不同金屬離子對稻田自然生物膜磷酸酶活性的影響①
2020-07-25 03:05:50蔡述杰鄧開英李九玉徐仁扣
土壤 2020年3期
關鍵詞:影響
蔡述杰,鄧開英,李九玉*,徐仁扣
不同金屬離子對稻田自然生物膜磷酸酶活性的影響①
蔡述杰1, 2,鄧開英1,李九玉1*,徐仁扣1
(1土壤與農業可持續發展國家重點實驗室(中國科學院南京土壤研究所),南京 210008;2中國科學院大學,北京 100049)
以稻田自然生物膜為研究對象,研究不同濃度金屬離子對生物膜酸性和堿性磷酸單酯酶(ACPase和ALPase)活性與酶反應動力學參數的影響。發現K+和Na+在0 ~ 1.0 mmol/L濃度下均對ACPase和ALPase無明顯影響,Ca2+和Mg2+對ACPase和ALPase活性均有激活效應,可使磷酸酶活性最大提高14.1% 和46.7%;Zn2+、Cu2+、Mn2+、Al3+和 Ag+對ACPase和ALPase活性均有抑制作用。Co2+在低濃度時對ACPase和ALPase活性有一定的激活作用,高濃度時變為抑制作用;Cr6+在0.1 mmol/L時對ACPase活性有促進作用,而在離子濃度0.25 ~ 1 mmol/L的范圍內,均對ACPase活性產生抑制作用。通過酶反應動力學分析發現,Mg2+的加入使ACPase與底物的親和力與催化效率增強,Cu2+對ACPase和ALPase的抑制作用主要是非競爭性抑制作用,Zn2+雖然提高ACPase與底物的親和力,但是降低了ALPase與底物的親和力與催化效率。金屬離子對酶活性的影響因金屬離子種類、濃度和磷酸酶類型而異,主要通過與酶活性位點或底物結合,改變酶的活性、對底物的親和力,以及影響酶基因的表達等實現的。研究結果將為評估自然生物膜在稻田磷的生物地球化學循環中的作用,以及稻田受重金屬污染的可能風險提供理論依據。
自然生物膜;磷酸酶活性;金屬離子;酶動力學常數;稻田
稻田水土界面之間的微生物通常以稻田自然生物膜的形式存在。自然生物膜是在自然環境條件下形成于淹水基質表面的微生物聚集體及其交織的非生物物質的集合體。微生物包括細菌、真菌、原生動物、藻類、病毒等;非生物物質包括微生物分泌的多糖、蛋白質、脂類等有機質以及鐵錳氧化物、碳酸鹽、硅酸鹽等無機礦物質[1]。稻田自然生物膜覆蓋于土壤表面,是外源污染物如重金屬離子進入土壤的必經之地,是多種物質包括養分、重金屬重要的存儲庫以及傳輸介質[2]。大量研究發現自然生物膜是水體中重要的短期磷庫,也對富營養化水體-底泥界面無機磷的緩存和緩釋起著重要的調控作用[3]。而且,自然生物膜,作為一種微生物聚集體,還可以分泌多種水解酶,對土壤和底泥中有機磷的降解和生物有效性等起著促進作用[4]。
當前,由于工業廢物排放、污水灌溉以及含有重金屬的農藥、化肥的不合理使用等,許多污染物包括過量的重金屬離子不斷進入稻田生態系統。這些重金屬污染可對稻田水土界面中的微生物產生毒害,阻礙了碳氮循環、代謝活性、酶活性等各種微生物過程[5]。自然生物膜在水環境中對重金屬的遷移、生物地球化學特性、生物可利用性和毒性等過程中,均起著非常重要的作用[6]。關于自然生物膜和金屬之間的相互作用進行了較多的研究,Dong等[7-9]發現自然生物膜對Pb、Cd、Cu、Zn等重金屬有很強的吸附作用。Serra等[10]報道了河流系統中自然生物膜群落對水體金屬Cu暴露的響應,分析表明自然生物膜中重金屬含量如Cd (II)和水體Cd (II)濃度的變化一致,具有較高的相關性。一些研究也表明水體中存在許多光合物種,如藍藻具有強烈的吸收金屬元素的能力。這些研究均表明自然生物膜具有較大潛在富集水體中金屬元素的能力[11]。但重金屬離子對稻田自然生物膜的磷酸酶活性的影響研究還極少涉及。
