土壤鎘污染對香樟幼苗光合和生理特性的影響*
2020-07-28 09:13:32周際海郜茹茹段洪浪濮海燕金志農
林業科學 2020年6期
周際海 程 坤 郜茹茹 段洪浪 濮海燕 金志農
(1.南昌工程學院 江西省退化生態系統修復與流域生態水文重點實驗室 南昌 330099; 2.安徽師范大學生命科學學院 重要生物資源保護與利用研究安徽省省級重點實驗室 蕪湖 241000)
隨著工農業集約化發展,大量重金屬污染物通過污水灌溉、農藥化肥施用、礦產資源開采等方式進入土壤,影響著人類和動植物的生存環境,其中鎘(Cd)污染問題尤為突出(曾星等, 2019)。鎘易被植物根系吸收,因此較高濃度鎘污染對植物體的生長會產生不良影響,主要表現為植物體光合作用降低、葉綠素含量減少、生物量下降、氣孔關閉、葉片萎縮、甚至出現死亡等現象(趙秀峰等, 2017; 呂波等, 2018; Zhangetal., 2003; Sandersonetal., 2019)。土壤中鎘等重金屬不僅限制植物正常生長,還可通過食物鏈進入人體,危害人體健康,20世紀60年代發生的日本“痛痛病”事件就是典型的鎘中毒事件。
土壤重金屬污染修復的方法有多種,其中植物修復因投資成本低、無二次污染等諸多優點受到重視。植物修復是利用植物固定、吸收或提取土壤中的重金屬,以達到降低其生物有效性或去除污染物的效果(陳順鈺等, 2019)。現已發現許多草本植物,如天藍遏蘭菜(Thlaspicaerulescens)(Bakeretal., 1994)、東南景天(Sedumalfredii)(朱凰榕等, 2019)、三葉鬼針草(Bidenspilosa)(劉沙沙等, 2018)、龍葵(Solanumnigrum)(Yuetal., 2015)和蜈蚣草(Pterisvittata)(楊樹深等, 2017)等對Cd具有超富集作用。
香樟(Cinnamomumcamphora)是我國南方主要園林與經濟樹種,主要用于園林綠化與精油提取等,有較高經濟價值。目前,有研究表明香樟對重金屬污染物具有一定富集能力(Hoetal., 2014; Zhanetal., 2011),而關于重金屬污染土壤中香樟生理生化特征變化的研究卻鮮有報道。將香樟用于重金屬污染土壤的修復,既可以治理重金屬污染,又可提取精油,獲得環境和經濟雙重效益。本研究選取1年生香樟作為供試植物,研究鎘對香樟的凈光合速率、葉綠素等生理生化特性的影響,為科學評估香樟用于鎘污染土壤的生態恢復提供初步的科學依據。
1 材料與方法
1.1 試驗材料 0~20 cm土層鮮土采自進賢縣江西省紅壤研究所。剔除石塊及根茬等,自然風干,過2 mm篩后供盆栽用。土壤基本理化性質為: 有機質7.24 g·kg-1,全氮1.58 g·kg-1,全磷0.96 g·kg-1,有效磷128.57 mg·kg-1,總鎘0.26 mg·kg-1,pH4.35,飽和含水量42.3%。1年生香樟由南昌工程學院香樟種質資源圃提供,株高28 cm,基徑4.12 mm,每株保留3個側枝。
