基于宏轉(zhuǎn)錄組學(xué)技術(shù)解析工業(yè)豇豆泡菜發(fā)酵過程中活性微生物群落結(jié)構(gòu)變化
2020-06-12 09:00:04魏雯麗宮尾茂雄吳正云張文學(xué)
食品與發(fā)酵工業(yè) 2020年10期
魏雯麗,宮尾茂雄,吳正云,張文學(xué),2*
1(四川大學(xué) 輕工科學(xué)與工程學(xué)院,四川 成都,610065)2(四川大學(xué)錦江學(xué)院 白酒學(xué)院,四川 眉山,620860) 3(東京家政大學(xué) 家政學(xué)部,日本 東京,173-8602)
泡菜作為傳統(tǒng)發(fā)酵蔬菜制品的代表,因其味道酸爽、質(zhì)地脆生、營養(yǎng)豐富等特點(diǎn)受到廣大消費(fèi)者的青睞[1-2]。目前,泡菜的生產(chǎn)模式逐漸從傳統(tǒng)家庭作坊轉(zhuǎn)變?yōu)楣I(yè)化生產(chǎn),工業(yè)泡菜的發(fā)酵工藝根據(jù)蔬菜的含水量分為兩類,一類是含水量較高的蔬菜(如蘿卜和青菜)直接用高鹽腌制[3],另一類是用泡菜母水泡制豇豆等含水量較低的蔬菜。與其他發(fā)酵蔬菜類似,工業(yè)豇豆泡菜也是一個(gè)多菌混合發(fā)酵體系,復(fù)雜的微生物群落對于豇豆泡菜獨(dú)特的口感、風(fēng)味和質(zhì)地起到了極其重要的作用。
近年來,泡菜中微生物群落結(jié)構(gòu)在國內(nèi)外得到廣泛關(guān)注,LIANG等[4]利用變性梯度凝膠電泳(polymerase chain reaction-denatured gradient gel electrophoersis,PCR-DGGE)技術(shù)探究了工業(yè)青菜發(fā)酵過程中微生物,結(jié)果表明乳桿菌屬(Lactobacillus)、假單胞菌屬(Pseudomonas)、弧菌屬(Vibrio) 和鹽單胞菌屬(Halomonas)是優(yōu)勢細(xì)菌屬和德巴利氏酵母屬 (Debaryomyces) 是優(yōu)勢真菌屬;朱琳等[5]通過基于16S rDNA的高通量測序在蘿卜泡菜中檢測到腸膜明串珠菌屬、乳球菌屬、魏斯氏菌屬和片球菌屬;LEE等[6]運(yùn)用宏基因組學(xué)技術(shù)發(fā)現(xiàn)明串珠菌屬(Leuconostoc)、乳桿菌屬(Lactobacillus)和魏斯氏菌屬(Weissella)是韓國泡菜發(fā)酵過程中的優(yōu)勢菌屬。但這些方法僅停留在DNA層面上,已經(jīng)死亡的微生物和處于非活躍狀態(tài)的微生物也包括在內(nèi),并不能真實(shí)反映發(fā)酵過程中真正發(fā)揮作用的微生物群落組成情況。
宏轉(zhuǎn)錄組學(xué)技術(shù)是以樣品中全部微生物RNA信息為分析對象,從轉(zhuǎn)錄組水平分析樣本微生物的組成及活性基因表達(dá)情況,其結(jié)果反映的是活性微生物的群落結(jié)構(gòu)[7-8]。近年來,宏轉(zhuǎn)錄組學(xué)技術(shù)已用于傳統(tǒng)酸面團(tuán)[7]和奶酪[9]的研究,本研究以工業(yè)豇豆泡菜為研究對象,運(yùn)用宏轉(zhuǎn)錄組學(xué)技術(shù)對發(fā)酵過程中活性微生物群落結(jié)構(gòu)進(jìn)行探究,以確定其優(yōu)勢活性微生物組成,同時(shí)對發(fā)酵過程中功能基因的表達(dá)情況進(jìn)行了探究,以期為對工業(yè)泡菜發(fā)酵機(jī)理解析及發(fā)酵過程控制提供理論基礎(chǔ)。
1 材料與方法
1.1 材料與試劑
理化試劑:NaOH、亞鐵氰化鉀、乙酸鋅、四硼酸鈉、對氨基苯磺酸、NaNO2、鹽酸萘乙二胺(均為分析純),成都市科龍化工試劑廠。