自然生物膜包含多種磷酸酶,如磷酸單酯酶、磷酸二酯酶、聚磷酸酶等,其中磷酸單酯酶活性最高、分布最廣泛。已有研究證明覆蓋于淹水基質表面的自然生物膜能夠釋放磷酸單酯酶來水解環境中的有機磷,這是提高環境中磷有效性的重要途徑[12]。磷酸單酯酶是廣泛存在于微生物中的一種金屬酶,其構象及活力受到金屬離子的影響[13],而且金屬離子對磷酸酶基因的表達也受其種類和濃度等影響[14]。目前關于金屬離子對于稻田自然生物膜磷酸酶活性影響的研究很少,因此本文研究了部分金屬離子對稻田自然生物膜酸性磷酸單酯酶(ACPase)和堿性磷酸單酯酶(ALPase)活性及酶動力學參數的影響,以期揭示金屬離子與自然生物膜磷酸酶的作用機制,為稻田磷的生物地球循環過程的認識和重金屬污染的風險評估提供理論依據。
1 材料與方法
1.1 自然生物膜富集與培養
在南京江寧區水稻田采集土壤溶液于自制玻璃缸(長×寬×高=30 cm × 20 cm × 50 cm)中,并將纖維載體淹沒其中。每周添加BG-11營養液[15],以充分維持生物膜的生長。將水缸置于人工氣候室中,光/暗周期為14 h(26 °C)/10 h(20°C),光強度為12 000 lux,濕度為80%。一個月后,微生物聚集體附著在載體上,形成穩定的生物膜。從玻璃缸中收集在纖維載體上附著的生物膜,并用于后續試驗。采用光學顯微鏡(Nilcon-80i,Japan) 和激光共聚焦掃描顯微鏡 (CLSM,Zeiss-LSM710,German) 對周生生物膜的形態進行表征。
1.2 試驗設計
用磷酸單酯酶標準底物對硝基苯基磷酸二鈉鹽(pNPP)來測定酶活性。不同金屬離子的選取如下:K+、Na+、Ca2+選取相對應的可溶性氯化鹽,Mg2+、Co2+、Al3+、Mn2+、Ni2+和Zn2+選取可溶性硫酸鹽,Cu2+和Ag+選取可溶性硝酸鹽, 用K2Cr2O7作為Cr6+的來源。Tris–HCl緩沖溶液被用于控制體系pH。將100 ml生物膜懸浮液在15 ℃下1 157 g離心10 min后移除上清液,用0.05 mol/L Tris–HCl緩沖液(酸性條件為pH 6.0,堿性條件為pH 9.0)重新懸浮生物膜樣品,離心和洗滌過程重復兩次,盡可能減少培養基質中本身攜帶離子的含量。將生物膜再懸浮于100 ml的0.05 mmol/L Tris-HCl緩沖液中,用于后續試驗。
1.2.1 不同離子對磷酸酶活性的影響 依據稻田土壤中各種離子的有效態含量以及重金屬污染對土壤酶活性的影響研究[16],在50 ml離心管中加入不同濃度的金屬離子溶液5 ml,控制金屬離子最終濃度為0、0.1、0.25、0.5、1.0 mmo/L,以測試其對磷酸酶活性的影響。按照相當于0.4 g/L干物質量的比例加入清洗后的生物膜懸液2 ml,統一用水定容至12 ml后在25 °C,200 r/min 的避光條件下振蕩2 h。然后加入5 ml的0.2 mol/L Tris- HCl緩沖液和2 ml的10 mmol/L pNPP溶液(終濃度為1 mmol/L),在37 °C、200 r/min 的避光條件下繼續振蕩1 h, 立即加入1 ml的0.5 mol/L NaOH終止水解反應。過濾后將濾液在405 nm下測定吸光度。每個處理設定3個重復。以不加上述金屬離子體系中的生物膜酶活性為100,其余條件下測定的酶活性與未加金屬離子條件測得酶活性相比得到相對酶活性。
