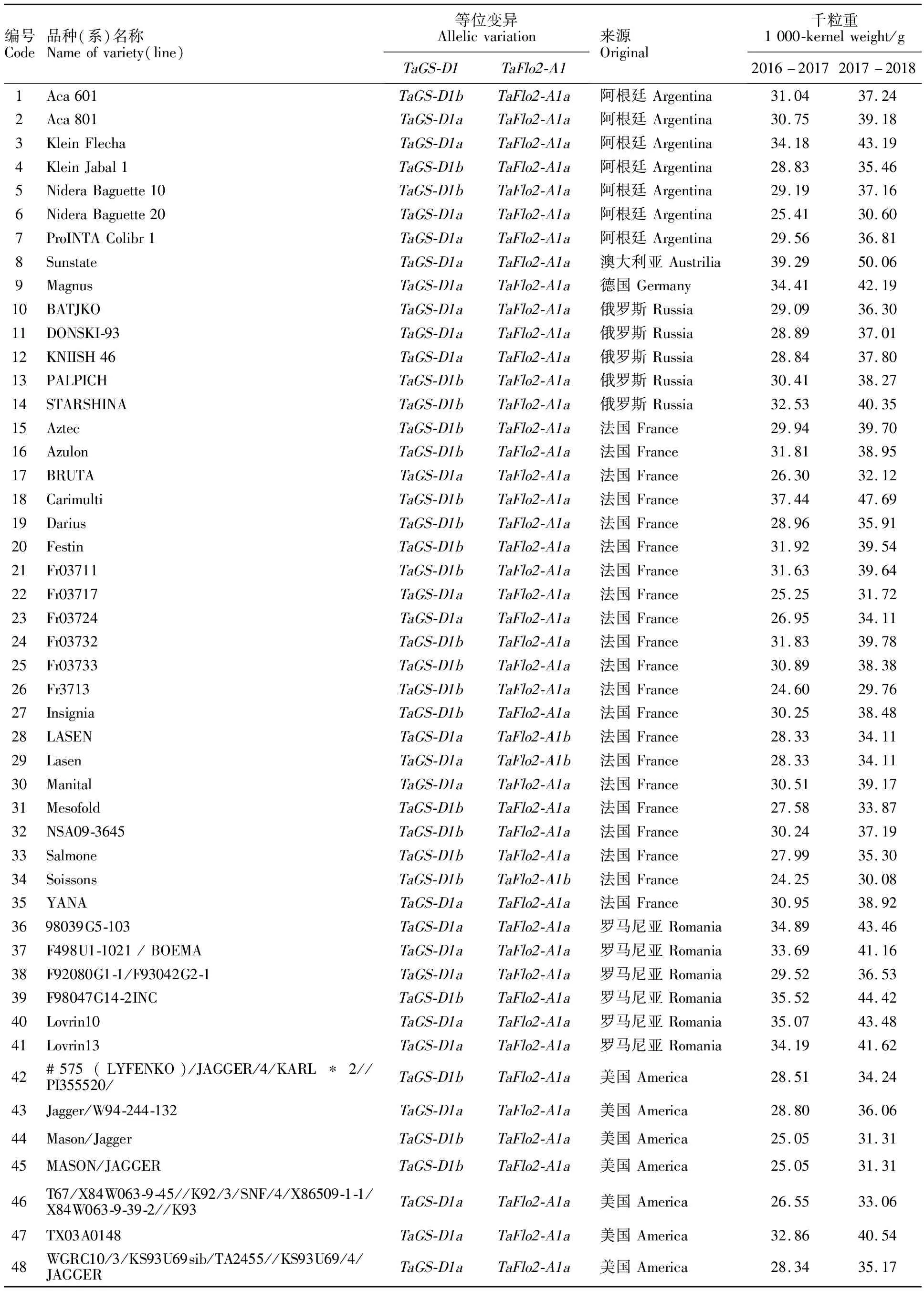
表2 中國和國外小麥品種(系)的千粒重及基因型
1.4 統計分析
采用Excel 2010對測定的千粒重數據進行整理分析,并利用 SPSS 21軟件對同一基因不同等位基因品種的千粒重進行t測驗和不同基因型品種的千粒重間進行鄧肯法多重比較。
2 結果與分析
2.1 小麥千粒重的表型變異
298份小麥品種(系)的兩年千粒重平均值為37.65 g,變化范圍為24.72~46.82 g,變異系數為12.31%;國外材料的平均千粒重低于國內各冬麥區的材料(表3)。
2.2 TaGS-D1和 TaFlo2-A1位點不同等位基因及其組合頻率分布
在298份冬小麥品種(系)中,利用GS7D標記檢測出TaGS-D1a和TaGS-D1b2種等位基因,擴增長度分別是562和522 