測序試劑:RNA PowerSoil?Total RNA Isolation Kit 提取試劑盒,美國 Mobio公司;Ribo-Zero rRNA Removal Kit,美國EPICENTRE公司; TruSeq Stranded mRNA LT Sample Prep Kit,美國Illumina公司; Agilent High Sensitivity DNA Kit,美國安捷倫公司。
1.2 儀器與設(shè)備
PHS-3C酸度計(jì),上海儀電科學(xué)儀器股份有限公司;WS202鹽度計(jì),上海民儀電子有限公司;LC2030高效液相色譜儀,日本島津;瓊脂糖凝膠電泳儀,中國北京市六一儀器廠;高速冷凍離心機(jī),美國賽默飛公司;Agilent 2100 Bioanalyzer,美國安捷倫公司; PCR儀,美國Bio-Rad公司;Illumina Hiseq 10X測序平臺 美國Illumina 公司。
1.3 實(shí)驗(yàn)方法
1.3.1 樣品采集與處理
分別采集發(fā)酵1、15、30、60、90 d的豇豆泡菜液(四川省眉山市某泡菜工廠),取樣后冰袋運(yùn)回實(shí)驗(yàn)室,抽濾過膜富集微生物置于無菌無酶離心管中,液氮速封后轉(zhuǎn)移至-80 ℃冰箱備用。該工廠豇豆泡菜發(fā)酵工藝大致為將含鹽量約10%(質(zhì)量分?jǐn)?shù))的泡菜母水與新鮮豇豆按1∶1的質(zhì)量比投入發(fā)酵池,木棍壓實(shí),在相對開放的環(huán)境中發(fā)酵3個(gè)月。
1.3.2 泡菜理化指標(biāo)測定
pH、鹽度分別使用pH計(jì)和鹽度計(jì)進(jìn)行測定;總酸參照GB/T 12456—2008食品總酸的測定[10]中酸堿滴定法測定,結(jié)果以乳酸計(jì);亞硝酸鹽含量采用 GB5009.33—2016食品安全國家標(biāo)準(zhǔn)食品中亞硝酸鹽與硝酸鹽的測定[11]測定。采用高效液相色譜法測定乳酸,乙酸和葡萄糖含量。測定條件為色譜柱Carbomix H-NP,7.8 mm×300 mm,5 μm;流動相0.25 mol/L H2SO4,流速0.6 mL/min, 柱溫55 ℃, 注射量10 μL。紫外檢測器波長210 nm檢測乳酸和乙酸, 示差檢測器檢測葡萄糖,通過與標(biāo)準(zhǔn)品的保留時(shí)間和峰面積的比較,對其進(jìn)行鑒定和定量。以上實(shí)驗(yàn)均做3次平行測定。
1.3.3 泡菜液樣品RNA提取及rRNA的去除
使用RNA PowerSoil?Total RNA Isolation Kit試劑盒提取泡菜液樣品的總RNA, 用1.5%(質(zhì)量分?jǐn)?shù))的瓊脂糖凝膠電泳檢測RNA的完整性,然后用紫外分光光度計(jì)定量。將rRNA 序列特異性的探針與總RNA雜交,用磁珠去除rRNA/探針復(fù)合物,再用乙醇沉淀法純化mRNA。
1.3.4 cDNA文庫構(gòu)建及測序
采用TruSeq Stranded mRNA LT Sample Prep Kit試劑盒構(gòu)建cDNA文庫,用Agilent 2 100 Bioanalyzer對構(gòu)建的文庫質(zhì)檢,再用Promega QuantiFluor對文庫定量。將需要上機(jī)的文庫梯度稀釋,并按所需數(shù)據(jù)量比例混樣,混合后的文庫變性解螺旋成單鏈。基于 Illumina Hiseq 10X測序平臺,采用全轉(zhuǎn)錄組鳥槍法策略,對上述構(gòu)建的測序文庫進(jìn)行2×150 bp的雙端測序,此步驟委托上海派森諾生物公司進(jìn)行。
1.3.5 測序數(shù)據(jù)分析