1.2.2 不同離子影響下磷酸酶活性的動力學分析 選取不同濃度的Mg2+、Zn2+和Cu2+(0、0.25、0.5、1.0 mmol/L)溶液,測定其對磷酸酶動力學參數的影響,pNPP濃度范圍為0 ~ 1 500 mmol/L。酶的催化速度隨著底物濃度的增加而增加,直到接近最大速度(max)的某一點。酶催化的反應動力學通常用符合米氏方程(Michaelis–Menten方程),其中米氏常數(m)是速度達最大速度一半時的底物濃度;米氏常數表明酶對底物的親和力:m越低,親和力越高。這些參數由Michaelis–Menten方程描述:

式中:是任意時刻酶催化反應的速度(μmol(g·h));是相應時間的底物濃度(mmol/L);max是最大催化反應速率(μmol(g·h));m是米氏常數(mmol/L)。動力學參數max和m使用Michaelis–Menten方程的線性變換 Hanes–Woolf 方程計算:

1.3 統計分析
本研究每組試驗重復 3 次,結果以平均值±標準誤差表示。采用 Microsoft Excel 2016 以及數據進行統計、制圖;采用 SPSS 中 Duncan 法對數據進行差異顯著性分析,< 0.05 表示差異顯著。
2 結果與討論
2.1 自然生物膜形態表征
采用光學顯微鏡(Nilcon-80i,Japan)和激光共聚焦掃描顯微鏡(CLSM,Zeiss-LSM710,German)對自然生物膜的形貌進行了表征(圖1)。結合文獻[17]分析,發現自然生物膜主要由光自養微藻,尤其是絲狀藻類組成。絲狀藻類交織在一起,很多細菌、真菌、原生動物和小型多細胞動物等附著在絲狀藻類上面。

(A. 光學顯微鏡(400×);B. 激光共聚焦掃描顯微鏡)
2.2 不同金屬離子對生物膜磷酸酶活性的影響
金屬離子對自然生物膜中ACPase和ALPase活性影響的結果表明,影響的方式有4種:抑制型、激活型、相對無害型和混合型(表1)。正一價堿金屬離子(K+和Na+)在0 ~ 1.0 mmol/L濃度范圍內均對ACPase和ALPase無明顯影響(>0.05),屬于相對無害型。Ca2+可以提高ACPase和ALPase的活性,在0.1 ~ 1.0 mmol/L濃度范圍內最大可提高磷酸酶的活性達14%(<0.05)。Mg2+對ACPase和ALPase活性均有顯著的激活效應,隨著離子濃度的增大,ACPase和ALPase活性不斷增強;在Mg2+濃度1.0 mmol/L時,酶活性最大可提高54%,屬于激活型。Zn2+、Cu2+、Mn2+、Al3+對ACPase和ALPase活性均有顯著抑制作用(<0.05),屬于抑制性;抑制劑濃度達到1.0 mmol/L時,最大抑制率可達85%。但是這些離子對于ACPase和ALPase活性的抑制程度不同,Cu2+、Al3+對ACPase活性的影響明顯大于ALPase,而Zn2+對ALPase活性的影響明顯大于ACPase。Ni2+、Ag+在低濃度(0.1 mmol/L)時對ACPase活性無明顯影響,隨著離子濃度的增加,ACPase和ALPase活性降低。Co2+在低濃度時對ACPase和ALPase活性有一定的激活作用,隨著濃度的增大,變為抑制作用,屬于混合型。Cr6+在低濃度(0.1 mmol/L)時對ACPase活性有促進作用,而在離子濃度0.25 ~ 1.0 mmol/L的范圍內,均對ACPase活性產生抑制作用,當Cr6+離子濃度達到1.0 mmol/L時,生物膜已喪失ACPase活性。而Cr6+對ALPase活性具有激活作用,隨著離子濃度的增大,ALPase活性不斷增加。

