“赤霞珠”葡萄自然發酵過程中高產β-葡萄糖苷酶酵母菌株篩選分析
2020-07-20 11:37:58夏俊芳古麗娜孜李璟瑜西熱娜依
食品工業科技 2020年13期
關鍵詞:酵母菌
夏俊芳,古麗娜孜,李璟瑜,禹 凱,西熱娜依,武 運
(新疆農業大學食品科學與藥學學院,新疆烏魯木齊 830052)
葡萄酒的香氣來自于葡萄漿果以及在加工、發酵和陳釀過程中產生的物質[1],包括無環萜烯醇、芳樟醇、香葉醇、橙花醇、香茅醇、松油醇以及單環萜烯醇等[2],它們以兩種形式存在:自由揮發物和非揮發性糖苷偶聯物[3]。自從首次報道葡萄果實中揮發性化合物的糖基化前體以來[4],人們普遍認為,糖苷在葡萄酒釀造過程中的水解作用是決定葡萄酒特征香氣形成的主要因素之一[5-6],大量研究表明糖苷結合物的釋放可以通過酸解或糖苷酶解的作用來實現,酸解在典型的葡萄酒釀造過程中是相當緩慢,而糖苷酶可以快速水解香氣前體并釋放揮發性物質[7],因此酶法被用作調節葡萄酒、提高葡萄酒香氣的重要方式。
β-葡萄糖苷酶(1,4-β葡萄糖苷酶,EC 3.2.1.21)是從糖苷前體釋放單萜物質的關鍵酶[8]。糖苷前體物被葡萄自身產生的酶和發酵中微生物產生的酶作用,促進葡萄酒香氣的形成[9],葡萄果實產生的β-葡萄糖苷酶主要位于漿果果皮的皮下壁,果實產生的β-葡萄糖苷酶在發酵條件(低pH、高葡萄糖或乙醇濃度)對葡萄酒的風味無明顯作用[10],而微生物(包括細菌、霉菌以及酵母等)產生的酶對葡萄酒的風味有明顯作用如:酵母菌在發酵中具有水解芳香前體物質的能力[11-12]。Perez等[13]使用七葉苷甘油瓊脂培養基從烏拉圭葡萄園分離出一些產β-葡萄糖苷酶優良釀酒酵母。Wang等[7]使用對硝基苯基β-D-吡喃葡萄糖苷(p-NPG)法分析兩株酵母內部和細胞外產β-葡萄糖苷酶情況。在釀酒條件下,酵母菌在各種糖苷上均有酶活性,因此不同地域環境存在各地區典型特征酵母[14-16],本地土著酵母與該地生態環境相適應,有利于釀造地域特色葡萄酒[17]。為了提高葡萄酒的特色香氣物質,探索釀酒酵母生物多樣性,本實驗從新疆天山北麓葡萄酒產區成熟赤霞珠(CabernetSauvignon)漿果自然發酵過程中篩選高產β-葡萄糖苷酶的優良酵母,利用WL培養基和賴氨酸培養基對其進行分類鑒定并構建18S rDNA發育樹,篩選地域特色高產β-葡萄糖苷酶的酵母菌株并探索脅迫因素下菌株產酶活力,為新疆葡萄酒的發展奠定基礎。
1 材料與方法
1.1 材料與儀器
“赤霞珠”(Cabernetsauvignon)葡萄 采摘自新疆天山北麓釀酒葡萄產區昌吉回族自治州大三工村甲、乙和丙葡萄種植園中,取樣成熟葡萄漿果裝入無菌袋,貯存于冰盒,24 h內送至實驗室用于酵母菌的分離篩選;商業酵母D254 煙臺帝伯仕自釀機有限公司;YPD固體培養基(葡萄糖20 g/L,酵母膏10 g/L,蛋白胨20 g/L,2%瓊脂)、初篩培養基(葡萄糖20 g/L,酵母膏10 g/L,蛋白胨20 g/L,2%瓊脂)滅菌后溫度降至60 ℃加入1 g/L p-NPG、糖類發酵基礎發酵管(蛋白胨10 g/L,NaCl 5 