bp(圖1),其中具有高千粒重TaGS-D1a等位基因的材料有236份(占79.2%),具有低千粒重TaGS-D1b等位基因的材料有62份(占20.8%)。TaGS-D1a和TaGS-D1b在國外及國內各冬麥區中分布不同(表4)。在所有材料中,整體上TaGS-D1a的分布頻率大于TaGS-D1b。從材料來源看,TaGS-D1a的分布頻率表現為西南冬麥區(95.5%)>長江中下游冬麥區(90.9%)>北部冬麥區(85.5%)>黃淮冬麥區(78.6%)>國外(55.4%)。
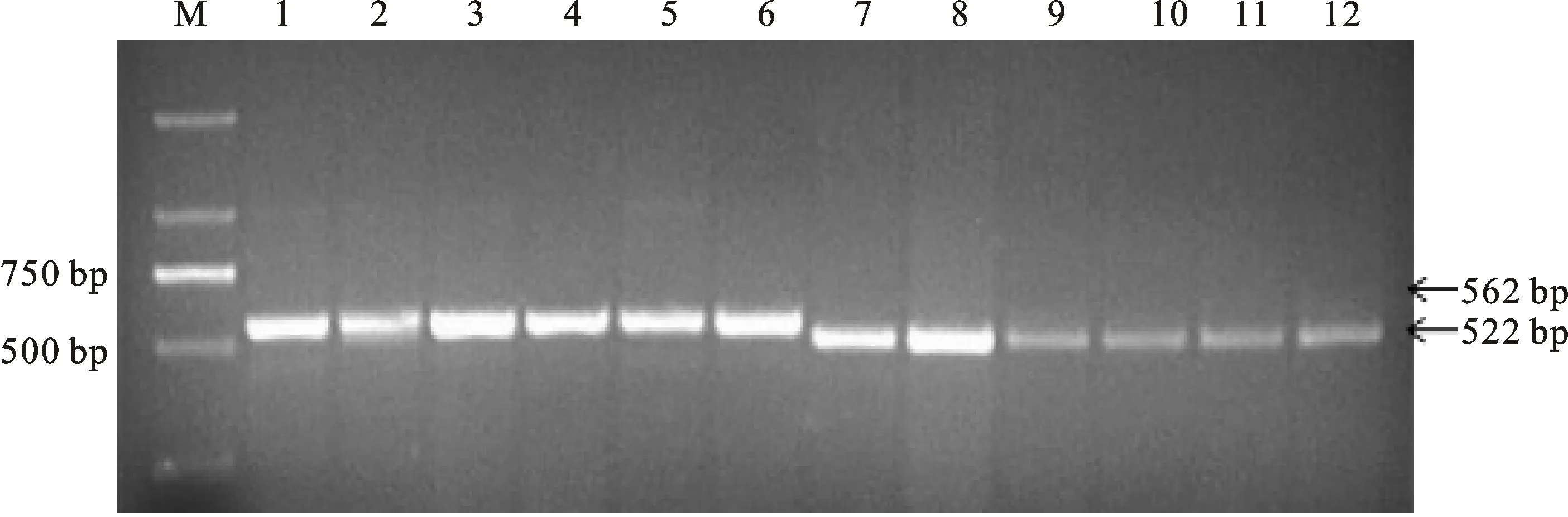
M:DL2000;1:小偃6號;2:豫麥47;3:周麥25;4:魯麥7號;5:濟麥21;6:阿夫;7:碧螞1號;8:百農3217;9:魯麥9號;10:周麥31;11:泰農731;12:豫麥21.