表 1 不同金屬離子對生物膜磷酸酶活性的影響
注:同行數據小寫字母不同表示不同濃度金屬離子對ACPase或ALPase相對活性影響達<0.05顯著水平。
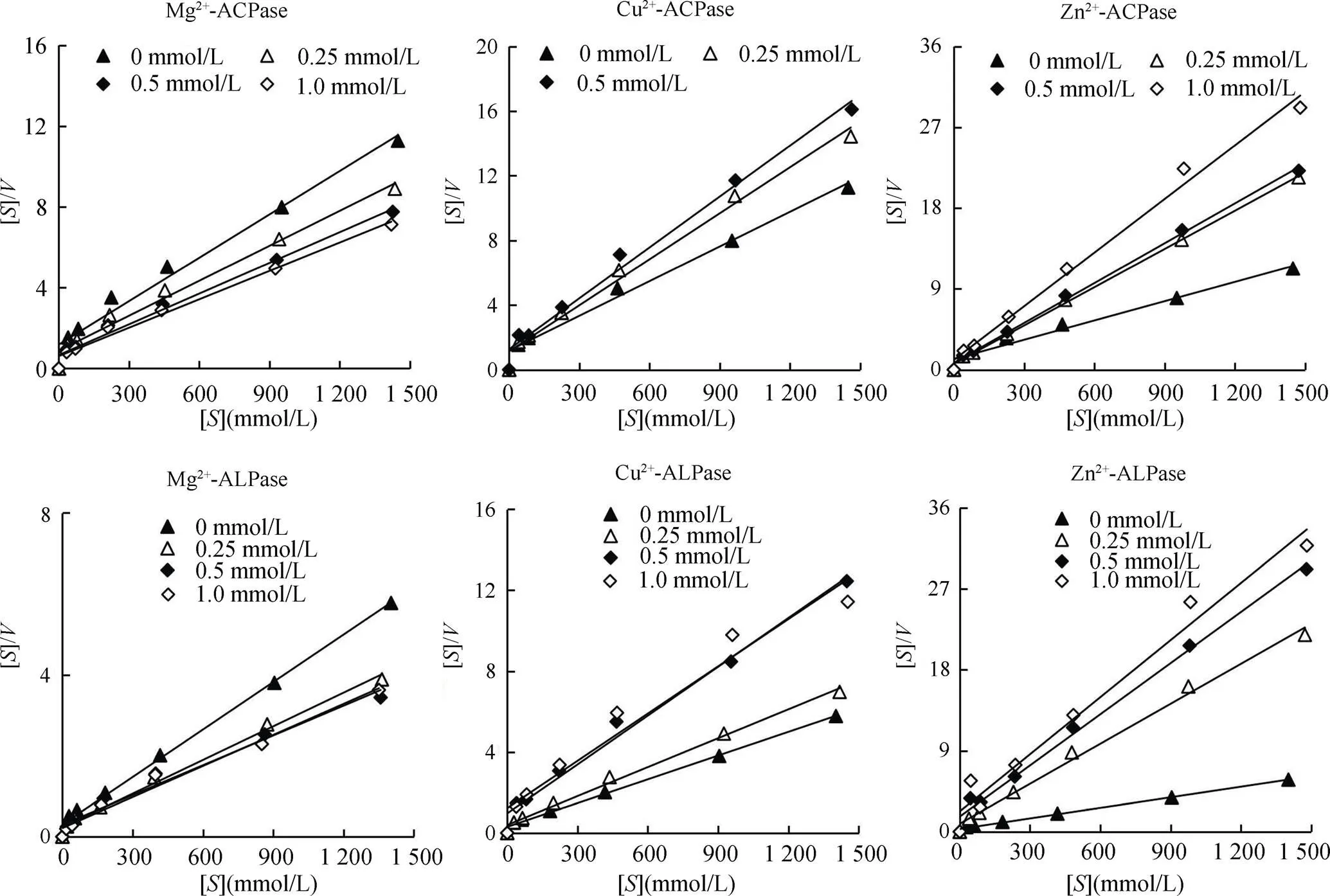
2.3 不同金屬離子對生物膜磷酸酶反應動力學的影響
磷酸酶是一種金屬酶,其酶活性的充分發揮需要兩種金屬離子:Mg2+和Zn2+,其中Zn2+位于酶活性中心,是酶活性必需基團之一,Mg2+對酶活性的表達起到重要作用[18];而Cu2+是土壤污染的幾種主要重金屬之一。所以選取Mg2+、Zn2+和 Cu2+對磷酸酶的動力學常數(m和max)進行探討。圖2中顯示了在Mg2+、Cu2+和Zn2+存在下,酶反應動力學變形方程的Hanes–Woolf曲線,[] /和[] 擬合方程的截距和斜率與不添加金屬離子的處理不同,說明金屬離子對生物膜磷酸酶活性產生了影響。擬合得出的m和max值表明(表2),無金屬離子加入下自然生物膜ACPase的m值為164.58 μmol/L,max值為pNP 138.89 μmol/(g·h),而ALPase的m值為76.90 μmol/L,max值為pNP 256.41 μmol/(g·h)。可知自然生物膜ALPase與底物的親和力與催化效率都強于ACPase。隨著Mg2+濃度的增加,ACPase的m值降低,max值增大,說明Mg2+的加入使ACPase與底物的親和力與催化效率增強;而ALPase的m值變化不大,max值增大,說明Mg2+的加入主要使ALPase催化效率增強。Cu2+的加入(1.0 mmol/L條件下ACPase完全失活,無法擬合)對ACPase的m值無明顯變化,而max值不斷下降,所以Cu2+對ACPase的抑制作用可認為是非競爭性抑制作用;在低濃度條件下(0 ~ 0.25 mmol/L),Cu2+的加入對ALPase的m值無明顯變化,而max值下降,也可認為是非競爭性抑制作用,而在高濃度范圍下(0.25 ~ 1 mmol/L),m值增大,max值降低,所以Cu2+在高濃度下同時降低了ALPase的親和力和催化效率,屬于混合型抑制作用。Zn2+的加入均降低了ACPase的m值和max值,表明Zn2+雖然提高ACPase與底物的親和力,但是降低了ACPase的催化速度。在低濃度范圍(0 ~ 0.25 mmol/L),Zn2+對ALPase的作用效果與ACPase類似,同時降低了m值與max值。在較高濃度范圍(0.25 ~ 1 mmol/L),隨著Zn2+濃度的升高,m值升高,max值降低,Zn2+同時降低了ALPase與底物的親和力與催化效率。
3 討論
由圖1 可以看出,自然生物膜主要是絲狀藻等光合藻類為主導的物種相互纏繞而構成自然生物膜的基質,而其他生物棲息于此基質上。許多微生物共存于自然生物膜中,意味著自然生物膜是一個有別于單一微生物種群的復雜微生態系統。在自然生物膜這種非均相體系中,金屬離子、膠體、微生物和酶之間的復雜反應都有可能干擾任何金屬離子對于磷酸酶的影響。已有研究證明自然生物膜通過本身這種復雜特殊的組成結構可能使其在抵抗外源污染物時,具有有別于簡單微生物種群的性質,由于自然生物膜能夠承受一定的金屬離子脅迫而不會嚴重喪失其生物化學活性[19]。一些關于稻田自然生物膜的研究表明,自然生物膜能夠適應一定濃度的重金屬污染,而胞外聚合物的吸附、群落中藻類豐度提高和組成變化導致碳代謝活性功能的響應,是自然生物膜能夠適應重金屬脅迫的主要原因[20]。本文中生物膜磷酸酶活性對重金屬的敏感性遠低于麥芽中磷酸酶的,如0.5 mmol/L的Cu2+和Zn2+抑制生物膜酸性的活性約40%,而同樣濃度的Cu2+和Zn2+使麥芽中磷酸酶的活性降低80% 以上[21]。活性污泥中微生物和介質環境更加復雜,因此0.5 mmol/L的Cu2+和Zn2+僅使活性污泥的磷酸酶活性降低約15%[22]。
