lncRNA HOXA11-AS抑制骨關節炎大鼠軟骨細胞凋亡與基質降解的作用研究
2020-07-29 02:06:50錢亮李宏軍施源鄧新超
實用骨科雜志 2020年7期
錢亮,李宏軍,施源,鄧新超
(武漢市第八醫院骨科,湖北 武漢 430010)
骨關節炎(osteoarthritis,OA)是一種退行性關節疾病,其主要特征是關節軟骨退化、軟骨細胞減少、軟骨下硬化重塑和滑膜炎[1],會導致關節疼痛和僵硬,嚴重時引起關節畸形甚至身體殘疾。每年全球有數百萬人受到OA的影響,據估計到2020年OA將成為第四大最致殘的疾病[2]。OA的發病機制涉及多個因素,其中肥胖、機械壓力、性別和衰老是其發生發展的關鍵因素。目前,控制關節炎引起的疼痛和癥狀是兩個主要的治療目標。而軟骨細胞主要負責軟骨的合成-分解代謝平衡,在關節軟骨的調節中起著關鍵作用[3]。越來越多的證據表明,骨關節炎的發生與軟骨細胞死亡、凋亡或程序性細胞死亡有關[4-7]。據報道,軟骨細胞凋亡在OA的軟骨細胞外基質降解中也起著關鍵作用[6,8]。
長鏈非編碼RNA(long-nocoding RNA,lncRNA)是一類長度200 nt的非編碼RNA,有研究表明,許多lncRNA在不同組織中以及在各種發育和病理條件下差異表達,并在腫瘤細胞的惡性生物學行為如增殖、凋亡、遷移和侵襲中具有重要調控作用[9-11]。lncRNA HOXA11-AS是最近發現的lncRNA分子,在胃癌、宮頸癌、卵巢癌等中均發揮著重要的功能[12-15],但其在骨關節炎軟骨組織和細胞中的研究尚未見報道。本研究通過檢測lncRNA HOXA11-AS在骨關節炎軟骨細胞中的表達,來探究其對軟骨細胞凋亡、基質降解以及相關通路的影響,以闡明lncRNA HOXA11-AS在骨關節炎進程中的作用。
1 資料與方法
1.1 主要試劑與材料 主要試劑:弗氏完全佐劑、青霉素鈉/慶大霉素、青霉素/鏈霉素、胰蛋白酶、杜氏改良Eagle培養基(dulbecco's modified eagle medium,DMEM)/特性F12培養基(ham's F12 nutrient medium,F12)購于美國Sigma公司;Lipofectamine2000、Annexin V-FITC試劑盒、細胞計數試劑盒(cell counting kit-8,CCK-8)、酶聯免疫吸附測定法(enzyme-linked immune sorbent assay,ELISA)試劑盒購于碧云天生物技術研究所;RNAiso Plus試劑盒和All-in-OneTMcDNA第一鏈合成試劑盒購于美國GeneCopoeia公司;SYBR Premix Ex TaqⅡ試劑盒購于日本Takara公司;抗Ⅱ型膠原(CollagenⅡ)、聚集蛋白聚糖(aggrecan)、基質金屬蛋白酶13(matrix metallo protein-13,MMP-13)、血小板結合蛋白基序的解聚蛋白樣金屬蛋白酶5(a disintesrin and metalloprotease with thrombospondin type 5 motifs,ADAMTS-5)、抗肌動蛋白(β-non-muscle,β-actin)、抗核因子κB抑制蛋白(anti-nuclear factor kappa B inhibitory protein,IκBα)、抗磷酸化核因子κB抑制蛋白(anti-phosphorylated nuclear factor kappa B inhibitory protein,p-IκBα)、抗核因子κB(nuclear factor kappa B,NF-κB)、抗磷酸甘油醛脫氫酶(glyceraldehyde phosphate dehydrogenase,GAPDH)購于美國Abcam 公司。其他試劑為進口分裝或國產分析純。
材料:清潔級健康雄性Sprague-Dawley(SD)大鼠(100±10)g購于武漢大學動物實驗中心。
1.2 方 法
1.2.1 OA模型建立 將44只清潔級健康雄性SD大鼠隨機分成兩組:正常組與模型組,每組22只。參照Panicker法[16]制作大鼠的OA模型:大鼠進行備皮與75%酒精消毒后,將100μL弗氏完全佐劑注射到右前側膝關節內,分別于第1、3、7天注射1次,每日強制大鼠進行活動60 min。7 d后如果觀察到大鼠右前側膝關節出現紅腫熱痛,同時伴有軟組織腫脹,出現積液以及右前側膝關節僵硬與活動程度受限等現象,說明模型制作成功。正常組大鼠右后肢內膝骨關節周圍組織中與造模大鼠同步注射等容積0.9%氯化鈉注射液。造模成功后處死兩組大鼠,解剖得到關節軟骨組織。
