低溫脅迫對不同雜交水稻組合苗期耐冷性及生理生化指標(biāo)的影響
2020-07-20 03:26:38沈航陳嬌黎佰楊
江蘇農(nóng)業(yè)科學(xué) 2020年12期
關(guān)鍵詞:水稻
沈航 陳嬌 黎佰楊


摘要:為明確雜交水稻萌發(fā)率、苗期耐冷性與耐冷性指標(biāo)的關(guān)系,以重慶師范大學(xué)10個自育雜交水稻組合和糯稻89-1為材料,測定種子的生理指標(biāo)、低溫萌發(fā)率及苗期低溫處理恢復(fù)生長后的死苗率。不同雜交水稻組合的可溶性糖含量、維生素C含量、過氧化氫酶活性差異明顯,種子自身所含這4種物質(zhì)與水稻低溫下的萌發(fā)率可能具有一定關(guān)系;種子萌發(fā)對溫度變化較敏感,每個組合中溫度為6 ℃處理?xiàng)l件下種子萌發(fā)率均高于11 ℃,初步推斷臨界點(diǎn)以下低溫層積對水稻種子的萌發(fā)有積極作用;試驗(yàn)結(jié)果表明,不同低溫條件處理下水稻苗期死亡率有明顯差異,溫度越低,水稻幼苗受害越大,復(fù)溫后死亡率越高;聚類分析將雜交組合分為A、B、C 3類,A組、B組生理指標(biāo)大部分都高于C組,低溫萌發(fā)不同雜交水稻組合內(nèi)變異系數(shù)較大。總的來說,水稻種子耐冷指標(biāo)對萌發(fā)率可能具有一定作用,在苗期,水稻對低溫較敏感,隨著時間和溫度增長受影響越大,根據(jù)耐冷指標(biāo)、萌發(fā)率、死苗率對不同水稻雜交組合進(jìn)行分類,研究結(jié)果可以為選育耐冷性水稻品種提供理論依據(jù)。
關(guān)鍵詞:糯稻89-1;雜交水稻;幼苗;耐冷;低溫脅迫、萌發(fā)率
水稻是我國最重要的糧食作物之一,與國家糧食安全密切相關(guān)[1]。水稻種植常受氣溫影響,一般每4~5年就發(fā)生1次較大規(guī)模的低溫冷害[2]。據(jù)統(tǒng)計,全世界有24個國家、1 500萬hm2以上的稻作面積時常受到低溫威脅,使水稻嚴(yán)重減產(chǎn),低溫冷害儼然已成為全球性自然災(zāi)害[3-6]。日本最早于20世紀(jì)30年代開始水稻的耐冷性研究[7],至今已有70多年的歷史,我國從20世紀(jì)70—80年代開始水稻耐冷性研究[8],如今,國內(nèi)外對于水稻的耐冷性研究已取得較好的進(jìn)展,在耐冷性基因定位克隆、耐冷生理生化和分子機(jī)制等方面都有很大的進(jìn)展。
研究水稻耐冷性的目的在于培育出耐冷性強(qiáng)的水稻品種,以減輕低溫冷害對水稻的傷害。水稻耐冷性研究雖然取得了很多的成果,但選育出的耐冷性強(qiáng)的水稻品種卻較少[9]。糯稻89-1是耐冷性極強(qiáng)的材料,其芽期、幼苗期的耐冷性均達(dá)到一級標(biāo)準(zhǔn)[10]。糯稻89-1能通過腋芽休眠越過冬季,次年萌發(fā)再生,越冬再生季產(chǎn)量達(dá)6 291 kg/hm2,與正季相當(dāng)[11],糯稻89-1是用于研究水稻耐低溫的珍稀材料。本研究以糯稻89-1為親本與多個品種(系)進(jìn)行雜交,對不同雜交組合后代的低溫萌發(fā)以及苗期的耐冷性進(jìn)行分析,以期為耐冷性水稻材料的篩選提供理論依據(jù)。