(V:任意時刻酶催化反應的速度(μmol(g·h);[S]:相應時間的底物濃度)
表 2 不同濃度Mg2+、Zn2+和Cu2+作用下ACPase和ALPase動力學常數max(μmol/(g·h))和m(μmol/L)的變化
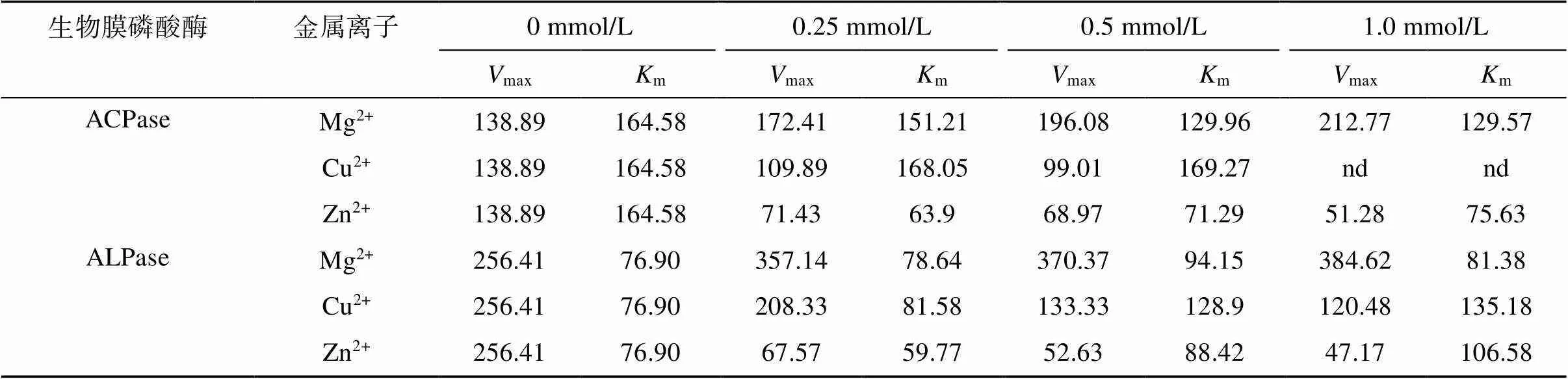
Table 2 Changes of kinetic constants (Vmax and Km) of ACPase and ALPase under different concentrations of Mg2+, Zn2+ and Cu2+
注:nd表示未測定。
金屬離子對酶活性的影響可表現為對酶蛋白的作用,一般分為 3 種類型[16]:①一些金屬離子的加入可以促進酶活性中心與底物間的配位結合,改變酶蛋白的表面電荷以及催化反應的平衡性質,從而增強酶活性,即有激活作用。②一些金屬離子占據了酶的活性中心或與酶分子的巰基、胺基和羧基結合,導致酶活性降低,即有抑制作用。③重金屬與酶沒有專一性對應關系,對酶活性影響小。而且,金屬離子還會影響磷酸酶基因的表達[14]。如細菌堿性磷酸酶的種類多樣,根據堿性磷酸酶蛋白序列的相似度和底物的不同,可主要分為 3 種類型:和。由于 Zn2+與 Mg2+是堿性磷酸酶的輔基,一定濃度的 Zn2+與 Mg2+的存在會促進細菌基因的表達;而一定濃度的 Ca2+和 Co2+能重新促進基因的表達[14]。因此,一定濃度的某些金屬離子通過促進磷酸酶基因的表達,促進磷酸酶活性,但這種作用與金屬離子的種類、濃度和磷酸酶基因的類型有關。但濃度過高時,則會因毒害作用而抑制酶活性表達。
我們的研究表明,正一價堿金屬離子(Na+和 K+)對磷酸酶活性無明顯影響,這一發現與 Chen 等[23]的發現一致,說明 Na+和 K+對稻田自然生物膜磷酸酶沒有對應關系。Mg2+促進了ACPase 和 ALPase 酶活性,因為其是磷酸酶中心結合位點的調節離子,加入能夠促進酶活性中心與底物間的配位結合,來改變酶催化反應的平衡性和酶蛋白的表面電荷,從而可增強酶活性[18]。Mg2+也是磷酸酶的輔基,并促進磷酸酶功能基因的表達[14],因此在 0 ~ 1.0 mmol/L 范圍內顯著提高了酸性和堿性磷酸酶活性。Ca2+、Co2+在一定條件下也能夠提升酶活性,Wu 等[24]研究發現加入一定濃度 Ca2+、Co2+后能促進磷酸酶基因的表達。但Ca2+、Co2+其對磷酸酶活性的激活作用明顯低于 Mg2+。Cr6+在酸性條件下抑制磷酸酶活性,在堿性條件下激發磷酸酶活性,這可能與 Cr6+在不同 pH 條件下 Cr6+的離子形態及其對藻類的毒性效應不同有關[25]。Zn2+、Cu2+、Mn2+、Al3+、Ag+對磷酸酶表現出了抑制作用,尤其是 Zn2+、Cu2+、Mn2+,可能是這些重金屬離子占據了酶的活性中心或與酶分子的巰基、胺基和羧基等基團結合導致酶活性降低甚至失活即有抑制作用[16],甚至直接毒害生物膜中的微生物,使其喪失生命力[5]。通常來說,酸堿度是影響藻類或細菌磷酸酶活性的重要因子,我們的研究發現自然生物膜的 ALPase 具有比 ACPase 更高的酶活性,這可能是由于自然生物膜屬于光營養性微生物聚集體,其光合作用導致微域環境呈堿性條件[26],并適應該堿性環境,酸性條件一定程度上抑制了微生物的生命活力。