在供試材料中,利用TaFlo2-Indel8標記共擴增出145和153 bp兩種片段類型(圖2),說明存在TaFlo2-A1b和TaFlo2-A1a2種等位基因。在298份冬小麥品種(系)中,56份具有高千粒重TaFlo2-A1b等位基因,242份具有低千粒重TaFlo2-A1a等位基因,說明具有高千粒重TaFlo2-A1b等位基因的材料分布頻率(18.8%)低于具有低千粒重TaFlo2-A1a等位基因的材料(81.2%)。從材料來源看,在參試材料中TaFlo2-A1b的分布頻率表現為長江中下游冬麥區(36.4%)>北部冬麥區(27.3%)>黃淮冬麥區(18.6%)>西南冬麥區(13.6%)>國外(10.8%)(表4)。

表4 不同類型冬小麥品種(系)等位變異的分布頻率
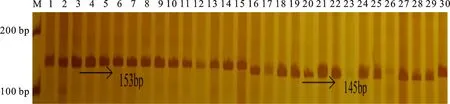
M:DL2000;1:淮麥18;2:淮麥20;3:淮麥21;4:皖麥29;5:皖麥33;6:皖麥38;7:矮抗58;8:武農 148;9:新麥19;10:新麥9號;11:豫麥13;12:豫麥18;13:豫麥34;14:豫麥35;15:豫麥57;16:蘭考906;17:豫麥47;18:豫麥49;19:鄭9023;20:中麥892;21:中麥871;22:周麥12;23:周麥16;24:周麥18;25:周麥19;26:周麥23;27:周麥30;28:揚麥15;29:魯麥23;30:煙農18.
在298份參試材料中共檢測出4種基因組合(基因型),分別為TaGS-D1a/TaFlo2-A1b、TaGS-D1a/TaFlo2-A1a、TaGS-D1b/TaFlo2-A1b和TaGS-D1b/TaFlo2-A1a,其分布頻率分別為15.4%、63.4%、3.0%和18.1%(表5)。在國內各冬麥區和國外品種(系)中,TaGS-D1a/TaFlo2-A1a基因型均占比最大,TaGS-D1b/TaFlo2-A1b基因型均占比最小或沒有。高千粒重基因型在不同小麥類型中均占比較小,且國外品種(系)均低于國內各冬麥區材料。不同來源小麥材料中4種基因型的分布頻率不同。其中,TaGS-D1a/TaFlo2-A1b基因型在長江中下游冬麥區分布頻率較高,達到36.4%;TaGS-D1a/TaFlo2-A1a基因型在西南冬麥區分布頻率較高,占比86.4%;TaGS-D1b/TaFlo2-A1b基因型在國外品種(系)中的分布頻率較高,為4.6%;TaGS-D1b/TaFlo2-A1a基因型在國外品種(系)中的分布頻率較高,達到36.9%。這說明國內小麥資源中高千粒重的TaGS-D1a/TaFlo2-A1b基因型豐富,尤其是在長江中下游冬麥區分布較多,而低千粒重的TaGS-D1b/TaFlo2-A1a基因型在國外品種(系)中分布較多。