g/L,pH7.6置一倒置的杜氏小管)、碳源同化基礎培養基(0.5%(NH4)2SO4,0.1% KH2PO4,0.01%NaCl,0.01% CaCl2,0.05% MgSO4·7H2O,0.02%酵母膏,2%瓊脂)、氮源同化培養基(2%葡萄糖,0.1% KH2PO4,0.05% MgSO4·7H2O,0.02%酵母膏,2%瓊脂) 北京奧博星責任有限公司;p-NPG 美國Sigma公司;對硝基苯酚 上海阿拉丁生化科技股份有限公司;WL培養基、賴氨酸培養基 北京路橋生物有限公司;其他試劑 均為國產分析純。
FE20 PLUS pH計 上海梅特勒托利多儀器有限公司;DJ100-3型電子分析天平 上海恒平科學儀器有限公司;DY04-13-44-00 壓力蒸汽滅菌器 上海東亞壓力容器制造有限公司;722 可見分光光度計 上海欣茂儀器有限公司;THZ-98AB恒溫培養振蕩箱 上海一恒科學儀器有限公司;HSY2-SP電熱恒溫水浴鍋 北京市永光明醫療儀器廠;HR40-IIAI生物安全柜 青島海爾特種電器有限公司。
1.2 實驗方法
1.2.1 赤霞珠(Cabernetsauvignon)葡萄汁發酵過程中酵母菌株的分離 稱量約3.5 kg成熟度好的赤霞珠葡萄(糖度25 °Brix),手工破碎后,置于小型無菌玻璃發酵罐中,室溫下進行自然發酵,每個時期(發酵前期、發酵中期、發酵后期)取發酵液1 mL于9 mL無菌生理鹽水中,梯度稀釋10-4~10-7倍,取100 μL各稀釋液涂布在YPD培養基上,28 ℃下培養2~3 d,每個稀釋度做2個重復。將分離得到的酵母菌株在YPD平板上進行經多次劃線純化后轉接于YPD試管斜面,4 ℃保存。
1.2.2 產β-葡萄糖苷酶酵母菌株篩選 初篩:將1.2.1分離到的菌株以1%接種量接種到液體YPD培養基,培養28 ℃,24 h后以1%接種量接種至初篩培養基,再培養72 h后噴1 mol/L Na2CO3后觀察黃色光圈大小及顏色深淺能初步反映酶活性高低,將有深黃色光圈菌株保存下來。
復篩:將初篩得到的菌株活化后,再以1%接種量接種到YPD液體培養基中,28 ℃下培養 48 h后取發酵液10 mL于4 ℃8000 r/min離心10 min,收集上清液,測定上清液中的酶活力,從中篩選酶活較高菌株。
1.2.3β-葡萄糖苷酶活性的測定 參照侯曉瑞等[18]的方法,對硝基苯酚標準曲線回歸方程為:Y=20.607X+0.0119,決定系數R2=0.9996,吸光度與對硝基苯酚含量成線性關系。取0.1 mL粗酶液與0.2 mL 35 mmol/L p-NPG(pH5.0的檸檬酸-磷酸鹽緩沖液配制)混勻,50 ℃保溫10 min,加入2 mL 1 mol/L Na2CO3溶液終止反應并顯色,于400 nm下測定吸光值。以加熱滅活的酶液做空白對照。
酶活力計算公式如下:
式中:U為酶活力單位,U/mL;C為對硝基苯酚(p-NP)的濃度;V為反應體系的體積;N 為原酶液稀釋倍數;t 為反應時間;0.1為所取上清液或細胞液的體積。
以商業酵母D254為對照,對初篩中顯色較強的菌株,對比不同酵母菌株產β-葡萄糖苷酶活力。
