豬生產性狀相關SNP位點研究進展
2020-06-21 15:16:40宋春雷高靈琦
南方農業(yè)·中旬 2020年1期
宋春雷 高靈琦

摘 要 單核苷酸多態(tài)性以其自身具有的一些優(yōu)良特性作為一種遺傳標記現(xiàn)已被廣泛地應用于禽畜等方面的科學研究中。基于此,簡單介紹SNP的特性和常用檢測分型的方法,并以我國主要經(jīng)濟動物-豬為中心,收集近三年國內SNP影響豬生長、肉質、繁殖等主要經(jīng)濟性狀的相關研究,從全面的角度分析豬SNP研究的相關進展,并對SNP的未來發(fā)展進行展望。
關鍵詞 豬;單核苷酸多態(tài)性(SNP);生產性狀
中圖分類號:S828 文獻標志碼:B DOI:10.19415/j.cnki.1673-890x.2020.02.076
在經(jīng)濟動物的遺傳育種過程中,科研工作者們致力于使用一些可靠的遺傳標記(Genetic Maker)來提高選擇效率與效能,提高經(jīng)濟效益,其中單核苷酸多態(tài)性(Single Nucleotide Polymorphism,SNP)現(xiàn)在已成為理想的遺傳標記之一。SNP是指在基因組上由單個核苷酸變異引起的DNA序列多態(tài)性,包括單個堿基的轉換或顛換等形式,且其中最少出現(xiàn)一種等位基因的頻率不小于1%。
我國是豬養(yǎng)殖大國,豬肉也是國民日常消費的主要肉類之一,2015年和2016年我國生豬存欄量分別為4.51億頭和4.35億頭。面對如此大的市場,只要對豬某一個重要性狀進行改良就可能帶來巨大的經(jīng)濟效益。檢測、鑒定影響重要性狀的SNPs并標記應用于育種養(yǎng)殖中,可以加快遺傳選育進度,提高經(jīng)濟效益。基于此,就SNP主要的檢測、功能活性研究方法以及在影響豬重要性狀上的研究進展進行綜述。
1 SNP的研究內容
1.1 SNP的特性
SNP自身所具備的一些優(yōu)良特性使得其在遺傳分析中成為一類能夠廣泛應用的遺傳標記,能夠更加清晰地剖分研究復雜的數(shù)量性狀、遺傳疾病,構建第三代遺傳圖譜。
SNP的優(yōu)良特性有以下4點。1)SNP位點的豐富性。SNP數(shù)量眾多且分布廣泛,幾乎遍布整個基因組,DNA序列中任意一個核苷酸都可能發(fā)生變異。據(jù)估計,基因組中每1 000個核苷酸就有1個SNP。2)SNP適于快速、規(guī)模化篩查。組成DNA的堿基雖然有4種,但因為導致SNP的通常只有兩種堿基,所以這是一種二態(tài)的標記,即二等位基因。由于SNP的二態(tài)性,非此即彼,在進行基因篩選時往往只需要進行+/-分析,而不用分析片段的長度,這就利于發(fā)展快速的規(guī)模化自動化技術篩選。3)SNP等位基因頻率易于估計。由于SNP的二態(tài)性,在任何種群中其等位基因的頻率都是可以估計的。4)SNP突變率低、遺傳穩(wěn)定性高。SNP是由基因組上單核苷酸變異引起的,每代每堿基突變率約為2×18-8。尤其是處于編碼區(qū)的SNP(cSNP)是高度穩(wěn)定的,相比較微衛(wèi)星等重復序列的多態(tài)性標記,其遺傳穩(wěn)定性更高。
1.2 SNP的檢測分型方法