2.3 TaGS-D1和 TaFlo2-A1位點不同等位基因及其組合千粒重比較
對TaGS-D1和TaFlo2-A1等位基因及其組合的平均千粒重進行分析(表5)表明,TaGS-D1a和TaFlo2-A1b等位基因的平均千粒重分別為38.18和39.12 g,均極顯著高于對應的等位基因TaGS-D1b和TaFlo2-A1a,進一步印證TaGS-D1a和TaFlo2-A1b等位基因與高千粒重相關,屬于優異等位變異。在TaGS-D1和TaFlo2-A1位點形成的4種基因型中,TaGS-D1a/TaFlo2-A1b的平均千粒重為39.82 g,極顯著高于其他基因型;TaGS-D1b/TaFlo2-A1a的平均千粒重為35.59 g,低于其他3種基因型,說明TaGS-D1a/TaFlo2-A1b與高千粒重相關且為最優組合,對千粒重的增加具有較大作用,且高于單一高千粒重等位基因增效作用。
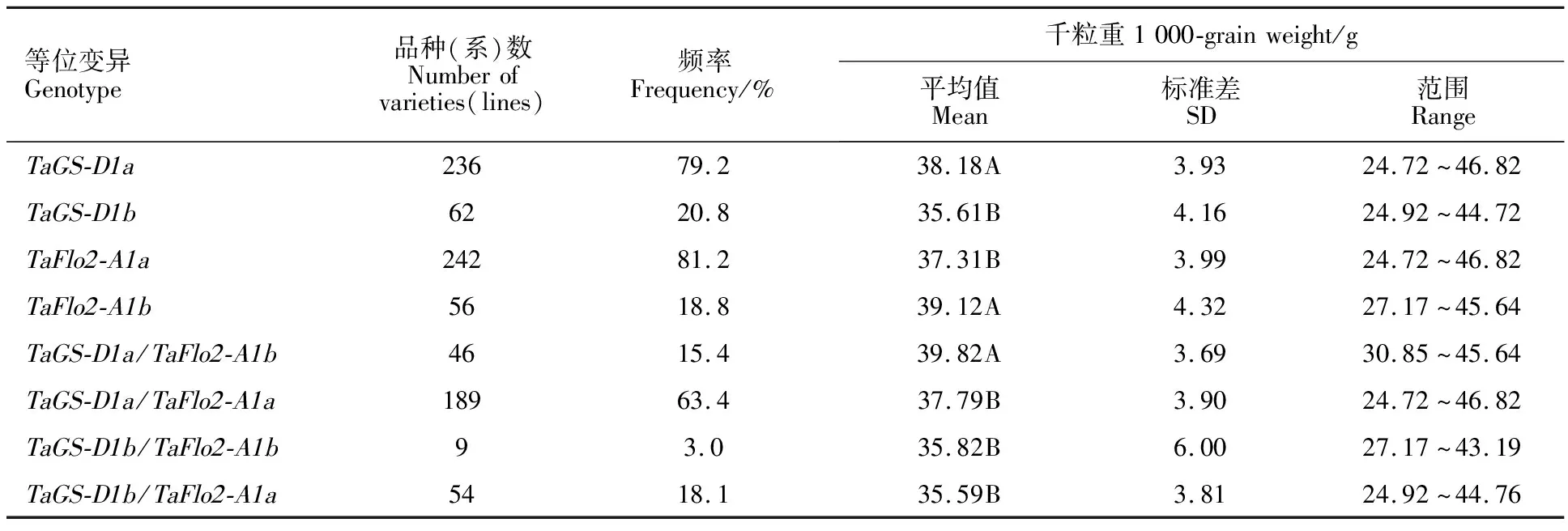
表5 不同基因間和不同基因型間千粒重分析
3 討 論
小麥粒重相關分子標記輔助選擇對小麥育種效率至關重要[28]。盡管粒重基因在品種間和環境間均存在顯著差異,但主要受遺傳控制,通過分子標記技術,可有效提高小麥高產育種效率。利用TaGS-D1位點功能標記GS7D和TaFlo2-A1位點功能標記TaFlo2-InDel8可較好地區分出小麥千粒重的大小。Zhang等[20]利用TaGS-D1位點的顯性標記GS7D檢測 175 份中國小麥材料,結果表明,TaGS-D1a等位基因分布頻率較高(80%),TaGS-D1a等位基因是優異及優勢等位變異。簡大為等也[21]印證了在51份新疆冬小麥資源中TaGS-D1a等位基因(70.6%)分布頻率明顯高于TaGS-D1b等位基因(29.4%)。Sajjad等[22]利用TaFlo2-A1位點的顯性標記TaFlo2-InDel8檢測262份中國小麥微核心種質,結果顯示TaFlo2-A1b占比僅為16.5%。本研究利用兩對功能標記GS7D和TaFlo2-InDel8對298份冬小麥品種(系)進行檢測,結果與Zhang等[20]和Sajjad等[22]研究基本一致。其中高千粒重TaGS-D1a分布頻率較高,可能與育種家在品種選育過程中優先選擇該等位變異有關。而高千粒重TaFlo2-A1b分布頻率較低,可能與千粒重受多基因控制有關,其效應被其他粒重基因所掩蓋,僅在少數材料中體現出高千粒重TaFlo2-A1b等位基因。雖然TaFlo2-A1b等位基因分布頻率遠低于TaFlo2-A1a等位基因,但TaFlo2-A1b等位基因材料較TaFlo2-A1a等位基因材料差異極顯著(P<0.01)。