1.2 試驗設計 采用室內盆栽試驗,依據中國《土壤環境質量標準》(GB15618—1995),將鎘污染濃度分別設置為0(CK)、1 mg·kg-1(Cd1,一級標準值的5倍)、10 mg·kg-1(Cd2,一級標準值的50倍)、30 mg·kg-1(Cd3,一級標準值的150倍),每個處理設5個重復。精確配置2 000 mg·L-1的含Cd2+溶液作為母液,依試驗設計,從盆栽底部均勻澆灌于盆中,使土壤鎘濃度分別達到相應脅迫濃度,栽種1年生香樟幼苗。按農田常規施肥量N 300 kg·hm-2、P 150 kg·hm-2,將水溶性肥料(N ≥ 30 g·L-1、P2O5≥ 14 g·L-1)稀釋后作基肥每盆施用40 mL。盆栽置于光照試驗臺上,盆的大小為直徑15 cm、高24 cm,每盆裝土2.5 kg。每周更換盆栽位置,每天稱量并澆入去離子水,使盆栽土壤含水量保持在飽和含水量的50%。培養期間光照與黑暗各12 h·d-1(開、關燈時間為7:00、19:00,光照強度200 μmol·m-2s-1),控制室內溫度為25~30 ℃,分別于第0、15、30、60天時,采樣測定香樟葉片光合作用參數、葉綠素等生理生化指標。
1.3 測定方法 光合作用參數: 采用帶有紅藍光源的Li-6400XT型便攜式光合測量系統,于采樣當天上午9: 00—11: 00,測定距離頂端的第5片葉在飽和光強下的凈光合速率(Pn)、氣孔導度(Gs)、胞間CO2濃度(Ci)和蒸騰速率(Tr)等; 測定的飽和光強為1 200 μmol·m-2s-1,流速為500 mL·min-1,CO2濃度為400 μmol·mol-1,葉室溫度為28 ℃。葉綠素(Chl)含量測定采用95%乙醇浸提法; 脯氨酸(Pro)含量測定采用酸性茚三酮比色法; 可溶性蛋白含量采用測定考馬斯亮藍G-250比色法; 可溶性糖含量測定采用蒽酮比色法; 超氧化物歧化酶(SOD)活性測定采用氮藍四唑比色法; 過氧化物酶(POD)活性測定采用愈創木酚比色法; 丙二醛(MDA)含量測定采用硫代巴比妥酸法(李合生, 1999)。
1.4 數據統計分析 試驗數據采用Excel 2013進行統計,利用SPSS 19.0進行單因素(One-way ANOVA)方差分析,Origin 9.5進行非線性回歸分析并繪圖。
2 結果與分析
2.1 鎘對香樟葉片光合作用參數的動態影響 由圖1可知,香樟葉片凈光合速率(Pn)、氣孔導度(Gs)、胞間CO2濃度(Ci)、蒸騰速率(Tr)和瞬時水分利用效率(WUEi)隨著鎘脅迫時間的增加,各鎘濃度處理的氣體交換參數均呈下降趨勢,且脅迫后期(30和60天)高濃度鎘處理(Cd3)對凈光合速率影響較顯著。第15天時, Cd1的Pn、Gs、Ci和Tr比對照CK分別增加約4.25%、35.78%、34.71%和24.15%,Cd2的分別增加約4.59%、40.37%、50.04%和11.78%,而Cd3的各項參數(凈光合速率除外)均低于CK。