1.2.4 產β-葡萄糖苷酶的菌株鑒定
1.2.4.1 WL、賴氨酸培養基分類鑒定 將1.2.3篩選的菌株挑單個菌落接種于10 mL YPD液體培養基,活化后劃線接種與WL培養基28 ℃培養72~96 h,根據菌落形態觀察對其進行初步分類。同時將1.2.2篩選的菌株挑單個菌落接種到5 mL無菌水中進行饑餓處理,120 h后接種到賴氨酸培養基,28 ℃培養120 h后觀察菌落生長情況,能夠在賴氨酸培養基上生長的酵母菌株屬于非釀酒酵母,無法生長的為釀酒酵母[18]。
1.2.4.2 生理生化鑒定 a.糖類發酵鑒定:將6種糖(葡萄糖、蔗糖、麥芽糖、半乳糖、乳糖、棉籽糖)分別加入糖類發酵基礎發酵管,分別接入0.1% 108CFU/mL酵母溶液,28 ℃ 200 r/min振蕩培養48 h,觀察杜氏小管內氣體產生情況,若小管頂部有氣體說明該菌能發酵此種糖[19]。b.碳源、氮源同化試驗:碳源同化,選用11種碳源作為同化試驗對象包括葡萄糖、蔗糖、半乳糖、麥芽糖、核糖、鼠李糖、乳糖、可溶性淀粉、甘露醇、蘋果酸、檸檬酸。分別加入上述11種不同的碳源,配制成不同碳源的液體培養基,備用;氮源同化,3種氮源作為同化試驗對象包括硝酸鉀、鹽酸尸胺和鹽酸乙胺,配制成不同氮源的液體培養基,備用;分別接入0.1% 108CFU/mL酵母溶液,28 ℃ 200 r/min振蕩培養48 h,如果試管出現混濁則為陽性“+”,否則計為陰性“-”[19]。
1.2.4.3 18S rDNA EF3/EF4區序列分析 利用18S rDNA的高度保守性,對篩選到的菌株進行分子生物學鑒定,這部分測序試驗由金唯智生物科技有限公司完成。18S rDNA PCR擴增引物EF3:5 ′-TCCTC TAAATGACCAAGTTTG-3′;EF4:5 ′-GGAAGGGRTG TATTTATTAG-3′[20],采用單菌落為模板進行PCR擴增。
PCR反應體系(25 μL)為:預混液12.5 μL,引物為EF3和EF4各0.5 μL,ddH2O 11 μL,模板DNA 0.5 μL。PCR擴增程序為95 ℃預變性 5 min;95 ℃變性 30 s,52 ℃退火 30 s,72 ℃延伸 1 min,32個循環,72 ℃ 延伸5 min。擴增后,取5 μL的擴增樣品在1%的瓊脂糖凝膠上80 V電壓下進行電泳檢測,將亮度好、純度高的特異性條帶(1500 bp 左右)的 PCR 產物用 TaKaRa Agarose Gel DNA Purification Kit Ver. 2.0(Code No. DV805A)試劑盒切膠回收,將對應目的片段置于-20 ℃下保存雙向引物測序,基因測序結果在NCBI通過BLAST進行序列比對分析,采用 MEGA 6.06軟件,構建菌株的系統發育樹。
1.2.5 不同脅迫因素對高產β-葡萄糖苷酶菌株產酶影響
1.2.5.1 乙醇對菌株產酶的影響 分裝5 mL YPD液體培養基于試管中,加入倒置的杜氏發酵管,滅菌后接種108CFU/mL G13、G21酵母菌懸液0.1 mL,分別加入無水乙醇,使其最終濃度分別為0%、10%、12%、14%、16%、18%和20%(v/v),28 ℃靜置培養48 h,分析乙醇對β-葡萄糖苷酶活力的影響。