1.2.1 對未知SNP進行分析
對未知SNP進行分析,即找尋未知的SNP或確定某一未知SNP與某遺傳病或性狀的關系。檢測未知SNP可以使用多種方法,如溫度梯度凝膠電泳(TGGE)、變性梯度凝膠電泳(DGGE)、單鏈構象多態(tài)性(SSCP)、限制性片段長度多態(tài)性(RFLP)和隨機擴增多態(tài)性DNA(RAPD)等,但這些方法只能發(fā)現(xiàn)含有SNP的DNA鏈,不能確知突變的位置和堿基類別,要想做到這一點,必須對那些含有SNP的DNA鏈進行測序,即基因測序技術。
1.2.2 對已知SNP進行分析
對已知SNP進行分析,即對不同群體SNP遺傳多樣性檢測或在臨床上對已知致病基因的遺傳病進行基因診斷。篩查已知SNP的方法有等位基因特異寡核苷酸片段分析(ASO)、突變錯配擴增檢驗(MAMA)和基因芯片技術(genechips)等。
1.3 豬SNP多態(tài)性分析
SNP是指在基因組上由單個核苷酸變異引起的DNA序列多態(tài)性,DNA序列直接影響氨基酸密碼子的轉錄,從而影響分子水平上蛋白質的翻譯。陳月嬋等[1]就豬基因單核甘酸多態(tài)性進行了大量的研究分析,運用了PCR-SSCP檢測方法對豬的多個基因進行了檢測分析,檢測出大量的多態(tài)位點和位點突變造成的編碼氨基酸的變化,結果如表1所示。
2 豬重要性狀SNP的研究
目前,可以通過QTL作圖對基因進行性狀連鎖分析,還可以對候選基因利用各種方法與單核苷酸變異的位點進行關聯(lián)分析。如果找到了與目的性狀有關的SNP位點,便可通過芯片篩選個體,并進行優(yōu)缺點分析,最后通過雜交、回交等改良品種。
需注意的是雖然研究的SNP與豬的重要性狀存在關聯(lián)性,但在不同品種的豬群中,具體效益可能會有所不同。
2.1 生長性狀和飼料利用SNP
生長性狀也稱肥育性狀,是中等遺傳力性狀,也是豬生產中復雜且重要的經(jīng)濟性狀。在生長性狀中以生長速度和飼料利用率為最重要。
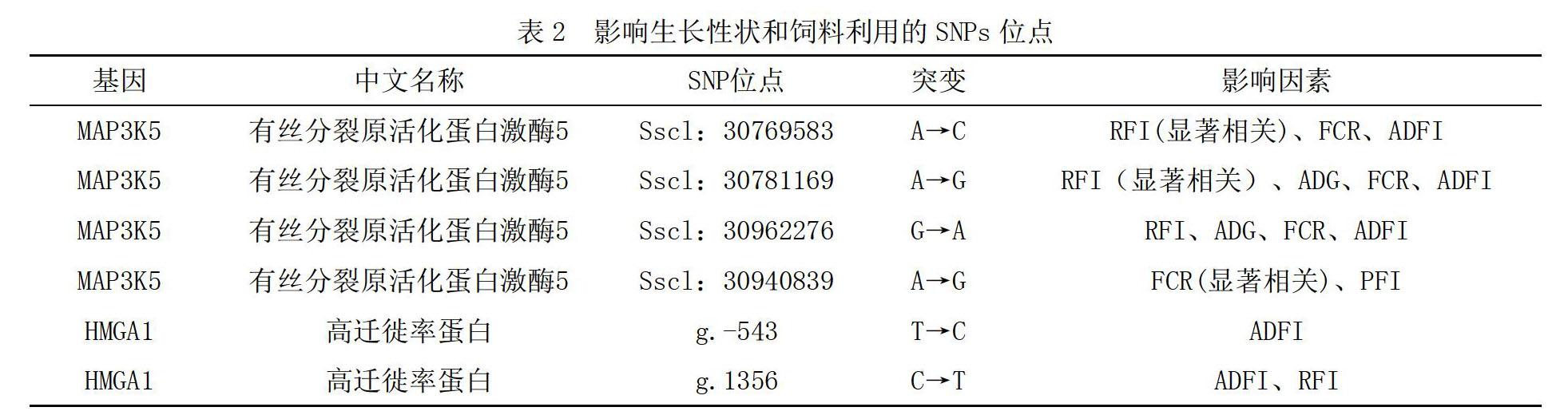
飼料成本決定了養(yǎng)殖成本,飼料利用效率的提高可以大大節(jié)約養(yǎng)殖成本。剩余采食量(Residual feed intake,RFI)是禽畜實際采食量與預期采食量的差值,也是衡量飼料轉化率(Feed conversion ration,F(xiàn)CR)的指標之一。隨著電子自動喂料技術的發(fā)明和普及,使平均日采食量(Average daily feed intake,ADFI)和平均日增重(Average daily gain,ADG)得以準確測定。基于以上條件,蒲蕾等[2]對杜洛克豬的MAP3K5基因和HMGA1基因進行了研究實驗。1)利用Illumina SNP60芯片對豬的全基因組進行分型,得到的結果和軟件基因庫中記錄的MAP3K5基因的SNP位置進行比對確定了4個SNPs位點,通過實驗數(shù)據(jù)收集發(fā)現(xiàn)4個SNPs位點中,Sscl:30769583 A>C、Sscl:30962276 G>A、Sscl:30781169 A>G位點與RFI性狀呈顯著相關,Sscl:30940839 A>G位點的突變與FCR性狀呈顯著相關。