第30天時,不同鎘濃度的Pn顯著降低,且隨鎘濃度增加逐漸下降,與CK相比,Cd1、Cd2和Cd3的Pn分別下降11.74%、10.48%和20.35%(P<0.05); 不同鎘濃度的Gs較CK平均顯著降低約21.23%,而不同鎘濃度處理間差異不顯著; 不同濃度鎘處理的Ci均顯著低于CK(P<0.05),其中Cd1的Ci最小,降低約12.56%; 不同鎘濃度處理均可降低香樟葉片的瞬時水分利用效率WUEi,而增加其氣孔限制值Ls。第60天時,不同鎘濃度的Pn較CK均有不同程度降低,其中Cd3的Pn顯著低于CK約24.32%(P<0.05); Cd1、Cd2和Cd3的Ci分別較CK顯著增加約33.49%、41.01%和36.40%(P<0.05); Cd1和Cd2的Tr分別較CK顯著增加約33.48%和54.60%(P<0.05); 而各濃度鎘處理的WUEi和Ls均低于CK,其中Cd2的最低,分別降低約41.13%和28.47%。整個培養期內各濃度鎘處理的Pn、Gs均有下降趨勢,但對香樟生長無明顯影響。以上結果表明,鎘脅迫對香樟光合作用各參數都有一定程度的影響,但香樟對本研究中的鎘脅迫均有一定耐性。

圖1 不同鎘濃度對香樟葉片光合作用參數的影響Fig.1 Effects of different concentrations of cadmium on the photosynthetic characteristics parameters
由圖2可知,各處理的凈光合速率和氣孔導度間呈非線性關系(y=k·ln(x) +yasym,asym代表氣孔導度趨向1時的增大凈光合速率),凈光合速率隨氣孔導度增加而增加,但存在閾值。由表1可知,曲線回歸系數R2、k和yasym的相關關系,不同處理間的凈光合速率和氣孔導度無顯著相關關系,說明本研究香樟的光合參數變化主要與鎘脅迫有關。
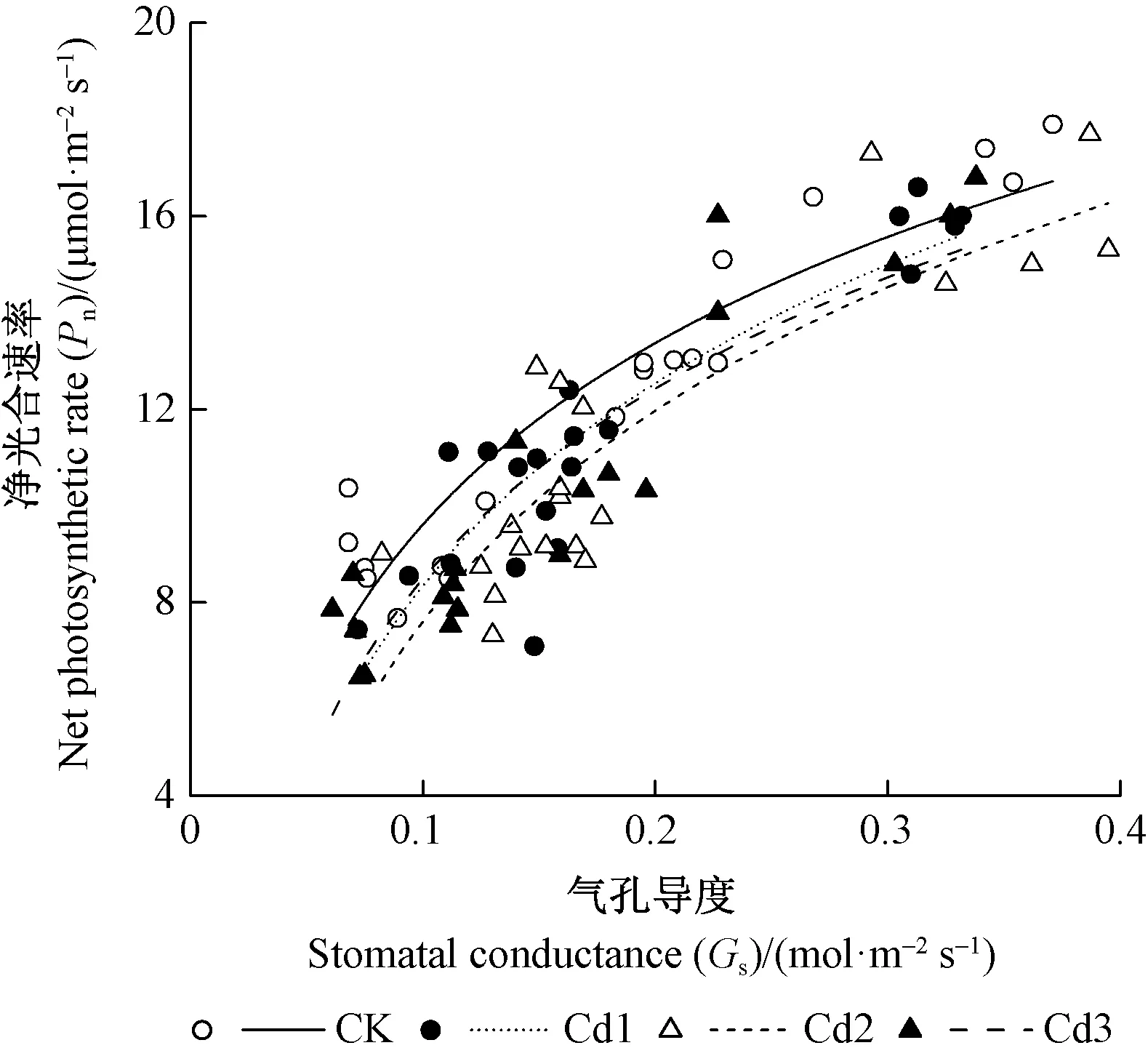
圖2 凈光合速率與氣孔導度關系Fig.2 The relationship between net photosynthetic rate and stomatal conductance
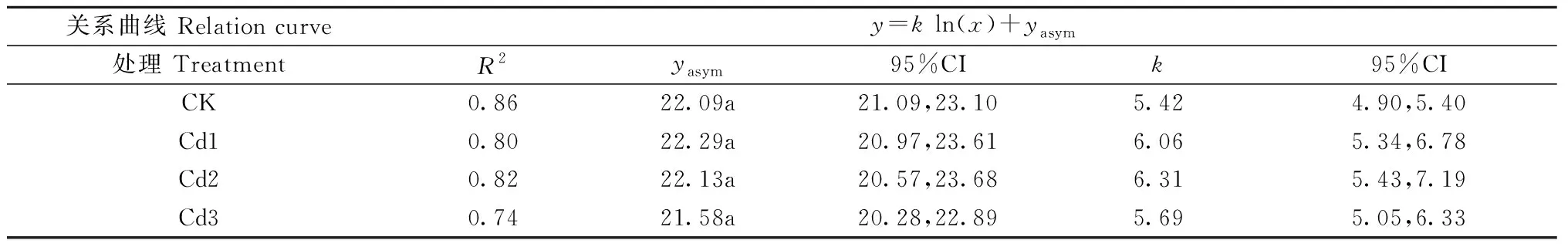
表1 凈光合速率與氣孔導度間關系特征①Tab.1 Nonlinear regression parameters between net photosynthetic rate and stomatal conductance