1.2.2 軟骨細胞分離與培養 將切下的軟骨組織剪碎,用含1%青霉素鈉/慶大霉素雙抗液的磷酸鹽緩沖液(phosphate buffered saline,PBS)沖洗,用0.25%胰蛋白酶充分消化,37℃震蕩孵育40 min,離心棄上清液,加入0.2% Ⅱ型膠原酶在37℃下震蕩孵育6 h,200目濾網過濾,收集濾液,離心棄上清液,得到原代軟骨細胞,將其置于含有10%胎牛血清和1%青霉素/鏈霉素抗生素的DMEM/F12培養基中,在37℃、5% CO2的培養箱中培養,每2天更換1次培養基。當細胞融合度達到80%~90%時,使用0.25%胰蛋白酶消化并收集細胞。取第3代的軟骨細胞用于實驗。
1.2.3 細胞轉染 將軟骨細胞按照每孔5×104個的密度接種到6孔板中,培養24 h后進行轉染。分別在兩個無酶離心管中加4 μg HOXA11-AS小干擾RNA(small interfering RNA,siRNA)(或4 μg陰性對照,NC siRNA)+250 μL opti minimal essential medium(OPTI-MEM)、5 μL Lipofectamine 2000+250 μL OPTI-MEM無血清培養基,室溫孵育5 min,將稀釋后的siRNA和Lipofectamine 2000充分混勻,室溫靜置20 min。將培養的細胞用PBS洗滌,添加新鮮培養基,每孔加入上述混合物培養6 h后,棄去上清,PBS清洗后加新鮮培養基繼續培養。
1.2.4 CCK-8檢測 將轉染的軟骨細胞以1×104個/孔均勻鋪于96孔板中,在37℃、5% CO2培養箱中孵育。分別于0、6、12、24、48、96 h時,每孔加10 μL CCK-8溶液,再將培養板放在培養箱中孵育2 h,在酶標儀上450 nm處檢測各組吸光光度值。
1.2.5 流式細胞術檢測 軟骨細胞轉染48 h后,收集懸浮細胞,離心棄去培養基,用不含乙二胺四乙酸(ethylene diamine tetraacetic acid,EDTA)的胰蛋白酶消化,預冷的PBS洗滌兩次,加入500 μL 1×Binding Buffer重懸細胞,在細胞懸浮液加入5 μL Annexin V-FITC,接著加入10 μL碘化丙啶(propidium iodide,PI),室溫避光孵育20 min,立即用流式細胞儀進行檢測,數據用CELL QUEST軟件分析處理。
1.2.6 ELISA檢測 軟骨細胞轉染48 h后收集并離心,得到細胞上清液。加入上清液于96孔板,在37℃下孵育2 h,PBS去除板內液體,洗滌后用濾紙印干;每孔加入抗Bcl-2相關X蛋白(Bcl 2-associated X protein,Bax)、抗B淋巴細胞瘤-2(B-cell lymphoma-2,Bcl-2)、抗半胱氨酸蛋白酶3(caspase-3)一抗工作液(稀釋為1︰1 000),37℃孵育1 h,去除板內液體,再次洗滌并用濾紙印干;每孔加入辣根過氧化物酶(horse radish peroxidase,HRP)標記山羊抗兔免疫球蛋白抗體(immunoglobulin G,IgG抗體)(1︰5 000),置37℃孵育1 h,洗板后印干;每孔加入底物工作液,置37℃暗處反應10 min,加入終止液,酶標儀檢測490 nm處光密度值(optical density,OD)。
1.2.7 實時熒光定量聚合酶鏈式反應 用RNAiso Plus提取轉染48 h后的軟骨細胞總核糖核酸(ribonucleic acid,RNA),根據Gene Copoeia逆轉錄試劑盒合成互補脫氧核糖核酸(complementary DNA,cDNA),以cDNA為模版,采用SYBR Premix Ex TaqⅡ試劑盒檢測基因表達,以GAPDH作為為內參照進行擴增。擴增體系:上游引物1μL、下游引物1 μL、cDNA 2 μL、SYBR Premix Ex TaqⅡ 12.5 μL、無酶水8.5 μL。反應條件:95℃預變性3 min,95℃變性15 s,60℃退火15 s,72℃延伸40 s,共40個循環。實驗重復3次,采用2-△△Ct法計算各檢測目的基因的相對表達量,使用的引物序列見表1。