通常來說,抑制劑對酶促反應的作用可分為[27]:①競爭性抑制:競爭性抑制是指當抑制物與底物的結構類似時,它們將競爭酶的同一可結合部位,阻礙了底物與酶相結合,導致酶催化反應速率降低。這種抑制使得m增大,而max不變。②非競爭性抑制:抑制劑與酶的非活性部位相結合,形成抑制物-酶的絡合物后會進一步再與底物結合;或是酶與底物結合成底物酶絡合物后,其中有部分再與抑制物結合。雖然底物、抑制物和酶的結合無競爭性,但兩者與酶結合所形成的中間絡合物不能直接生成產物,導致了酶催化反應速率的降低,這種抑制使得max變小,但m不變。③反競爭性抑制:抑制劑不直接與游離酶相結合,而僅與酶-底物復合物結合形成底物-酶-抑制劑復合物,從而影響酶促反應的現象。這種抑制作用使得max、m都變小,但max/m比值不變。Mg2+同時降低了ACPase和ALPase的m值,說明其有助于提高生物膜磷酸酶與底物結合的能力。Cu2+對ACPase的非競爭性抑制作用說明Cu2+可能與ACPase的非活性部分結合,或與酶促反應中的中間產物絡合物結合并阻止產物生成,使酶的催化活性降低。而Zn2+對于磷酸酶以及Cu2+對于ALPase的抑制作用并不符合競爭抑制性、非競爭抑制性或反競爭性抑制性作用,可能是直接對生物膜產生了毒害作用。通常來說,在土壤、植物、活性污泥或動物糞便中的酸性或堿性磷酸酶動力學常數m值是相似的,在0.2 ~ 0.8 mmol/L 之間,而我們的研究發現自然生物膜的磷酸酶的m值在0.1左右,即使在金屬離子的影響下,m值仍小于0.2,說明自然生物膜的磷酸酶對底物有更強的親和力,而且這種親和力受金屬離子的影響相對較小。而酶動力學常數max值通常隨著所研究的材料的生物量濃度或不同物質的富集程度而顯著變化,例如,各種磷酸酶的max值在植物材料中比在土壤、動物糞便和污水污泥中大得多。對max值產生顯著影響的抑制劑濃度也會有很大差異。例如,Cu2+對活性污泥中堿性磷酸酶的抑制模型與土壤中游離和固定化酸性磷酸酶的抑制模型相似,但Cu2+對活性污泥中堿性磷酸酶的抑制濃度(0.1 ~ 2.5 mmol/L)明顯高于土壤的(0.1 ~ 0.4 mmol/L) 。這可能是由于活性污泥中生物量或磷酸酶的濃度較高[22]。而我們研究的稻田自然生物膜的生物組成可能與活性污泥中微生物群落的復雜性更加相似。
4 結論
1)不同金屬離子對稻田自然生物膜酸性或堿性磷酸酶具有不同的作用,有可能抑制、激活或無明顯作用,這種作用因金屬離子的種類、濃度和磷酸酶類型而異。
2)金屬離子影響自然生物膜中磷酸酶活性,主要通過改變酸性或堿性磷酸酶與底物的親和力和催化效率,以及影響磷酸酶基因的表達實現的。
3)自然生物膜中磷酸酶對底物有很強的親和力,金屬離子對這種親和力的影響較小。而且由于自然生物膜本身是一個復雜的微生態系統,因此抵御金屬離子毒性的作用也明顯強于單一微生物種群。
[1] Larned S T. A prospectus for periphyton: Recent and future ecological research[J]. Journal of the North American Benthological Society, 2010, 29(1): 182–206.