2.2 鎘對香樟葉綠素含量的影響 由圖3可知,隨著鎘脅迫時間及濃度增加,各類色素和葉綠素a/b并不呈單一的上升或下降趨勢。第15天時,Cd1和Cd3的葉綠素a、葉綠素b及葉綠素總量較CK最高分別顯著降低約29.96%、33.19%和25.76%,而Cd2的各色素差異不顯著。第30天時,鎘脅迫下的葉綠素a和總葉綠素含量較CK分別顯著降低26.46%和16.72%(P<0.05); 與CK相比,Cd1和Cd2的葉綠素b含量分別降低11.70%和18.72%,而Cd3的顯著增加47.65%(P<0.05); Cd3的葉綠素a/b顯著低于CK約60.74%,而其他各處理間無顯著差異。第60天時,與CK相比,不同濃度鎘處理的葉綠素a、葉綠素b和葉綠素總量均不同程度下降,其中Cd3的各類色素含量最低,其葉綠素a、葉綠素b和葉綠素總量分別較CK顯著降低31.04%、38.10%和33.37%(P<0.05); 鎘脅迫處理對葉綠素a/b無顯著影響。以上結果表明,鎘脅迫可以影響香樟葉片葉綠素等色素的合成,進而對光合作用產生一定的影響。

圖3 不同濃度鎘對香樟葉片葉綠素含量的影響Fig.3 Effects of different concentrations of cadmium contaminated on chlorophyll content of C. camphora leaves
2.3 鎘對香樟葉片滲透調節物質的影響 隨著鎘脅迫時間的增加,不同鎘濃度處理的脯氨酸含量呈先下降后上升再下降的趨勢(表2)。第15天時,Cd1、Cd2和Cd3的脯氨酸含量分別比CK高36.57%、11.41%和65.10%,除Cd2與CK差異不顯著外,其他處理間差異顯著(P<0.05)。第30天時,Cd1、Cd2和Cd3的脯氨酸含量分別比CK高8.42%、41.23%和54.38%,除Cd1與CK差異不顯著外,其他處理間差異顯著(P<0.05)。第60天時,Cd3的脯氨酸含量最高,比CK高14.85%(P<0.05),而其他處理與CK間均無顯著差異。隨著鎘脅迫時間的增加,不同濃度鎘處理的可溶性糖含量呈上升趨勢,且不同處理間存在一定差異(表2)。在整個培養期內,Cd3的可溶性糖含量高于其他處理,比CK高43.74%(P<0.05)。第15天時,除Cd2的可溶性糖含量與CK無顯著差異外,其他處理間均有顯著差異(P<0.05)。第60天時,Cd1、Cd2和Cd3的可溶性糖含量分別高于CK約26.61%、29.73%和30.27%(P<0.05),其他處理間無顯著差異。隨著鎘脅迫時間的增加,不同濃度鎘處理的可溶性蛋白含量呈先下降后上升趨勢(表2)。在整個培養期內,Cd2和Cd3的可溶性蛋白含量顯著高于Cd1和CK(P<0.05),且Cd2和Cd3間差異不顯著。第15天和30天時,Cd2和Cd3的可溶性蛋白含量最高分別高于CK約65.43%和73.15%(P<0.05)。以上結果表明,香樟可以通過調控滲透調節物質的合成來適應鎘脅迫環境。

表2 不同濃度鎘對香樟葉片滲透調節物質的影響Tab.2 Effects of different concentrations of cadmium on the osmotica of C. camphora leaves