表1 實時熒光定量PCR引物序列
1.2.8 蛋白質印跡法 使用RIPA裂解緩沖液(radio immuno precipitation assay lysis buffer,RIPA lysis buffer)提取轉染48 h后軟骨細胞總蛋白,用聚氰基丙烯酸正丁酯(bicinchoninic acid,BCA)測定蛋白質濃度。將蛋白樣品上樣到10%十二烷基硫酸鈉聚丙烯酰胺凝膠電泳(sodium dodecyl sulfate polyacrylamide gel electrophoresis,SDS-PAGE)進行電泳,切膠并轉移至聚偏二氟乙烯膜(polyvinylidene fluoride,PVDF),5%脫脂奶粉室溫封閉2 h。加入抗Collagen-Ⅱ、抗aggrecan、 抗MMP-13、抗ADAMTS-5、抗β-actin、抗IκBα、抗p-IκBα、抗NF-κB p65 4℃過夜。洗膜,加HRP標記的山羊抗兔 IgG抗體(1∶5000),室溫孵育2 h;洗膜后,采用ECL化學發光液(electrochemiluminescence,ECL)顯影,使用Image-Pro Plus 6.0分析蛋白質灰度值。
1.2.9 細胞免疫熒光染色 將轉染48 h后軟骨細胞用PBS清洗后,加4%多聚甲醛中固定15 min,在含有0.5%TritonX-100的PBS中通透15 min,PBS清洗。在室溫下用10%山羊血清封閉細胞2 h,隨后加入NF-κB一抗(1︰200),4 ℃孵育過夜。第2天用PBS洗滌后,滴加FITC標記的熒光二抗(1︰250)室溫避光孵育1 h,然后加4',6-二脒基-2-苯基吲哚(4',6-diamidino-2-phenylindole,DAPI)避光孵育5 min,PBS清洗3次后使用熒光顯微鏡捕獲圖像。
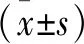
2 結 果
2.1 HOXA11-AS在OA軟骨細胞中的表達情況 使用qRT-PCR檢測HOXA11-AS在OA大鼠的表達是否發生變化。通過檢測OA大鼠和正常組大鼠的軟骨細胞發現,與正常組大鼠軟骨細胞相比,OA大鼠軟骨細胞中HOXA11-AS的表達水平顯著上調(P<0.01,見圖1)。