MAP3K5基因可能是通過改變豬體內的激素調節(jié)、生長因子等,來實現(xiàn)對RFI、FCR這些性狀的調控。這4個SNPs位點可記錄作為分子標記。2)通過對大白豬和民豬群體的重測數(shù)據(jù)分析,發(fā)現(xiàn)了HMGA1基因的2個SNPs位點。并采用質譜分型技術對杜洛克豬群體進行這2個SNPs位點的基因分型。收集統(tǒng)計相關數(shù)據(jù)后得出HMGA1基因g.-543 T>C和g.1356 C>T位點突變可以使杜洛克豬在不影響ADG的同時降低同期間內的ADFI,即同日齡個體體重更重,同體重個體日齡更短。尤其g.1356 C>T位點突變還可以使杜洛克豬有更薄的背膘厚和更低的RFI。這2個SNPs位點可記錄作為分子標記,為豬遺傳育種提供輔助參考。如表2所示,為影響生長性狀和飼料利用的SNPs位點。
2.2 繁殖性狀SNP
豬繁殖性狀包括母豬的窩產仔數(shù)、初生活仔數(shù)、仔豬初生重、乳頭數(shù)、泌乳力、斷奶時育成仔豬數(shù)、斷奶窩重等。公豬的繁殖性狀主要包括精液的質量、精子的活力、精液的產量、公豬性欲以及公豬使用年限。
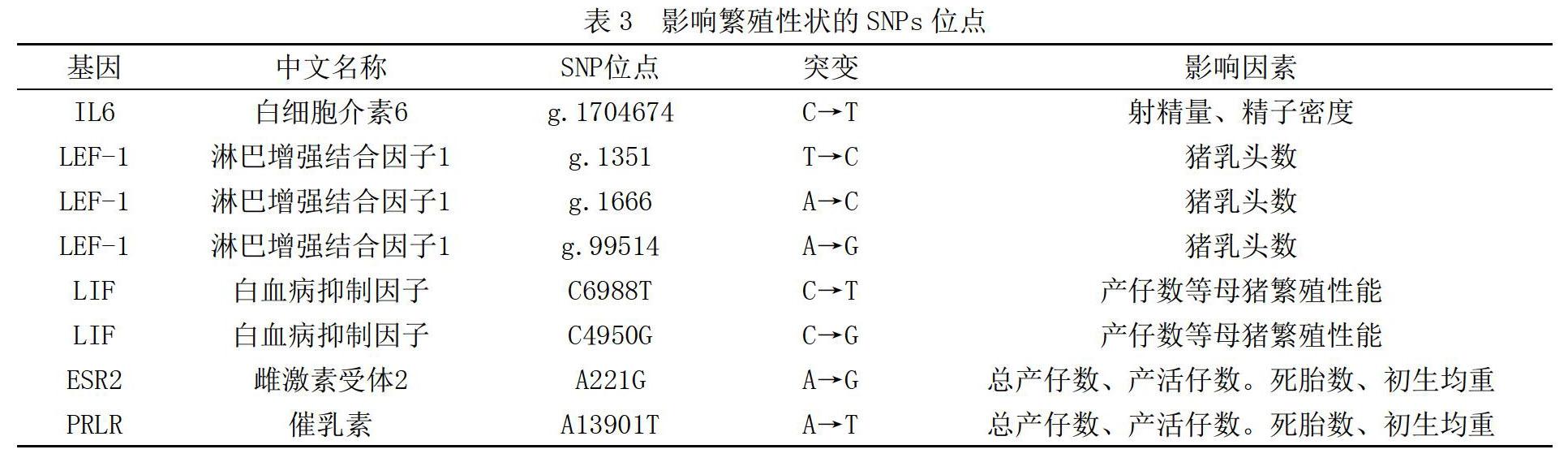
查安東等[3]在IL6基因內檢測到1個SNP位點g.1704674 C>T,并通過PCR-RFLP檢測其分型。統(tǒng)計結果發(fā)現(xiàn),在杜洛克豬種g.1704674 C>T位點與射精量呈顯著相關(P<0.05),T等位基因為優(yōu)勢等位基因;而在大白豬中,g.1704674 C>T位點與精子密度呈顯著相關(P<0.05),說明SNP對公豬繁殖性狀有一定影響。Tetzlaff等[4]研究發(fā)現(xiàn)了在LEF-1基因中有2個SNPs(g.1351 T>C、g.1666 A>C)與豬乳頭數(shù)顯著相關,而后Xu等[5]在該基因上再次檢測到新的突變位點(g.99514 A>G、g.119846 C>T),關聯(lián)分析后發(fā)現(xiàn)均與豬乳頭數(shù)顯著相關。胡閃耀等[6]就此展開研究,以美系大白豬為研究對象,發(fā)現(xiàn)僅g. 99514 A>G位點與豬乳頭數(shù)顯著相關,而另一位點的相關性并沒有達到顯著水平,對乳頭數(shù)變異沒有影響推測,可能只是連鎖性的標記。劉碧霞等[7]研究發(fā)現(xiàn),大白母豬LIF基因上C6988T位點的CT基因型和C4950G位點的GC基因型為繁殖性狀的增效基因型,2個SNPs對繁殖性狀的影響呈現(xiàn)了明顯的協(xié)同效應,即這2個SNPs與母豬產仔數(shù)等繁殖性狀有顯著關聯(lián)。高天[8]等在嘉興黑豬ESR2、FSHβ和PRLR基因中篩選并確定了8個SNPs,并與母豬的繁殖性狀展開了關聯(lián)分析,分析發(fā)現(xiàn)ESR2基因中的A221G位點和PRLR基因中的A13901T位點均與個體繁殖性狀呈顯著關聯(lián),而其余位點并未發(fā)現(xiàn)有顯著關聯(lián)。如表3所示,為影響繁殖性狀的SNPs位點