1.2.5.2 葡萄糖對菌株產酶的影響 分裝5 mL YPD液體培養基于試管中并加入葡萄糖,使其最終濃度分別為2%、5%、10%、15%、20%和25%,加入倒置的杜氏發酵管,滅菌后接種108CFU/mL G13、G21酵母菌懸液0.1 mL,28 ℃靜置培養48 h,分析葡萄糖濃度對β-葡萄糖苷酶活力的影響。
1.2.5.3 pH對菌株產酶的影響 分裝5 mL YPD液體培養基于試管中并調整pH為2.0、3.0、4.0、5.0、6.0、7.0,加入倒置的杜氏發酵管,滅菌后接種108CFU/mL G13、G21酵母菌懸液0.1 mL,28 ℃靜置培養48 h,分析pH對β-葡萄糖苷酶活力的影響。
1.3 數據處理
所有數據均為3次平行實驗的平均值,數據統計分析采用Statistix 8.1(分析軟件,St Paul,MN)軟件包中Linear Models程序,差異顯著性采用Duncan檢驗進行多重比較,采用Excel 2016整理實驗數據、作圖。
2 結果與分析
2.1 葡萄酒相關酵母菌株分離
共分離到187株葡萄酒相關酵母,甲葡萄園分離到65株,乙葡萄園分離到70株,丙葡萄園分離到52株葡萄酒相關酵母菌。在發酵的整個過程都分離到相關釀酒酵母,主要集中在發酵的前期和中期,甲、乙、丙三個葡萄園在菌株分離的情況見表1。

表1 酵母菌株分離信息Table 1 The isolation information of yeast strains
2.2 產β-葡萄糖苷酶酵母菌株
187株出發菌株根據1.2.2方法將產酶能力初步分為4個等級:不產酶、弱產酶、中等產酶、較強產酶,如圖1。

圖1 產酶篩選培養基顯色Fig.1 The colour of selective medium for enzyme production注:A:不產酶;B:弱產酶;C:中等產酶;D:較強產酶。
2.3 產β-葡萄糖苷酶酵母酶活性特性分布
初篩得到的產β-葡萄糖苷酶活力的酵母菌株強度大小及分布情況見表2,甲園發酵前期較強,中等產酶的菌株占該園總產酶酵母總數的32%,乙園發酵前期較強,中等產酶菌株占該園產酶酵母總數的23%;丙園發酵前期較強,中等產酶菌株占該園產酶酵母總數的37.1%。總體分析,甲、乙、丙園中產β-葡萄糖苷酶酶活力菌株主要分布在發酵前期。
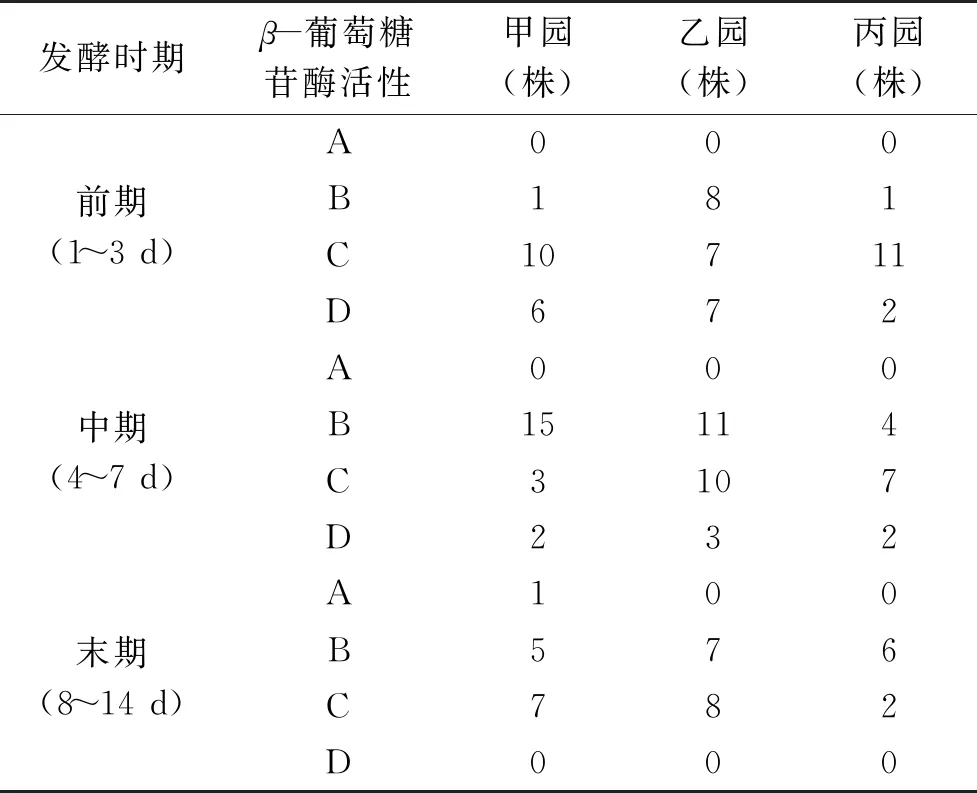
表2 產β-葡萄糖苷酶酵母在不同園區不同發酵期的分布Table 2 Distribution of yeast producing β-glucosidase at different fermentation stages in different parks
2.4 產β-葡萄糖苷酶酵母菌株的復篩
初篩中顯色較強的22株菌株(表2中顯示為D較強產酶菌株),以商業酵母D254為對照,采用1.2.3方法進行β-葡萄糖苷酶活性測定,結果見表3,其中G13酶活、G21酶活均高于對照商業酵母菌株D254(P<0.01),G8、G17、G19與對照商業酵母菌株D254的酶活力相當(P>0.05)。