1 材料與方法
1.1 試驗(yàn)材料
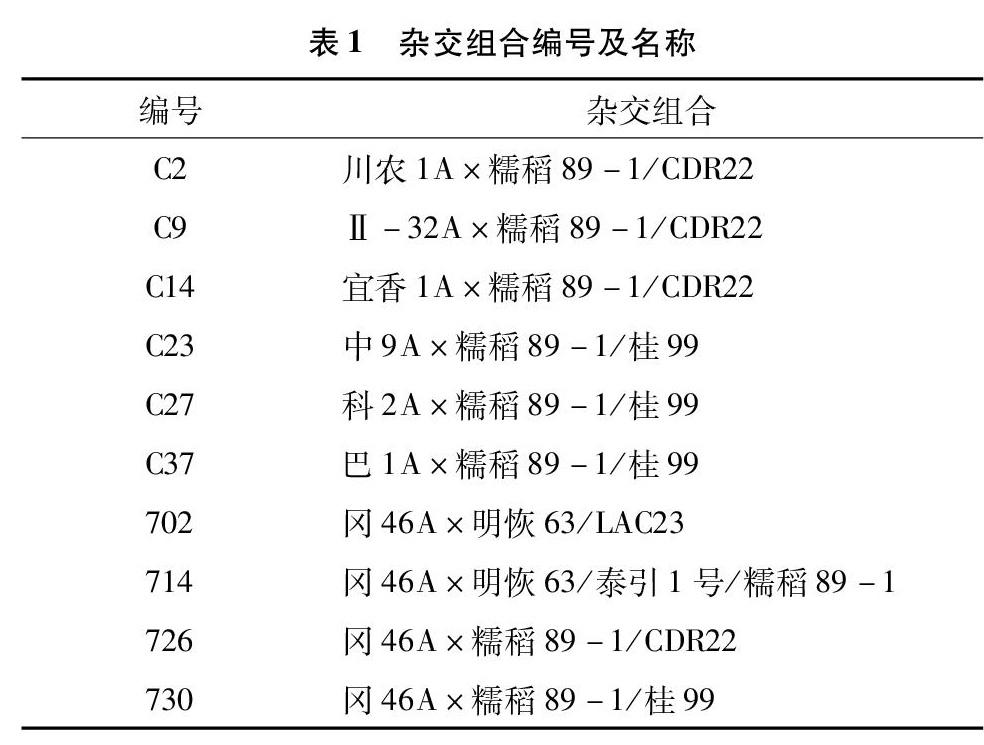
以重慶師范大學(xué)自育的10個雜交水稻組合、糯稻89-1為材料(表1)。試驗(yàn)在重慶師范大學(xué)水稻育種基地進(jìn)行,于2017年3月播種,8月收集種子,保存3個月后進(jìn)行生理指標(biāo)的測定和不同溫度下的萌發(fā)試驗(yàn)。所配組合中糯稻89-1為一級耐冷性品種。
1.2 試驗(yàn)方法
1.2.1 種子生理指標(biāo)的測定 精選飽滿的籽粒進(jìn)行測定。各組合分別做3次重復(fù),取平均值。可溶性糖(SS)含量的測定采用蒽酮-硫酸比色法[12];過氧化氫酶(CAT)活性的測定采用高錳酸鉀滴定法[13];丙二醛(MDA)含量的測定采用硫代巴比妥酸(TBA)顯色法[12];維生素C含量的測定采用鉬藍(lán)比色法[14]。
1.2.2 種子萌發(fā)率 選取飽滿的籽粒經(jīng)常規(guī)方法消毒后,置于培養(yǎng)皿中。先用蒸餾水侵泡10 min后,去除半飽滿籽粒。用3%的次氯酸侵泡30 min后,用去離子水沖洗干凈并均勻平鋪于無菌培養(yǎng)皿中,放置于不同溫度下培養(yǎng),其間每天定時換水并保持培養(yǎng)皿濕潤。設(shè)置的溫度分別為6、11、16、27 ℃,12 d 后統(tǒng)計萌發(fā)率,各組合3次重復(fù),取平均值。12 d 后,將6、11 ℃條件下未萌發(fā)的水稻種子轉(zhuǎn)入27 ℃條件下繼續(xù)萌發(fā),12 d后統(tǒng)計萌發(fā)率。
1.2.3 苗期耐冷性鑒定 取不同雜交水稻組合種子100粒,滅菌后置于27 ℃條件下,在0.8%瓊脂糖培養(yǎng)基上培養(yǎng),待幼苗長至3葉1心時,留50株長勢相近的幼苗移至溫度為6、11、16 ℃條件下培養(yǎng)12 d后測量苗高、根長,統(tǒng)計死亡率,12 d后,將溫度為6、11 ℃條件下的水稻幼苗移至27 ℃條件下恢復(fù)生長7 d,統(tǒng)計死苗率。3次重復(fù),取平均值。
1.3 數(shù)據(jù)處理
采用Microsoft Excel 2010處理試驗(yàn)數(shù)據(jù)和繪制圖表,用SPSS 22.0進(jìn)行平均值、單因素方差、相關(guān)性及聚類分析,聚類選用Ward法分析。
2 結(jié)果與分析
2.1 不同雜交水稻組合生理指標(biāo)比較
10個供試雜交水稻組合不同生理生化指標(biāo)差異明顯,MDA含量的差異性最大,CAT活性差異性最小。其中,維生素C含量變化范圍為6.325~10.983 mg/100 g(圖1-A);可溶性糖含量變化范圍為20.73%~36.52%,平均值為26.84%(圖1-B);過氧化氫酶活性變化范圍為58.9~92.8 H2O2 mg/(g·min)(圖1-C);丙二醛含量變化范圍為0.016 4~0.250 4 μmol/g(圖1-D)。726雜交水稻組合各項(xiàng)指標(biāo)均低于平均值,MDA含量比平均值降低726.59%。