不同來源小麥品種(系)中,具有高千粒重的TaGS-D1a和TaFlo2-A1b等位基因及其TaGS-D1a/TaFlo2-A1b基因型的分布頻率均存在較大差異(表4),其中長江中下游冬麥區高千粒重分布頻率較高,而國外品種(系)分布較低。究其原因一方面可能與多年粒重人工選擇有關。長江中下游冬麥區以大穗大粒選擇為主,對粒重的重視程度和選擇力度較強,而國外品種產量的變化體現在穗粒數和穗數,千粒重變化不大,對高千粒重選擇較小,該等位基因未被廣泛選擇利用[29]。另一方面可能與育種需求及不同地理環境的生態條件有關。高千粒重小麥將更有利于我國小麥粒重遺傳改良及篩選高產品種。我國長江中下游冬麥區氣候溫和,雨水充沛,但易發生病蟲害,導致穗數不足,千粒重優勢突出[30],而國外人口密度基數較小,對于高千粒重選育需求相對較低,因此高千粒重TaGS-D1a和TaFlo2-A1b等位基因育種選擇利用將集中在個別少數國家中。此外,還可能在品種改良過程中,我們僅關注國外材料中的一些個別性,從而造成國外品種(系)中高千粒重TaGS-D1a和TaFlo2-A1b等位基因占比較少,同時反映出不同地域品種在遺傳組成上的明顯差異[31-33]。
從小麥主栽品種推廣應用以及目前一些新品種的高產紀錄來看,高千粒重對我國小麥產量的提高至關重要[34,2]。高千粒重TaGS-D1a/TaFlo2-A1b基因型的千粒重顯著高于其他組合,且高于單基因高千粒重TaGS-D1a與TaFlo2-A1b等位基因,說明高千粒重基因型將對小麥粒重遺傳改良具有較大的提升空間,同時可作為小麥粒重的輔助選擇的實用性標記。時佳等[35]也證明了較單一高千粒重等位基因,多個基因型可獲得更高千粒重。品種審定中可利用高千粒重基因型將有效準確地獲得高千粒重品種材料;育種家在高千粒重選育過程中也可以通過TaGS-D1a/TaFlo2-A1b基因型,對其選配親本提供研究基礎[36],篩選出具有高產潛力的小麥種植資源,從而提升我國小麥千粒重。千粒重除受多基因控制外,還受籽粒大小、灌漿速率等因素影響。在今后的高千粒重小麥品種選育過程中,對基因型的累加聚合和多指標綜合分析十分重要,將是一種高產育種的研究方向,可加速高產小麥育種步伐。