表3 不同酵母菌株β-葡萄糖苷酶活性Table 3 Activity of β-glucosidase of different yeast strains
2.5 高產β-葡萄糖苷酶酵母菌株的鑒定
2.5.1 WL、賴氨酸培養基分類分析 將復篩得到的5株高產β-葡萄糖苷酶酵母菌株劃線接種于WL培養基、賴氨酸培養基,菌落特征見表4,菌株G8、G13、G17、G19、G21均為釀酒酵母(Saccharomycescerevisiae)。

表4 WL培養基和賴氨酸培養基鑒定結果Table 4 Identification results of WL medium and lysine medium
2.5.2 菌株生理生化特性
2.5.2.1 糖發酵試驗 表5是產β-葡萄糖苷酶酵母糖類發酵的試驗結果,不同酵母菌株對不同糖的利用能力不同,其中5株菌對葡萄糖、蔗糖的發酵結果均為強陽性,相比上述糖對麥芽糖、半乳糖、棉籽糖的發酵利用率較差;5株菌均不發酵乳糖。

表5 產β-葡萄糖苷酶酵母糖類發酵的試驗結果Table 5 Results of fermenting glucose by β-glucosidase yeast strains
2.5.2.2 碳源、氮源同化試驗結果 高產β-葡萄糖苷酶酵母菌株的碳、氮源同化結果見表6。G8、G13、G17、G19、G21共5株菌株的碳源同化試驗結果中,均能同化葡萄糖、蔗糖、半乳糖、麥芽糖;而5株菌對核糖、鼠李糖、乳糖、蘋果酸、檸檬酸均不能同化;氮源同化試驗結果,所有菌株均不能同化硝酸鉀、鹽酸尸胺、鹽酸乙胺。
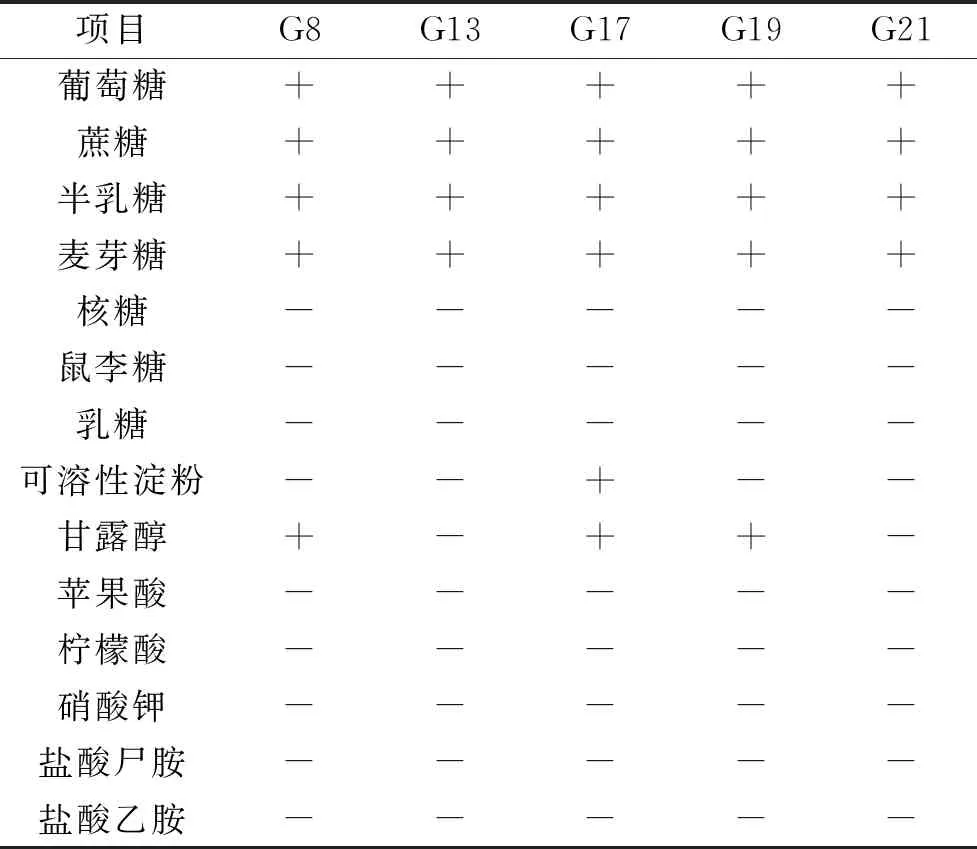
表6 產β-葡萄糖苷酶酵母菌株生理生化實驗結果Table 6 Results of physiological and biochemical on β-glucosidase yeast strains
根據發酵糖、碳源同化、氮源同化等試驗結果,并對照《The Yeasts,a Taxonomic Study》[21]對釀酒酵母的描述,菌株G8、G13、G17、G19、G21共5株菌均為酵母屬(Saccharonmyces)的釀酒酵母(Sacchromucescerevisiae)。
2.5.3 18S rDNA序列分析 通過PCR擴增產生的DNA片段為單一條帶,大小長度約為 1500 bp,擴增產物無明顯非特異擴增現象,結果見圖2。菌株18S rDNA序列經測序后提交GenBank,BLAST比對分析,選擇較為相似的序列構建系統發育樹,如圖3,G8、G13、G17、G19、G21菌株與釀酒酵母(Saccharomycescerevisiae)的18S rDNA序列同源性最高,達到100%,故確定G8、G13、G17、G19、G21菌株均為釀酒酵母(Saccharomycescerevisiae),這與WL、賴氨酸、糖類發酵、碳氮源同化分析的結果相同。