[2] Wu Y, Yang J L, Tang J, et al. The remediation of extremely acidic and moderate pH soil leachates containing Cu (II) and Cd (II) by native periphytic biofilm[J]. Journal of Cleaner Production, 2017, 162: 846–855.
[3] Li J Y, Deng K Y, Hesterberg D, et al. Mechanisms of enhanced inorganic phosphorus accumulation by periphyton in paddy fields as affected by calcium and ferrous ions[J]. Science of The Total Environment, 2017, 609: 466–475.
[4] Pohlon E, Marxsen J, Küsel K. Pioneering bacterial and algal communities and potential extracellular enzyme activities of stream biofilms[J]. FEMS Microbiology Ecology, 2010, 71(3): 364–373.
[5] 張妍, 崔驍勇, 羅維. 重金屬污染對土壤微生物生態功能的影響[J]. 生態毒理學報, 2010, 5(3): 305–313.
[6] Headley J V, Gandrass J, Kuballa J, et al. Rates of sorption and partitioning of contaminants in river biofilm[J]. Environmental Science & Technology, 1998, 32(24): 3968–3973.
[7] Dong D M, Derry L A, Lion L W. Pb scavenging from a freshwater lake by Mn oxides in heterogeneous surface coating materials[J]. Water Research, 2003, 37(7): 1662–1666.
[8] Dong D M, Hua X Y, Li Y, et al. Cd adsorption properties of components in different freshwater surface coatings: the important role of ferromanganese oxides[J]. Environmental Science & Technology, 2003, 37(18): 4106–4112.
[9] Dong D M, Li Y, Zhang J J, et al. Comparison of the adsorption of lead, cadmium, copper, zinc and Barium to freshwater surface coatings[J]. Chemosphere, 2003, 51(5): 369–373.
[10] Serra A, Corcoll N, Guasch H. Copper accumulation and toxicity in fluvial periphyton: the influence of exposure history[J]. Chemosphere, 2009, 74(5): 633-641.
[11] Fortin C, Denison F H, Garnier-Laplace J. Metal– phytoplankton interactions: modeling the effect of competing ions (H+, Ca2+, and Mg2+) on uranium uptake[J]. Environmental Toxicology and Chemistry, 2007, 26(2): 242.
[12] Ellwood N T W, Pippo F D, Albertano P. Phosphatase activities of cultured phototrophic biofilms[J]. Water Research, 2012, 46(2): 378–386.
[13] Applebury M L, Coleman J E. Escherichia coli alkaline phosphatase[J]. Journal of Biological Chemistry, 1969, 244(2): 308.
[14] Luo M, Guo Y C, Deng J Y, et al. Characterization of a monomeric heat-labile classical alkaline phosphatase from Anabaena sp. PCC7120[J]. Biochemistry (Moscow), 2010, 75(5): 655–664.
[15] Guzzon A, Bohn A, Diociaiuti M, et al. Cultured phototrophic biofilms for phosphorus removal in wastewater treatment[J]. Water Research, 2008, 42(16): 4357–4367.
[16] 高秀麗, 邢維芹, 冉永亮, 等. 重金屬積累對土壤酶活性的影響[J]. 生態毒理學報, 2012, 7(3): 331–336.
[17] Lu H Y, Yang L Z, Shabbir S, et al. The adsorption process during inorganic phosphorus removal by cultured periphyton[J]. Environmental Science and Pollution Research, 2014, 21(14): 8782–8791.
[18] 廖金花, 陳清西. 金屬離子對鮑魚堿性磷酸酶活力的影響[J]. 廈門大學學報(自然科學版), 2004, 43(S1): 12–15.
[19] Wu Y H, Li T L, Yang L Z. Mechanisms of removing pollutants from aqueous solutions by microorganisms and their aggregates: A review[J]. Bioresource Technology, 2012, 107(107): 10–18.
[20] Yang J L, Tang C L, Wang F W, et al. Co-contamination of Cu and Cd in paddy fields: Using periphyton to entrap heavy metals[J]. Journal of Hazardous Materials, 2016, 304: 150–158.
[21] 陸珊, 胡源, 蘇波, 等. 幾種金屬離子對麥芽酸性磷酸酶活性及構象的影響[J]. 化學研究與應用, 2007, 19(4): 386–389.