注:*與正常組相比,P<0.01
2.2 HOXA11-AS對軟骨細胞凋亡的影響 為進一步揭示HOXA11-AS對OA軟骨細胞的影響,使用siRNA技術來沉默HOXA11-AS在軟骨細胞中的表達。經過RT-PCR檢測發現,與陰性對照組(NC siRNA)對比,轉染HOXA11-AS siRNA 48 h后,軟骨細胞中HOXA11-AS的表達呈顯著下降(見圖2a,P<0.01)。通過CCK-8檢測不同轉染時間(0、6、12、24、48、96 h)軟骨細胞的活力,與NC siRNA組相比,軟骨細胞轉染HOXA11-AS siRNA后,在轉染12、24、48、96 h后細胞的活性顯著下降(見圖2b,P<0.01),結果表明HOXA11-AS能提高軟骨細胞活性。

a qRT-PCR檢測轉染48 h后細胞中HOXA11-AS的表達水平 b CCK-8檢測不同轉染時間軟骨細胞的活力
通過流式細胞術檢測轉染siRNA后軟骨細胞的凋亡情況,結果顯示:與NC siRNA相比,轉染HOXA11-AS siRNA的軟骨細胞,其凋亡率明顯提高(P<0.01),說明HOXA11-AS能夠抑制軟骨細胞的凋亡(見圖3)。

注:*與正常組相比,P<0.01
通過ELISA檢測轉染后細胞上清液中大鼠軟骨細胞凋亡標志物發現,與NC siRNA相比,轉染HOXA11-AS siR-NA的軟骨細胞Bax、Caspase-3的表達量顯著上升(P<0.05),而Bcl-2的表達量顯著低于NC siRNA組(P<0.05,見表2)。

表2 兩組大鼠軟骨細胞凋亡標志物表達量比較
2.3 HOXA11-AS對軟骨細胞基質降解的影響 接著我們研究了HOXA11-AS對軟骨細胞基質降解的影響,與NC siRNA相比,轉染HOXA11-AS siRNA的軟骨細胞中MMP-13、ADAMTS-5 mRNA及蛋白表達水平顯著上調(P<0.01),而Collagen Ⅱ、aggrecan的mRNA和蛋白表達水平均顯著下調(P<0.01,見圖4),這一結果表明HOXA11-AS能抑制軟骨細胞外基質的降解。
2.4 HOXA11-AS對NF-κB信號通路的影響 與NC siRNA組比較,轉染HOXA11-AS siRNA組的軟骨細胞中IκBα蛋白表達水平明顯降低(P<0.01),p-IκBα和NF-κB p65的蛋白表達水平均明顯升高(P<0.01,見圖5a)。與此結果一致,通過細胞免疫熒光檢測發現HOXA11-AS siRNA組中NF-κB p65在軟骨細胞核中的表達水平較高(見圖5b)。由此說明沉默HOXA11-AS的表達后激活了NF-κB信號通路,可見HOXA11-AS可抑制NF-κB信號通路的激活。