圖3 酵母菌株的系統發育樹Fig.3 The phylogenetic tree for yeast strains
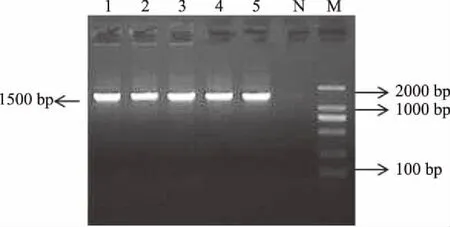
圖2 酵母菌株的18S rDNA擴增電泳結果Fig.2 Results of 18S rDNA amplify of yeast strains注:M:Maker;1、2、3、4、5分別是 G8、G13、G17、G19、G21,N:陰性質控。
2.6 不同脅迫因素對高產β-葡萄糖苷酶的菌株產酶影響分析
2.6.1 乙醇對菌株產酶影響分析 對前期實驗篩選的高產酶G13、G21菌株進行產酶分析,由圖4,隨著乙醇濃度的升高,G13、G21產β-葡萄糖苷酶的活性均隨乙醇濃度升高而逐漸下降,在10%乙醇濃度下,大約80%的酶活性被保留,乙醇濃度為12%~16%時,G13菌株在14%乙醇濃度酶活力達到37.21 U/mL,G21菌株在14%乙醇濃度酶活力達到35.51 U/mL,相比12%的乙醇濃度的酶活,兩株菌產酶活力有所上升,這可能是因為適度濃度的乙醇有助于提高細胞膜的通透性,使細胞內的酶有效釋放到胞外,從而提高菌株產酶的活力。而當乙醇濃度為16%時,兩株菌產β-葡萄糖苷酶下降到最高酶活的40%,表明G13、G21菌株具有較好的耐受乙醇能力,且在適度(12%~14%)乙醇濃度范圍產酶能力較10%乙醇濃度有所提高。

圖4 乙醇對產β-葡萄糖苷酶菌株產酶的影響Fig.4 Effect of ethanol on enzyme production of β-glucosidase yeast strains注:組內多重比較,不同字母表示差異顯著(P<0.05),相同字母表示差異不顯著(P>0.05);圖5、圖6同。
2.6.2 葡萄糖對菌株產酶影響分析 由圖5可知,隨著葡萄糖濃度的增大,兩株菌產β-葡萄糖苷酶能力逐漸減小,5%葡萄糖濃度時酶活力下降了35%,10%葡萄糖濃度時僅保留了50%的最大酶活,當含糖量達20%~25%時產酶活力僅為最佳酶活力的30%,在葡萄酒釀造中,新疆北麓葡萄酒產區葡萄醪液糖度一般為240 g/L,折合百分率糖度大致為24%,表明G13、G21兩株菌可耐受較高濃度的葡萄糖且能夠產酶,但在高糖濃度下,菌株產酶受到強烈抑制,可見高糖條件對菌株β-葡萄糖苷酶的產生極為不利。

圖5 葡萄糖對產β-葡萄糖苷酶菌株產酶的影響Fig.5 Effect of glucose on enzyme production of β-glucosidase yeast strains
2.6.3 pH對菌株產酶影響分析 從圖6可知,G13、G21菌株培養48 h,pH4.0時,G13酶活達到47.89 U/mL(P<0.05),pH5.0時,G21酶活達到最高45.62 U/mL(P<0.05),當pH3.0時G13、G21菌株均保持40%的最大酶活,說明在葡萄酒條件pH3.0~3.5條件,G13、G21菌株可發揮產酶活力。

圖6 pH對產β-葡萄糖苷酶菌株產酶的影響Fig.6 Effect of pH on enzyme production of β-glucosidase yeast strains