2.2 不同溫度條件下不同雜交水稻組合種子萌發(fā)率比較
為明確種子耐冷性生理指標(biāo)對萌發(fā)率的影響,對10個不同雜交水稻組合進(jìn)行不同溫度下種子萌發(fā)試驗(yàn)。恒溫萌發(fā)條件下,溫度為27 ℃時萌發(fā)率為96.67%,溫度為16 ℃時種子雖能萌發(fā),但萌發(fā)率均低于23%(圖2)。變溫萌發(fā)中,溫度為6、11 ℃的條件下培養(yǎng)種子12 d后萌發(fā)率仍為0,繼轉(zhuǎn)入溫度為27 ℃萌發(fā),培養(yǎng)1 d后開始露白;6 ℃轉(zhuǎn)至27 ℃的平均萌發(fā)率為91.29%,11 ℃轉(zhuǎn)至27 ℃的平均萌發(fā)率為84.48%。每個組合中6 ℃處理過的種子萌發(fā)率均高于11 ℃。但6 ℃轉(zhuǎn)至27 ℃萌發(fā)率低于27 ℃條件下的萌發(fā)率,可能是因?yàn)榈蜏叵绿幚淼臅r間過長。不同低溫處理對轉(zhuǎn)溫后不同雜交水稻組合種子萌發(fā)率的影響有所差異,其中對C23、702、730的雜交水稻組合影響最大,但對726雜交水稻組合影響較小。且726在不同溫度條件下的萌發(fā)率均為最低,與耐冷性生理指標(biāo)相對應(yīng)(圖3)。
糯稻89-1/CDR22與川農(nóng)1A、Ⅱ-32A、宜香1A雜交組合歸為A組,糯稻89-1/桂99與中9A、科2A、巴1A 3個不育系所配同父異母組合歸為B組。4個不同恢復(fù)系與岡46A不育系所配同母異父雜交組合歸為C組。A組、B組維生素C含量、可溶性糖含量、過氧化氫酶活性平均值均高于C組,溫度為27 ℃條件下種子萌發(fā)率是低溫(16℃)條件下的5倍以上,低溫萌發(fā)不同組合內(nèi)的變異系數(shù)較大。變溫下,溫度為6 ℃處理的種子萌發(fā)率均大于 11 ℃,表明不同的低溫處理對種子萌發(fā)有明顯的影響(表2)。
2.3 低溫脅迫對水稻幼苗及恢復(fù)生長的影響
從圖4可以看出,低溫處理下,水稻幼苗普遍生長緩慢或接近停滯,并出現(xiàn)干枯腐爛的現(xiàn)象,且不同低溫脅迫對幼苗的影響差異明顯。溫度為16 ℃條件下,不同水稻雜交組合幼苗均能夠繼續(xù)生長,但生長速度較27 ℃慢,培養(yǎng)3 d后,C9、C2雜交組合葉尖開始出現(xiàn)變黃跡象,繼續(xù)培養(yǎng) 9 d 后發(fā)現(xiàn)不同水稻雜交組合葉尖均有不同程度的變黃,730、702、C37、C9雜交組合葉尖變黃程度嚴(yán)重,由葉尖變黃轉(zhuǎn)為整葉變黃,最大變黃長度達(dá)3.3 cm,而糯 89-1 及726、714、C14雜交組合生長狀態(tài)良好,葉尖變黃株數(shù)少。溫度為6、11 ℃低溫處理下,不同組合后代死苗率差異較大,糯89-1和714在2個溫度條件下死苗率均為0,溫度為11 ℃條件下幼苗死亡率普遍高于6 ℃ 條件下。27 ℃恢復(fù)生長7 d后,濕度為6 ℃處理的幼苗死亡率明顯高于11 ℃處理的幼苗,且除糯89-1外的幼苗經(jīng)過恢復(fù)生長期幾乎全部死亡,C23、714雜交水稻組合由0上升至100%。C9、726、702雜交水稻組合幼苗死亡率相對較低,分別為84.00%、84.16%、77.86%。濕度為11 ℃處理下的幼苗死亡率雖然較恢復(fù)生長前有不同程度的增加,但大多數(shù)組合能恢復(fù)正常生長,其中C2、714雜交水稻組合幼苗死亡率較低,分別為4.05%、6.25%。