2.4 鎘對香樟葉片超氧化物歧化酶、過氧化物酶活性和丙二醛的影響 由圖4A可知,隨著鎘脅迫時間的增加,超氧化物歧化酶(SOD)活性總體呈下降趨勢。第15天時,Cd1、Cd2和Cd3的SOD活性顯著低于CK(P<0.05)。第30天時,Cd2和Cd3的SOD活性分別高于CK約26.19%和18.53%(P<0.05),而Cd1的則低于CK約4.07%,Cd3與其他處理間差異不顯著。第60天時,Cd2和Cd3的SOD活性分別高于CK約21.31%、13.48%(P<0.05),CK與Cd1、Cd2與Cd3間的SOD活性無顯著差異。由圖4B可知,隨著鎘脅迫時間的增加,過氧化物酶(POD)活性總體呈上升趨勢,且不同處理的SOD活性均表現為Cd3> Cd2> Cd1> CK。第15天時,Cd1、Cd2和Cd3的POD活性分別高于CK約18.33%、28.09%和65.13%(P<0.05),各處理間差異顯著。第30天時,Cd1、Cd2和Cd3的POD活性分別高于CK約16.49%、39.18%和44.33%(P<0.05)。第60天時,Cd1、Cd2和Cd3的POD活性分別高于CK約24.94%、29.20%和48.60%(P<0.05)。由圖4C可知,整個培養期內Cd2的丙二醛(MDA)含量最高,而CK最低。第15天時,Cd2和Cd3的MDA含量分別高于CK約21.36%和16.94%(P<0.05),但各處理間無顯著差異。第30天時,Cd2的MDA含量高于CK約24.21%(P<0.05),其他處理間無顯著差異。第60天時,Cd2和Cd3的MDA含量分別高于CK約41.75%和34.38%(P<0.05)。以上結果表明香樟可以通過調控抗氧化系統來適應鎘脅迫環境。

圖4 不同濃度鎘對香樟葉片超氧化物歧化酶 (A)、過氧化物酶活性 (B) 和丙二醛 (C) 的影響Fig.4 Effects of different concentrations of cadmium on the activities of SOD (A), POD (B) and MDA content (C) in C. camphora leaves
3 討論
3.1 光合作用參數對鎘脅迫的響應 光合作用與生產力和產量等密切相關(Zhouetal., 2019)。Cd脅迫可通過影響與光合有關的元素吸收與同化、改變色素含量、干擾葉片氣孔開閉行為、影響電子傳遞等,進而影響植物的光合速率,使植物失綠、黃葉、生長減緩。簡敏菲等(2015)研究表明,Cd脅迫下光合氣體交換參數(Pn、Gs、Tr和WUEi)隨著脅迫時間及Cd濃度的增加表現出下降趨勢。本研究發現短時間(15天)鎘脅迫對香樟葉片的Pn、Gs和Ci無顯著影響,而長時間(30和60天)鎘脅迫下的Pn較CK均有不同程度的下降,其中最高濃度鎘處理Cd3的凈光合速率下降最明顯,表明高濃度鎘對香樟光合能力的抑制作用更為強烈。