長江中游鳙群體的微衛星遺傳多樣性分析
2020-07-27 09:15:34羅相忠鄒桂偉梁宏偉
淡水漁業 2020年4期
沙 航,羅相忠,鄒桂偉,梁宏偉
(中國水產科學研究院長江水產研究所,武漢 430223)
鳙(Aristichthysnobilis)俗稱花鰱,是我國重要的大宗淡水養殖魚類之一。2018年全國養殖產量達309.64萬噸,僅次于草魚和鰱[1]。隨著水利設施的興建,洄游通道和產卵場被破壞,鳙的自然群體遭受到了極大沖擊,種群群體數量急劇下降,成為長江主要魚類中數量減少最顯著的種類之一[2]。與此同時,人工養殖個體逃逸以及遺傳資源管理的不科學也對自然群體遺傳多樣性造成一定的擾動[3,4]。為了恢復長江水生生物多樣性和漁業資源,近年來開展了大規模的增殖放流,鳙等四大家魚親本放流工作也取得了良好的效果[5]。科研工作者先后利用同工酶、RAPD、線粒體DNA和微衛星等技術對長江鳙群體的遺傳多樣性進行了分析,長江中下游鳙具有豐富的遺傳多樣性[6],人工繁殖群體的遺傳多樣性明顯低于自然群體[7],而放流群體與天然群體的遺傳多樣性相差不大[8]。長江中游江段及其一級支流是鳙重要的自然產卵場,是維持長江中游鳙種質資源的重要場所,持續長期監測長江中游鳙自然群體的遺傳結構和遺傳多樣性,對評估長江中游鳙種質資源現狀以及制定科學的增殖放流漁業管理措施具有重要意義。本研究采用微衛星分子標記對長江水系中游的3個鳙群體的遺傳本底進行分析,以期為今后保護和合理利用種質資源提供參考。
1 材料與方法
1.1 樣本來源與基因組DNA提取
3個鳙群體于2018年5月分別采自石首老河國家四大家魚原種場(石首,SS),監利老江河國家四大家魚原種場(監利,JL)和湖南魚類原種場(長沙,CS),石首和監利原種場采集的鳙樣本均為繁殖季節在長江采集的幼苗經原種場培育而成的1齡鳙,湖南魚類原種場的鳙采自湘江。每個群體隨機采集30尾個體,共90尾。鰭條經無水乙醇固定后置于-20 ℃保存。取0.2 g鰭條組織經無菌水漂洗后置于2 mL離心管烘干剪碎,用高鹽法提取基因組DNA[9],經1%瓊脂糖電泳檢測后,于-20 ℃冰箱保存備用。
1.2 微衛星引物及PCR擴增檢測
采用本課題組自行開發的12對微衛星引物,并用Fam熒光基團修飾上游引物的5′端(表1),引物由武漢天一輝遠生物科技有限公司合成。PCR反應體系為25 μL,包含2×Taq PCR Mix 12.5 μL,10 μmol/L上下游引物各1 μL,50 ng/μL模板DNA 1 μL,dd H2O 9.5 μL。反應條件為94 ℃預變性5 min,94 ℃ 15 s,53~57 ℃退火30 s,72 ℃ 30 s,共35個循環,72 ℃延伸5 min,4 ℃保存。PCR擴增產物在ABI Prism3730 XI 測序儀進行毛細管電泳,采用DataCollection 軟件和GeneMarker V2.2.0軟件采集和分析數據并輸出等位基因掃描圖譜,然后分析各個位點的基因型。

表1 鳙12對微衛星引物信息Tab.1 Primers information of 12 microsatellite markers of A.nobilis
1.3 數據處理