3 討論與結(jié)論
植物種子播種后能否正常萌發(fā)是不同的內(nèi)源性和環(huán)境因素相互作用,共同調(diào)節(jié)種子休眠與萌發(fā)之間的平衡的結(jié)果[9,15-16]。10個水稻雜交組合可溶性糖含量、維生素C含量、過氧化氫酶活性差異明顯,這與不同水稻雜交組合遺傳物質(zhì)的差異有關(guān)。3個指標(biāo)與種子低溫下萌發(fā)率以及低溫處理后恢復(fù)正常溫度下的萌發(fā)率具有相關(guān)性,可能是種子可溶性糖含量、維生素C含量、過氧化氫酶活性與低溫下種子的萌發(fā)率有關(guān),本試驗(yàn)的結(jié)果與張啟雷等的研究結(jié)論[17]相同。
種子萌發(fā)對溫度的變化較敏感,低溫是種子萌發(fā)的脅迫因素之一[18]。低溫環(huán)境下,水稻種子萌發(fā)率會降低,其降低程度與種子耐低溫能力有關(guān)。本研究中6、11 ℃低溫條件下,不同水稻雜交組合萌發(fā)率均為0,說明該溫度抑制了種子的萌發(fā)。溫度為16 ℃時不同水稻雜交組合種子均有萌發(fā),但萌發(fā)率普遍較低,糯89-1在16 ℃萌發(fā)12 d萌發(fā)率仍然能達(dá)到78%,說明低溫耐性強(qiáng),這與趙正武等的研究結(jié)果[18]具有一致性。本研究中,不同水稻雜交組合種子在不同溫度處理后,恢復(fù)正常溫度萌發(fā)時其萌發(fā)率均具有明顯差異,初步推斷種子萌發(fā)階段,低溫脅迫的強(qiáng)度對于脅迫后水稻種子活力的恢復(fù)有重要的影響。不同水稻雜交組合的種子在經(jīng)過6、11 ℃ 低溫處理后,其萌發(fā)率仍保持在74.07%~96.39%之間,大部分甚至高于優(yōu)良親本糯 89-1,其中涉及的潛在育種可利用潛力以及對深入研究種子經(jīng)歷低溫后萌發(fā)的生理機(jī)制和遺傳效應(yīng)都具有重要的利用價值。當(dāng)溫度低于12 ℃時,遭遇中等倒春寒(日平均氣溫≤12.0 ℃ 4~6 d或日平均氣溫≤14.0 ℃ 7~10 d)甚至是強(qiáng)倒春寒(日平均氣溫≤12.0 ℃≥7 d或日平均氣溫≤14.0 ℃≥11 d)時,種子不能萌發(fā),度過倒春寒后,氣溫回暖,種子活力基本恢復(fù),方可繼續(xù)萌發(fā);除種子萌發(fā)受低溫影響外,低溫還會抑制幼苗的生長,使幼苗出現(xiàn)葉尖變黃、生長緩慢或存活率單向減少等現(xiàn)象[19-20]。低溫引起植株葉片形態(tài)的變化是植物對冷害最直觀的反映,主要包括葉片卷曲、萎縮、葉色褐化,甚至干枯死亡[21]。本研究中,溫度在6 ℃條件下,幼苗幾乎不生長,并且葉片失綠變黃干枯。