凈光合速率(Pn)下降主要可分為氣孔限制和非氣孔限制2種(Adeletal., 2005),而Ci值的高低可作為判斷氣孔限制和非氣孔限制的主要參考依據,Pn、Gs和Ci同時下降可能是氣孔限制;Pn、Gs下降,Ci增加則表明可能是非氣孔限制所致(祁金洋等, 2018)。本研究Pn隨著Gs的增加呈非線性增加且存在閾值,而不同濃度鎘處理的Pn和Gs間關系無顯著差異。香樟葉片的Pn在鎘脅迫后期隨鎘濃度的增加而降低,這可能是香樟葉綠素含量下降,也可能是因為鎘脅迫引起香樟脫落酸含量增加(Kazninaetal., 2014),迫使Gs下降,導致光合作用受到抑制,進而影響Pn和Gs,使其隨培養時間及鎘濃度的增加逐漸降低。
3.2 鎘脅迫對葉綠素合成的影響 葉綠素是植物葉片進行光吸收、光傳遞和光轉化的場所,在一定程度上是反映植物光合能力受環境脅迫程度的重要指標(王怡霖等, 2019)。不同濃度鎘處理下香樟葉綠素含量均有不同程度的降低,這與楊葉萍等(2016)研究結果相似。可能是香樟在鎘脅迫環境中,吸收了較多Cd2+進入香樟細胞組織內,與葉綠體蛋白質上的羥基配位螯合或與其中的鐵、鎂等離子發生置換,使葉綠體的形態和基粒結構發生變化,葉綠體功能紊亂,生理機制平衡逐漸被抑制作用代替(Adeletal., 2005),導致葉綠素含量下降。培養期內,Cd1處理的葉綠素含量雖低于對照,但隨著鎘脅迫時間的增加有逐漸增加趨勢,可能是低濃度鎘對香樟葉綠素合成的影響程度較弱,香樟逐漸對鎘脅迫產生一定的適應性,香樟可通過消耗更多能量或與其他抗氧化酶協同維持機體內的生理代謝平衡。在較長時間高濃度鎘脅迫下會導致香樟光合色素及光合作用下降,但是香樟通過調控Pn和Gs等生理特性以適應或降低鎘脅迫的影響,增強自我調節機制。
3.3 鎘脅迫對香樟葉片滲透調節物質的調控 植物受重金屬脅迫時,其生理生化特性可反映植物的應對機制。脯氨酸、可溶性糖和可溶性蛋白作為植物細胞內重要的滲透調節物質,其累積與植物體內過量活性氧自由基的清除及膜脂過氧化傷害的減輕密切相關(張麗萍等, 2016; Batooletal., 2014; Sinishaetal., 2018),在植物逆境脅迫中發揮重要作用。本研究發現,無論高還是低濃度的鎘脅迫,香樟幼苗葉片的脯氨酸、可溶性糖含量和可溶性蛋白含量均有一定程度增加,且高濃度鎘處理Cd3的各物質含量最高,這與Ali等(2014)研究植物體內可溶性蛋白含量會隨鎘濃度增加而降低不同,可能與不同鎘濃度的變化有關。各調節物質含量的提高可能是在鎘脅迫下香樟葉片通過自身的調控機制,如螯合、區化等作用產生更多的脯氨酸、可溶性糖和可溶性蛋白,保持細胞原生質滲透勢和膨壓,緩解逆境脅迫造成的植株細胞脫水或水分代謝異常,以維持植株的正常生長發育。陳良華等(2017)研究認為,10 mg·kg-1鎘處理下脯氨酸含量增加,有利于平衡美洲黑楊(Populusdeltoides)雌、雄株葉肉細胞內的滲透勢,減緩葉肉細胞受到的氧化危害。此外,陸秀君等(2008)研究也發現,長時間鎘脅迫可誘導刺槐(Robiniapseudoacacia)葉片大量形成脯氨酸、可溶性糖和可溶性蛋白,以提高植物的抗逆性。