2.3 肉質性狀SNP
肉質的優(yōu)劣是通過許多肉質指標來判定的,常見的有pH、肉色、系水力、大理石紋、肌肉脂肪含量和公豬膻味等指標。我國種豬遺傳評估方案中的肉質性狀有:肌肉pH、肉色、滴水損失和大理石紋。
喬木等[9]通過測序,發(fā)現(xiàn)豬RTL1基因中存在1個SNP位點,利用PCR-Fnu4H I-RFLP方法在9個豬種中進行了分型,并在360頭大×梅F2代群體中進行了性狀關聯(lián)分析。結果表明,在肉質性狀方面該位點與背最長肌pH、股二頭肌肌肉肉色值呈顯著相關(P<0.01),與背最長肌系水力、背最長肌肌肉色值呈極顯著相關(P<0.05)。楊華等[10]采用PCR-Msp I-RFLP方法對IGFBP2基因的g.171 C>T位點進行分型,關聯(lián)分析得出g.171 C>T位點與肉質性狀顯著相關。張越等[11]研究了H-FABP基因對肉質性狀的影響,通過測序確定了2個SNPs位點,命名為SNP1和SNP2,研究顯示SNP1對肉質性狀沒有顯著影響,SNP2僅與谷氨酸含量顯著關聯(lián),谷氨酸作為影響豬肉的主要鮮味物質之一,對肉質性狀有影響,可作為參考位點進行深入研究。如表4所示,為影響肉質性狀的SNPs位點。
2.4 胴體性狀SNP
胴體性狀是一種高遺傳力性狀,豬的胴體性狀主要有背膘厚度、胴體長度、眼肌面積、腿臀比例和胴體瘦肉率等。
喬木等[9]研究的RTL1基因,同一個SNP位點,檢測出的AA、AG、GG這3個分型中,AA基因型個體的內脂率、平均背膘厚、肩部膘厚等顯著高于GG基因型,參考不同豬種中基因型的分布規(guī)律,國內豬種中A等位基因占比明顯高于國外豬種,因而具有較高的肥肉率,該SNP位點突變具有促進脂肪沉積的效應,可作為改善豬胴體性狀的分子標記。楊華等[10]研究的IGFBP2基因的g.171 C>T位點,關聯(lián)分析也得出g.171 C>T位點與胴體性狀顯著相關。如表5所示,為影響胴體性狀的SNPs位點。
3 SNP存在的問題與展望
SNPs研究現(xiàn)在正處于發(fā)展之中,雖然其具有很好的前景,但目前仍存在以下4點問題。
1)SNPs改變了基因原來的結構和連鎖率,主要表現(xiàn)為生物對外界反應的不適應。所以,隨著SNP的增加,可能會相應導致致命性疾病的增加。
2)雖然目前已經(jīng)有了幾個公開的SNP數(shù)據(jù)庫,但由于一些商業(yè)利益的考慮,不是所有的科研工作者都可以自由詳盡地利用SNP數(shù)據(jù)庫,勢必將阻礙SNP信息的獲取與利用。
3)目前雖然有大量檢測SNP的方法,且已經(jīng)建立了高度自動化和高通量的SNP檢測分型技術,但大都價格昂貴,速度較慢,這限制了其在科學研究中的應用,所以迫切需要有低成本、高生產率的新方法涌現(xiàn)。
4)SNP命名問題。SNP的命名是很混亂的,從本文的信息整理就可以看出來,不同的研究者采用不同的記錄方法,目前沒有一個統(tǒng)一標準的命名方法,不同的組織機構命名不一樣,并且各自為政,堅持自己的命名方法,由于不同SNP由不同實驗室測定,現(xiàn)在至少存在6種不同的命名系統(tǒng)。這為后人學習以及科研成果統(tǒng)計造成了很大的障礙。