轉(zhuǎn)入27 ℃恢復(fù)生長時,在溫度為6 ℃條件下脅迫12 d的幼苗幾乎全部生長霉菌死亡,溫度為11 ℃條件下大部分組合能度過恢復(fù)生長期,正常生長,低溫脅迫嚴(yán)重影響幼苗生長,脅迫期間溫度越低,對幼苗造成的傷害越大,在恢復(fù)生長期死亡率越高。此外,水稻雜交組合702盡管在冷害時出現(xiàn)整株枯黃的現(xiàn)象但在溫度為27 ℃復(fù)育后死亡率最低,這可能與越冬再生的性狀有關(guān)。
聚類分析的結(jié)果表明,10個水稻雜交組合的低溫耐受性與糯89-1差異較大。水稻雜交組合C2、C9、C14以及C23、C27、C37為同父異母組合,714、726、730為同母異父組合,這9個組合并未按同父異母或是同母異父的組配方式聚為一類,各組合低溫耐受性差異不大,說明苗期的耐冷性不存在細(xì)胞質(zhì)遺傳,而是受細(xì)胞核基因控制的,劉建豐等研究發(fā)現(xiàn),雜交稻F1苗期耐冷性傾向于母本,受父本影響較小[22-23],出現(xiàn)不同結(jié)果可能是由于試驗(yàn)所利用的低溫處理?xiàng)l件和耐冷性評價標(biāo)準(zhǔn)以及試驗(yàn)材料耐冷性程度不同。另外,除了702,其他9個組合均有糯89-1血緣,這9個雜交組合的苗期耐冷性與702相差不大,表明糯89-1耐冷性的遺傳力較低,這與金潤洲對耐冷性遺傳研究的結(jié)果[24]相反,可能是所用材料的不同導(dǎo)致的。
水稻種子在低溫脅迫下的萌發(fā)率很大程度上由自身生理物質(zhì)含量的高低決定,而丙二醛含量、可溶性糖含量、維生素C含量、過氧化氫酶活性高低對水稻種子在低溫脅迫下種子的萌發(fā)情況可能起到關(guān)鍵作用。水稻在苗期遇到冷害會影響秧苗的發(fā)育,不同雜交組合受到低溫影響的表現(xiàn)基本相同,這與潘孝武等的試驗(yàn)結(jié)果[25]基本一致。本研究主要致力于挖掘以糯稻89-1為主要親本的不同雜交水稻組合的耐冷生理指標(biāo)與萌發(fā)率和苗期耐冷能力的關(guān)系,在涉及到糯稻89-1耐冷性狀遺傳方面還須進(jìn)一步深入研究。
參考文獻(xiàn):
[1]鄧興旺,王海洋,唐曉艷,等. 雜交水稻育種將迎來新時代[J]. 中國科學(xué)(生命科學(xué)),2013,43(10):864-868.
[2]高益波,景元書,劉明璐,等. 抽穗揚(yáng)花期低溫強(qiáng)度對水稻生長發(fā)育的影響與模擬[J]. 江蘇農(nóng)業(yè)科學(xué),2018,46(5):53-57.
[3]Sthapit B R,Witcombe J R. Inheritance of tolerance to chilling stress in rice during germination and plumule greening[J]. Crop Science,1998,38(3):660-665.
[4]王士強(qiáng),趙海紅,趙黎明,等. 水稻冷害生理功能變化與調(diào)控研究進(jìn)展[J]. 中國農(nóng)學(xué)通報,2017,33(36):1-6.