3.4 鎘脅迫對香樟葉片抗氧化系統的影響 SOD是植物體內酶促防御系統中重要的保護酶之一,有助于將氧自由基團轉化為H2O2和O2,減輕自由基對植物體細胞的毒害(胡國濤等, 2016; 楊波等, 2018; Yilmazetal., 2018)。本研究結果表明,鎘脅迫前期(15天),不同濃度鎘處理的SOD活性低于CK,而在鎘脅迫后期,中高濃度鎘處理的SOD活性高于低濃度鎘處理和CK,由此認為較長時間高濃度鎘脅迫會誘導香樟提高SOD活性來清除自由基,以維持香樟正常生長機能。這與湯葉濤等(2010)研究發現滇苦菜(Picrisdivaricata)SOD活性隨著鎘濃度的增加而提高有類似之處。POD具有去除過氧化氫和酚類等毒性物質的重要功能(張軍等, 2018)。POD活性隨著鎘脅迫時間及鎘濃度的增加而增加,可能是受鎘脅迫后,更多的鎘離子進入香樟組織,使其組織內有毒害作用的H2O2不斷累積增加,從而誘導植物體POD活性增加,以抵御外界不良環境。MDA是植物細胞膜脂過氧化物,其含量的高低反映了植物細胞膜結構受損傷的程度(Lluganyetal., 2013)。本研究結果表明,在整個鎘脅迫期內,中高濃度鎘處理(Cd2和Cd3)的MDA含量高于CK,在脅迫60天時影響最為顯著,這與Stoláriková-Vaculíková等(2015)研究結果相似。長時間高濃度鎘脅迫對香樟葉片細胞膜系統具有損害作用,而香樟會通過調節體內SOD和POD活性來消除毒害物質,是香樟耐鎘的主要機制之一。關于高濃度鎘脅迫機制及調節物質的應答機制還有待深入研究。
本研究結果表明,在一定濃度的鎘污染土壤中,香樟可通過調控光合作用、抗氧化保護酶系統和滲透調節物質等生理生化過程適應鎘污染環境,維持其正常生長。但本研究僅開展了短期(60天)鎘脅迫下香樟的生理生化特性研究,可能是一些短期現象,更長時間鎘脅迫對香樟的生理生化影響是否相似,有待后期長期深入監測研究,以期為香樟用于重金屬污染場地的生態修復提供理論依據。
4 結論
香樟葉片的凈光合速率、葉綠素、滲透調節物質、抗氧化酶系統和MDA等生理生化特征對不同濃度鎘的響應存在一定差異。短時間內各濃度鎘處理香樟葉片的凈光合速率(Pn)無明顯變化,較長時間鎘脅迫會導致Pn降低,且鎘濃度越高,葉片的Pn越低。鎘脅迫會導致香樟的葉綠素含量出現不同程度的降低,而使香樟葉片的脯氨酸、可溶性糖含量增加,且以高濃度鎘處理Cd3最高。培養后期(30和60天),SOD和POD活性顯著上升,可減輕逆境對香樟葉片膜系統造成的損傷。香樟在重金屬鎘脅迫下能保持較好生長,因此香樟對鎘具有較強耐性,具備修復重金屬污染土壤的潛力。