4 結語
雖然目前SNP的應用還存在一些不足,但依然存在很大的潛力。可以預期,SNP的應用將促進分子標記技術與其他生物技術的結合,大大加速傳統(tǒng)育種技術的革新,隨著分子生物技術的提高,SNP的應用前景將更加廣泛。
參考文獻:
[1] 陳月嬋,翟春媛,劉娣,等.豬TLR4基因2個突變位點的PCR-SSCP分析[J].黑龍江畜牧獸醫(yī),2012(17):
1-3.
[2] 蒲蕾,劉欣,岳靜偉,等.杜洛克豬HMGA1基因多態(tài)位點與生長、飼料利用性狀的關聯(lián)分析[J].中國畜牧獸醫(yī),2016,43(12):3268-3274.
[3] 查安東,邢桂玲,高曉雯,等.IL6基因內含子3多態(tài)性與仔豬腹瀉、生長和公豬繁殖性狀的關聯(lián)分析[J].畜牧獸醫(yī)學報,2017,48(9):1674-1682.
[4] Tetzlaff S,Jonas E,Phatsara C,et al.Evidence for association of lymphoid enhancer-binding factor-1 (LEF1) with the number of functional and inverted teats in pigs[J].Cytogenetic & Genome Research,2009,124(2):139-146.
[5] Xu R X,Wei N,Wang Y,et al.Association of Novel Polymorphisms in Lymphoid Enhancer Binding Factor 1 (LEF-1) Gene with Number of Teats in Different Breeds of Pig[J].Asian Australasian Journal of Animal Sciences,2014,27(9):1254-1262.
[6] 胡閃耀,楊華,姜佳佳,等.美系大白豬LEF-1基因多態(tài)性與乳頭數(shù)及生長性狀的相關性研究[J].豬業(yè)科學,2017,34(1):116-117.
[7] 劉碧霞,楊善海,嚴達偉,等.大白母豬LIF基因多態(tài)性及其對繁殖性狀的影響[J].云南農業(yè)大學學報(自然科學),2017,32(4):620-626.
[8] 高天,鐔忠斌,王曉杜,等.嘉興黑豬ESR2和FSHβ及PRLR基因多態(tài)性與繁殖性狀相關性分析[J].中國畜牧雜志,2016,52(5):1-7.
[9] 喬木,程碧軍,武華玉,等.豬RTL1基因多態(tài)性及其與胴體、肉質性狀的關聯(lián)分析[J].湖北農業(yè)科學,2017,56(20):3948-3951.
[10] 楊華,徐珍,左波.豬IGFBP2基因多態(tài)性及其與胴體、肉質性狀的關聯(lián)分析[J].中國畜牧雜志,2017,53(3):
25-28.
[11] 張越,陸萌,朱衛(wèi)華,等.豬心臟脂肪酸結合蛋白基因多態(tài)性檢測及與肉質性狀的關聯(lián)分析[J].中國畜牧雜志,2016,52(1):7-12.
(責任編輯:趙中正)