[5]王主玉,申雙和. 水稻低溫冷害研究進(jìn)展[J]. 安徽農(nóng)業(yè)科學(xué),2010,38(22):11971-11973.
[6]崔 迪,楊春剛,湯翠鳳,等. 低溫脅迫下粳稻選育品種耐冷性狀的鑒定評價[J]. 植物遺傳資源學(xué)報,2012,13(5):739-747.
[7]Nishiyama I. Proceedings of the symposium on climate and rice[M]. Manila:IRRI,1976:159-185.
[8]韓龍植,張三元. 水稻耐冷性鑒定評價方法[J]. 植物遺傳資源學(xué)報,2004,5(1):75-80.
[9]王尚明,賀浩華,肖葉青,等. 水稻東野1號苗期耐冷性遺傳分析[J]. 湖北農(nóng)業(yè)科學(xué),2008,47(1):1-4.
[10]王春萍,雷開榮,李正國,等. 低溫脅迫對水稻幼苗不同葉齡葉片葉綠素?zé)晒馓匦缘挠绊慬J]. 植物資源與環(huán)境學(xué)報,2012,21(3):38-43.
[11]趙正武,李仕貴,雷樹凡. 糯稻89-1越冬性遺傳研究[J]. 中國農(nóng)業(yè)科學(xué),2006,39(12):2399-2405.
[12]趙世杰,史國安,董新純. 植物生理學(xué)實(shí)驗(yàn)指導(dǎo)[M]. 北京:中國農(nóng)業(yè)科學(xué)技術(shù)出版社,1998:136-139.
[13]湯紹虎,羅 充. 植物生理學(xué)實(shí)驗(yàn)教程[M]. 重慶:西南師范大學(xué)出版社,2012.
[14]孫 群,胡景江. 植物生理學(xué)研究技術(shù)[M]. 楊凌:西北農(nóng)林科技大學(xué)出版社,2006.
[15]簡水溶,萬 勇,羅向東,等. 東鄉(xiāng)野生稻苗期耐冷性的遺傳分析[J]. 植物學(xué)報,2011,46(1):21-27.
[16]Shu K,Meng Y J,Shuai H W,et al. Dormancy and germination:how does the crop seed decide?[J]. Plant Biology,2015,17(6):1104-1112.
[17]張啟雷,劉 強(qiáng),高 輝,等. 內(nèi)源抗壞血酸對水稻種子萌發(fā)及幼苗生長的影響[J]. 熱帶亞熱帶植物學(xué)報,2016,24(3):273-279.
[18]趙正武,王述民,李世平. 珍稀稻種資源越冬糯稻89-1研究初報[J]. 雜交水稻,2000,15(3):3.
[19]李太貴,王 磊. 低溫下水稻幼苗形態(tài)生理應(yīng)變研究[J]. 生態(tài)學(xué)雜志,1996,15(6):61-63,50.
[20]Mahajan S,Tuteja N. Cold,salinity and drought stresses:an overview[J]. Archives of Biochemistry and Biophysics,2005,444(2):139-158.
[21]許 英,陳建華,朱愛國,等. 低溫脅迫下植物響應(yīng)機(jī)理的研究進(jìn)展[J]. 中國麻業(yè)科學(xué),2015,37(1):40-49.
[22]劉建豐,陳立云. 水稻耐冷性研究現(xiàn)狀與展望[J]. 作物研究,1996,10(2):42- 44.
[23]李 平,劉鴻先,王以柔,等. 低溫對雜交水稻及其親本三系始穗期旗葉光合作用的影響[J]. 植物學(xué)報,1990,32(6):456-464.
[24]金潤洲. 日本關(guān)于水稻耐冷性鑒定及其遺傳的研究[J]. 水稻文摘,1990,9(3):1-5.
[25]潘孝武,李小湘,黎用朝,等. 湖南早秈稻品種芽期和苗期耐冷性初步研究[J]. 雜交水稻,2015,30(5):59-64.鄧惠清,唐燦明,劉正鑾,等. 陸地棉空間誘變SP1~SP3主要性狀變異分析[J]. 江蘇農(nóng)業(yè)科學(xué),2020,48(12):64-67,74.
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學(xué)版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農(nóng)業(yè)(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農(nóng)業(yè)(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農(nóng)業(yè)(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