用Popgene32軟件分析各個微衛星位點在3個鳙群體的等位基因數(Na)、有效等位基因數(Ne)、觀測雜合度(observed heterozygosity)、期望雜合度(He)、Shannon信息指數(I),并計算群體間的Nei′s遺傳距離。Hardy-Weinberg平衡偏離指數(D)按以下公式計算:D=(Ho-He)/He。基于群體間的Nei′s遺傳距離用MEGA X軟件繪制NJ聚類圖。用Arlequin3.1軟件計算兩兩群體間的遺傳分化指數(Fst),采用分子方差分析(AMOVA)計算群體的遺傳結構,通過1 000次模擬重復抽樣檢測群體間Fst值的顯著性[10]。用Cervus3.2軟件計算各位點的多態信息含量(PIC)。
2 結果分析
2.1 微衛星位點的多態性
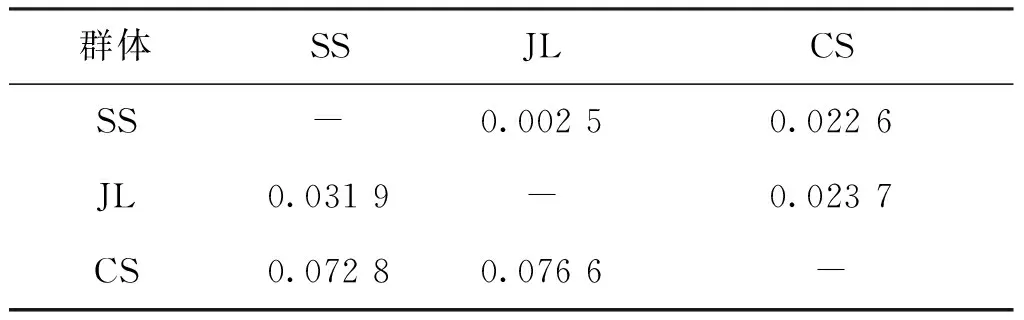
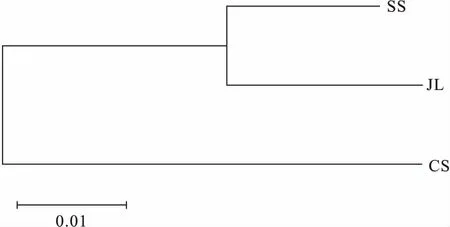
12對微衛星引物在SS、JL和CS 3個群體中共檢測到117個等位基因,3個群體中Na介于5~19,Ne為1.731~11.567,平均Na和Ne分別為9.750和4.050(見表2)。其中Ans055和Ans196位點Na最多(19個),Ans014和Ans119位點Na最少(5個)。12個位點Ho為0.398~0.778,He為0.425~0.919,其中11個位點偏離Hardy-Weinberg平衡(D<0)。I為0.782~2.588。PIC為0.371~0.907,平均PIC為0.618。12個位點中有8個位點為高度多態位點(PIC>0.5),4個位點為中度多態位點(0.25 表2 鳙12個微衛星位點的遺傳多態性Tab.2 Genetic diversity of 12 microsatellite loci in A.nobilis 石首、監利和長沙 3個鳙群體遺傳多樣性見表3。3個群體平均Na介于5.500~8.500,平均Ne為3.294~4.244,平均Ho和平均He分別介于0.557~0.582和0.620~0.730,其中長沙群體的平均Ne、平均He和I均最高。從PIC上看,長沙群體的遺傳多樣性最高,而石首群體的遺傳多樣性相對較低,整體上3個群體的PIC較高(PIC>0.5),遺傳多樣性較豐富。 表3 鳙3個群體的遺傳多樣性Tab.3 Genetic diversity among the three populations of A.nobilis 3個鳙群體間的Nei′s遺傳距離和遺傳分化指數見表4。3個群體間的遺傳距離為0.031 9~0.076 6,監利與長沙群體遺傳距離最大(0.076 6),石首和監利群體間的遺傳距離最小(0.031 9)。群體間遺傳分化指數為0.002 5~0.023 7,其中石首、監利和長沙3個群體兩兩之間的遺傳分化指數均小于0.05,三個群體之間遺傳分化程度較低。基于群體間Nei′s遺傳距離的NJ聚類結果如圖1所示,石首和監利群體先聚為一支,然后與長沙群體聚為一支。AMOVA分析顯示(表5)3個群體中群體內的遺傳變異占總體變異的95.60%,群體間的遺傳變異為4.40%,3個群體間遺傳分化指數為0.044(Fst<0.05),表明群體間的遺傳變異主要來自群體內的個體之間。 