[22] Xie C S, Lu R J, Huang Y, et al. Effects of ions and phosphates on alkaline phosphatase activity in aerobic activated sludge system[J]. Bioresource Technology, 2010, 101(10): 3394–3399.
[23] Chen Q X, Zheng W Z, Lin J Y, et al. Effect of metal ions on the activity of green crab (Scylla serrata) alkaline phosphatase[J]. The International Journal of Biochemistry & Cell Biology, 2000, 32(8): 879–885.
[24] Wu J R, Shien J, Shieh H K, et al. Cloning of the gene and characterization of the enzymatic properties of the monomeric alkaline phosphatase (PhoX) from Pasteurella multocida strain X-73[J]. Fems Microbiology Letters, 2007, 267(1): 113–120.
[25] 趙娜, 朱琳, 馮鳴鳳. 不同pH條件下Cr6+對3種藻的毒性效應[J]. 生態毒理學報, 2010, 5(5): 657–665.
[26] Lu H Y, Wan J J, Li J Y, et al. Periphytic biofilm: a buffer for phosphorus precipitation and release between sediments and water[J]. Chemosphere, 2016, 144: 2058–2064.
[27] 呂迎春, 孫志偉, 宋佳偉,等. 磷和金屬對榮成天鵝湖水體堿性磷酸酶活性和動力學性質的影響研究[J]. 環境科學學報, 2018, 38(10): 4083–4089.
Effects of Metal Ions on Phosphatase Activity of Periphytic Biofilm in Paddy Fields
CAI Shujie1,2, DENG Kaiying1, LI Jiuyu1*, XU Renkou1
(1 State Key Laboratory of Soil and Sustainable Agriculture, Institute of Soil Science, Chinese Academy of Sciences, Nanjing 210008, China; 2 University of Chinese Academy of Sciences, Beijing 100049, China)
The effects of metal ions on acidic and alkaline phosphatase (ACPase and ALPase) activities and kinetic parameters of periphytic biofilm in paddy fields were studied. The results suggested that both K+and Na+had no significant effect on ACPase and ALPase activities in the concentration range of 0-1.0 mmol/L while both Ca2+and Mg2+had activating effects on ACPase and ALPase activities, which increased phosphatase activity by 14.1% and 46.7%, respectively. The metal ions of Zn2+, Cu2+, Mn2+, Al3+and Ag+all had inhibitory effect on ACPase and ALPase activities. While Co2+could promote ACPase and ALPase activities at low concentration, but inhibit them at high concentration. And Cr6+promoted ACPase activity at 0.1 mmol/L, while inhibited ACPase activity in the concentration range of 0.25-1.0 mmol/L. The addition of Mg2+enhanced the affinity and catalytic efficiency of ACPase with substrates. The inhibitory effect of Cu2+on ACPase and ALPase activities was mainly considered as non-competitive inhibition. Although Zn2+increased the affinity of ACPase with substrate, it reduced the affinity and catalytic efficiency of ALPase with substrate. The effect of metal ions on enzyme activity varied with the type and concentration of metal ions, and the type of phosphatase. It is mainly achieved by binding with the site or substrate of enzyme activity, changing the affinity between enzyme activity and substrate, and influencing the expression of enzyme genes. The results can provide a theoretical basis for assessing the role of periphytic biofilm in the biogeochemical cycle of phosphorus in paddy fields, and the possible risk of heavy metal contamination in paddy fields.
Periphytic biofilm; Phosphatase activity; Metal ions; Enzyme kinetic constant;Paddy fields
S511
A
10.13758/j.cnki.tr.2020.03.015
蔡述杰, 鄧開英, 李九玉, 等. 不同金屬離子對稻田自然生物膜磷酸酶活性的影響. 土壤, 2020, 52(3): 525–531.
國家重點基礎研究發展計劃(973計劃)青年科學家專題(2015CB158220),國家自然科學基金項目(41877102)和中國科學院創新交叉團隊和創新促進會項目(2014272)資助。
(jyli@issas.ac.cn)
蔡述杰(1992—),男,福建寧德人,碩士研究生,主要從事稻田土壤界面化學研究。E-mail: sjcai@issas.ac.cn
猜你喜歡
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
當代陜西(2021年2期)2021-03-29 07:41:24
家庭影院技術(2020年10期)2020-12-14 07:54:18
媽媽寶寶(2017年3期)2017-02-21 01:22:28
中國塑料(2016年3期)2016-06-15 20:30:00
通信電源技術(2016年3期)2016-03-26 07:13:38
知識經濟·中國直銷(2016年3期)2016-02-27 16:15:49
現代檢驗醫學雜志(2014年6期)2014-02-02 03:02:04
閱讀與作文(小學低年級版)(2011年3期)2011-01-01 00:00:00