a Western blot檢測轉染后軟骨細胞中基質降解相關基因蛋白表達情況

b Western blot檢測轉染后軟骨細胞中基質降解相關基因蛋白表達情況

a qRT-PCR檢測轉染后軟骨細胞中基質降解相關基因mRNA表達情況

b 免疫熒光檢測轉染后軟骨細胞核中NF-κB p65的表達情況(免疫熒光檢測,400 μm)
3 討 論
軟骨細胞是關節軟骨中存在的唯一一種細胞類型,它能夠調節軟骨基質合成與分解之間的平衡。當關節受到過度的機械壓力或炎癥刺激時,軟骨細胞會就發生變性和凋亡,從而導致軟骨基質的合成與分解失衡,誘發骨關節炎的不可逆病理過程[17]。越來越多的證據表明,軟骨細胞的凋亡極大地促進了骨關節炎的發生與發展[18-20]。
通過近年來的研究發現,lncRNA參與了多種生物學過程,如控制細胞周期、調節免疫應答調節、調控細胞分化和維持胚胎干細胞多能性等[21]。同時,越來越多的證據表明,多種lncRNA與OA的發病機制有關。例如,Steck等[22]證明lncRNA H19是軟骨細胞中合成代謝的標志物;Su等[10]發現,與正常對照相比,OA軟骨樣品中的lncRNA MEG3表達明顯下調,并可能通過調節血管生成來促進OA的發展進程;Song等[11]的研究表明,與正常軟骨細胞相比,在OA患者的軟骨細胞中發現了lncRNA GAS5高表達,而GAS5的過表達還誘導了一些MMPs家族成員的表達,如基質金屬蛋白酶2(matrix metallo protein-2,MMP-2)、基質金屬蛋白酶3(matrix metallo protein-3,MMP-3)、基質金屬蛋白酶9(matrix metallo protein-9,MMP-9)和MMP-13。由此說明,lncRNA在骨關節炎軟骨細胞中起到一定的作用,有希望作為OA治療的靶標。然而,關于HOXA11-AS在骨關節炎中的研究至今卻未見報道。通過本研究,我們建立了骨關節炎的大鼠模型,并檢測了HOXA11-AS在軟骨細胞中的表達情況,發現與正常組相比,HOXA11-AS在OA大鼠的軟骨細胞中高表達。
為了進一步確認HOXA11-AS在OA中的功能作用,通過siRNA技術抑制了HOXA11-AS的表達,在不同時間段測定軟骨細胞的存活率后發現,從轉染HOXA11-AS siRNA 12 h后開始,細胞活力顯著下降。同樣,在轉染HOXA11-AS siRNA的軟骨細胞中,細胞凋亡率高于轉染NC siRNA對照組,相關細胞凋亡標志物的表達也發生了變化,促凋亡因子Bax、Caspase-3的表達量顯著上升,而抑凋亡因子Bcl-2的表達量顯著下降,這些結果均從側面反映了HOXA11-AS能夠抑制軟骨細胞的凋亡。
而在骨關節炎的發展中,細胞外基質降解起著至關重要的作用,降低軟骨細胞基質降解水平可以抑制骨關節炎的進展[23]。aggrecan和Collagen Ⅱ是細胞基質的重要成分,MMP和ADAMTS的過表達能夠促進細胞外基質的降解,從而加速軟骨組織的破壞[24],當MMP-13和ADAMTS-5過度表達時,促進Collagen Ⅱ和aggrecan的分解。通過研究發現,轉染HOXA11-AS siRNA組的軟骨細胞中,MMP-13、ADAMTS-5基因和蛋白表達水平均明顯升高,同時,Collagen Ⅱ和aggrecan的基因和蛋白表達水平明顯下降[25]。由此說明,HOXA11-AS可以抑制軟骨細胞中MMP-13和ADAMTS-5的過表達以及Collagen Ⅱ和aggrecan表達下降,從而抑制細胞外基質降解。
NF-κB通路作為調控炎癥反應的經典通路,被激活后會引起一系列炎癥級聯反應,從而誘導細胞凋亡和軟骨細胞外基質降解,促進軟骨組織退化[26]。通過本研究檢測發現,轉染HOXA11-AS siRNA組軟骨細胞中IκBα蛋白表達水平明顯降低,同時p-IκBα和NF-κB p65均明顯升高,進而通過細胞免疫熒光檢測得到NF-κB p65在HOXA11-AS siRNA組軟骨細胞核中的表達水平較高。IκBα是NF-κB活化的抑制劑,但其活化后能顯著促進NF-κB活化,NF-κB轉移入核并表達為NF-κB p65[27]。結合實驗結果表明,HOXA11-AS可抑制NF-κB信號通路的激活。
通過本研究我們首次明確了lncRNA HOXA11-AS在OA軟骨細胞中高表達,HOXA11-AS的表達會抑制軟骨細胞凋亡、基質降解和抑制NF-κB通路的激活,由此說明HOXA11-AS在骨關節炎進程中起著重要的作用,可作為骨關節炎預防與治療的靶點。
猜你喜歡
中老年保健(2021年5期)2021-12-02 15:48:21
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
基層中醫藥(2021年9期)2021-06-05 07:14:14
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
今日農業(2020年19期)2020-12-14 14:16:52
臨床醫藥文獻雜志(電子版)(2017年11期)2017-05-17 04:48:39
海峽科技與產業(2016年3期)2016-05-17 04:32:12