表4 鳙3個群體間的遺傳距離(對角線下)和遺傳分化指數(對角線上)Tab.4 The genetic distance (below diagonal) and genetic differentiation indices (above diagonal) among the three populations of A.nobilis 圖1 基于遺傳距離構建的3個鳙群體的NJ聚類樹Fig.1 NJ clustering tree of tht three populations of A.nobilisbased on Nei′s genetic distance 表5 鳙3個群體的AMOVA分析Tab.5 AMOVA analysis among the three populations of A.nobilis 群體的遺傳多樣性和遺傳潛力可以用等位基因數、雜合度和多態信息含量等遺傳參數來衡量[11]。雜合度是反映群體遺傳變異程度的重要參數,雜合度越大遺傳多樣性越高[12]。本研究中的SS(He=0.633)、JL(He=0.651)和CS(0.730)3個群體的雜合度均高于朱曉東[13](石首, He=0.287 6;監利,He=0.321 4)和田華[14](監利,He=0.550 2)的研究結果,與朱傳坤[15]的研究結果較為接近(石首老河,He=0.661)。長沙群體的多態信息含量(0.606)高于單淇[16]的研究結果(0.557),而雜合度略低。基于Botstein[17]的群體遺傳多樣性判別標準,本研究的3個群體的遺傳多樣性均較高(PIC>0.5)。目前長江中游3個群體的遺傳多樣性仍保持在較高水平。遺傳多樣性與群體的大小呈一定的正相關。鳙的自然群體中以長江群體數量最大,基因庫最豐富[18]。雖然其經歷高強度捕撈、產卵場破壞、棲息環境惡化等,但近十年來隨著國家和地方對漁業資源的保護,鳙的增殖放流活動持續開展,其遺傳多樣性并未發生持續性下降。 本研究中石首、監利和長沙3個群體大多數位點表現為雜合子缺失(D<0)。耿波等[19]在對鳙的四川和江西群體研究中也發現存在雜合子缺失現象。關于雜合子缺失的形成原因目前仍存在爭議,可能與研究群體樣本量、種群退化、性別比例失衡和無效等位基因有關[20,21],就本研究而言可能是由于分析的樣本數量較少或位點上存在無效等位基因導致。傅洪拓等[22]對長江不同江段日本沼蝦(Macrobrachiumnipponense)的研究中發現,日本沼蝦82.5%的群體位點表現為雜合子缺失,推斷其主要是無效等位基因導致的。在青石斑魚(Epinephelusawoara)[23]、大鱗副泥鰍(Paramisgurnusdabryanus)[24]和擬穴青蟹(Scyllaparamamosain)[25]等微衛星分析中也存在由無效等位基因導致的雜合子缺失現象。 遺傳距離是衡量群體間遺傳分化的重要指標,群體間遺傳距離越小,遺傳分化程度越低,群體間親緣關系越近[26]。固定指數(Fst)通常被用來衡量群體間的遺傳分化程度,Fst<0.05時,群體間遺傳分化較小;0.05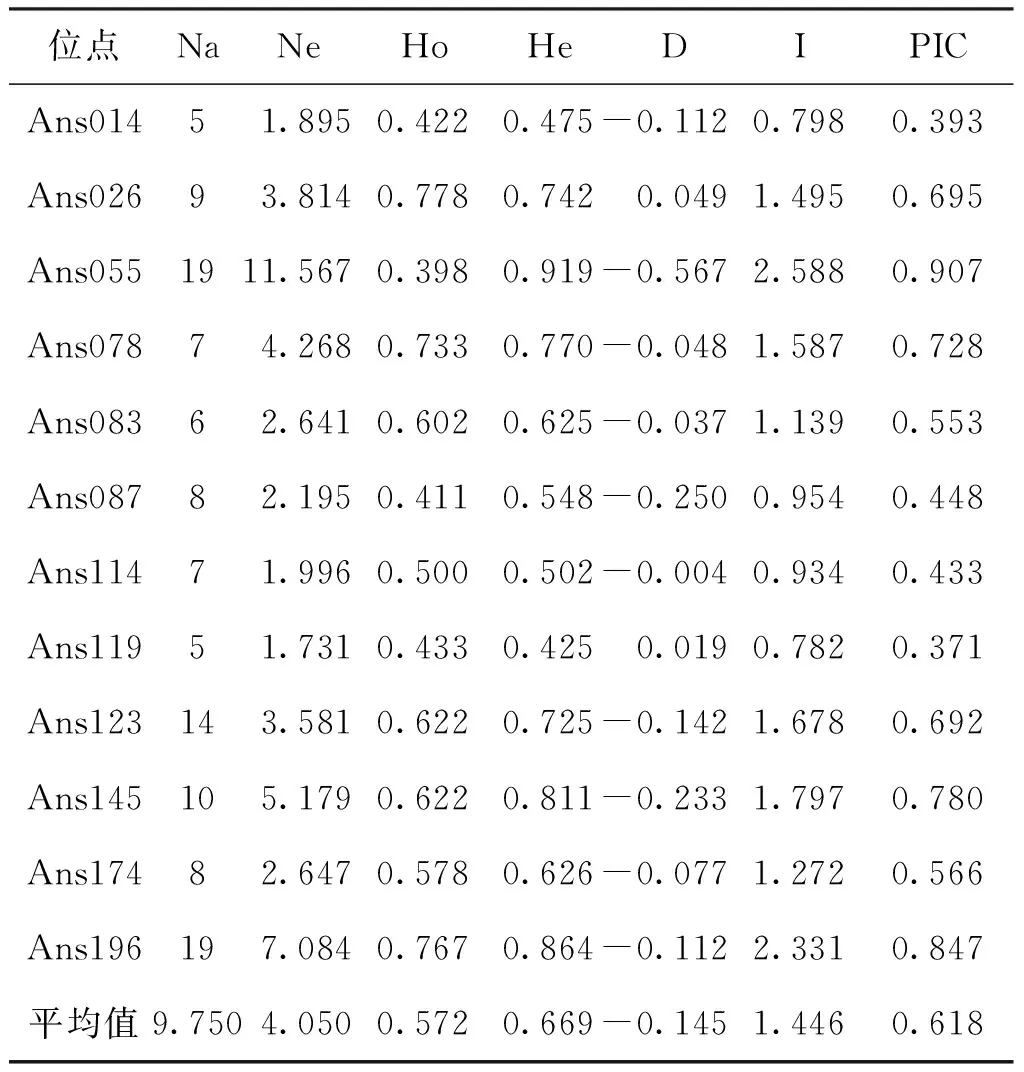
2.2 鳙3個群體的遺傳多樣性

2.3 鳙3個群體間的遺傳分化

3 討論
3.1 鳙群體的遺傳多樣性
3.2 鳙3個群體的遺傳結構