3 結論與討論
從新疆天山北麓釀酒葡萄產區昌吉回族自治州大三工村甲、乙和丙3個葡萄種植園中的成熟“赤霞珠”釀酒葡萄自然發酵過程中共分離到187株葡萄酒相關酵母,其中甲葡萄園分離到65株,乙葡萄園分離到70株,丙葡萄園分離到52株。在發酵的整個過程都分離到相關釀酒酵母,主要集中在發酵的前期和中期,產β-葡萄糖苷酶酶活力菌株也主要分布在發酵前期,初篩中顯色較強的22株菌株采用p-NPG法進行β-葡萄糖苷酶活性測定,并以商業酵母D254為對照,結果5株酵母菌株酶活具有較強產β-葡萄糖苷酶活力,其中G13、G21酶活均高于對照商業酵母菌株D254,G8、G17、G19酶活與對照商業酵母菌株D254的酶活力相當。復篩得到的5株產β-葡萄糖苷酶酵母菌株結合WL營養瓊脂和賴氨酸培養基、糖類發酵、碳氮源同化試驗、18S rDNA測序鑒定及系統進化樹確定五株高產β-葡萄糖苷酶酵母菌株G8、G13、G17、G19、G21均為釀酒酵母(Saccharomycescerevisiae),其中高產酶的G13、G21酵母菌株可耐受較高濃度的乙醇、一定濃度的高糖環境且在葡萄酒pH3.0~3.5條件,發揮產β-葡萄糖苷酶活力。
本研究得到的產β-葡萄糖苷酶的酵母菌株均為釀酒酵母,未篩選到非釀酒酵母,可能是因為本區成熟葡萄的含糖量高,表皮附著的酵母很快在發酵24 h適應環境,成為優勢菌株,釀酒酵母通過色素沉著將糖轉化為乙醇和二氧化碳,當然很多研究發現非釀酒酵母(假絲酵母屬Candida、漢森氏酵母屬Hanseniaspora、梅奇酵母屬Metschnikowia和畢赤酵母屬Pichia)產酶活力高于釀酒酵母[6,13,22-23],是糖苷酶的潛在來源,但在葡萄酒高糖、10%乙醇濃度、低pH下非釀酒酵母產酶活性受到抑制,而一些產β-葡萄糖苷酶的釀酒酵母,如本實驗中的G13、G21菌株耐受葡萄酒環境的影響,因此篩選不論強的或者弱的產β-糖苷酶的菌株如在葡萄酒發酵條件下能否穩定產酶對葡萄酒香氣的增香至關重要,后續研究應提高產β-葡萄糖苷酶酵母菌株的產酶活力或采取高產酶的非釀酒酵母和釀酒酵母共混發酵以提高葡萄酒的香氣及品質。
猜你喜歡
發明與創新·初中生(2024年6期)2024-06-16 00:00:00
昆明醫科大學學報(2022年1期)2022-02-28 07:43:38
中國民間療法(2021年8期)2021-07-22 05:53:28
軍事文摘·科學少年(2021年1期)2021-02-04 08:03:45
當代水產(2019年3期)2019-05-14 05:43:24
天然產物研究與開發(2018年9期)2018-10-08 03:25:32
煙草科技(2015年8期)2015-12-20 08:27:04
藥學與臨床研究(2015年4期)2015-06-05 11:35:51
閱讀與作文(小學低年級版)(2015年8期)2015-05-30 10:48:04
食品工業科技(2014年9期)2014-03-11 18:15:31