原始數(shù)據(jù)經(jīng)過質(zhì)控,剔除rRNA序列后獲得高質(zhì)量reads, 使用Trinity對所有測序讀段進(jìn)行從頭組裝[12],采用CD-HIT對各樣本組裝拼接后得到的轉(zhuǎn)錄本以95%的相似度和90%的覆蓋率進(jìn)行歸并去冗余, 并以最長的序列作為該 Unigene 的代表序列。將每個(gè)樣本的Unigene序列與NCBI-NT數(shù)據(jù)庫進(jìn)行BLASTN比對(設(shè)置期望值E-value<0.001),利用MEGAN軟件采取最低共同祖先算法采用獲得物種注釋[13],結(jié)合 Unigene 序列在各樣本中的表達(dá)量數(shù)據(jù),統(tǒng)計(jì)物種在各個(gè)樣本中的表達(dá)豐度,獲得在門和屬分類學(xué)水平上活性菌群結(jié)構(gòu)組成分布。將Unigene 序列集上傳至 KAAS,與KEGG代謝通路數(shù)據(jù)庫比對,從而獲得各 Unigene 的功能注釋和分類信息[14]。
2 結(jié)果與分析
2.1 工業(yè)豇豆泡菜發(fā)酵過程中理化特性變化
工業(yè)豇豆泡菜發(fā)酵過程中理化指標(biāo)變化如圖1所示。發(fā)酵第1天pH值為4.78,發(fā)酵15 d后迅速下降至4.0以下,隨后維持在3.50左右。隨著發(fā)酵進(jìn)行,總酸含量明顯上升,發(fā)酵30 d達(dá)到最高10.01 g/kg, 之后略有下降,到發(fā)酵結(jié)束時(shí)總酸含量為8.32 g/kg。與總酸質(zhì)量濃度變化趨勢相似,發(fā)酵前30 d,乳酸質(zhì)量濃度顯著增加至9.02 g/L,隨后逐漸下降。另外,整體上,乙酸含量逐漸增加而葡萄糖質(zhì)量濃度從3.76 g/L逐漸降低至0.22 g/L。發(fā)酵前期,泡菜母水及豇豆表面存在的乳酸菌等微生物利用新鮮豇豆中的葡萄糖等糖類產(chǎn)生乳酸和乙酸等有機(jī)酸使得總酸含量快速增加,pH下降,發(fā)酵后期,由于乳酸積累過多,抑制了乳酸菌的生長代謝,同時(shí)微生物為了克服乳酸脅迫而消耗乳酸,導(dǎo)致乳酸及總酸含量下降。發(fā)酵過程中的鹽度從11%逐漸上升直至發(fā)酵結(jié)束達(dá)到13.1%。這可能是該工廠在泡制豇豆時(shí)未進(jìn)行完全封池,隨著發(fā)酵進(jìn)行,發(fā)酵池中的水分蒸發(fā),鹽度上升。亞硝酸鹽含量隨著發(fā)酵的進(jìn)行逐漸下降,亞硝酸鹽主要來自豇豆原料和雜菌代謝產(chǎn)生,隨著乳酸菌的生長代謝產(chǎn)生乳酸、 乙酸和酒石酸等有機(jī)酸提供酸性發(fā)酵環(huán)境不僅抑制雜菌生長還能使亞硝酸鹽發(fā)生氧化還原反應(yīng),同時(shí)部分微生物產(chǎn)生亞硝酸還原酶和細(xì)菌素也對亞硝酸鹽的降解起著重要作用[15-16]。本次取樣未能檢測出亞硝峰,推測可能在1~15 d出現(xiàn)亞硝峰。發(fā)酵過程中的亞硝酸鹽含量都遠(yuǎn)低于GB 2762—2017 食品中污染物限量腌漬蔬菜亞硝酸鹽限量規(guī)定的20 mg/kg[17]。
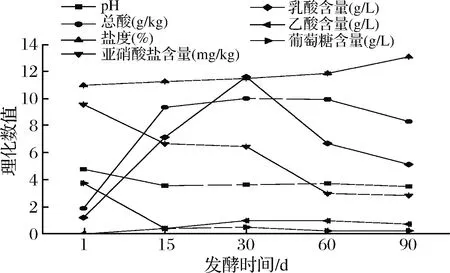
圖1 工業(yè)豇豆泡菜發(fā)酵過程中理化特性變化Fig.1 The changes of physicochemical properties of industrial cowpea paocai during the fermentation
2.2 序列組裝統(tǒng)計(jì)及α多樣性指數(shù)分析
測序后的原始數(shù)據(jù)經(jīng)過質(zhì)控,得到高質(zhì)量序列后由Trinity 軟件進(jìn)行拼接組裝,相應(yīng)統(tǒng)計(jì)結(jié)果如表1所示。5個(gè)豇豆泡菜樣本分別獲得 183 858、90 693、96 263、48 246和19 955條序列且拼接效果較好。為了比較5個(gè)樣本的多樣性,使用 QIIME 軟件對其計(jì)算α多樣性指數(shù),結(jié)果如表2所示。Chao1指數(shù)和ACE指數(shù)衡量物種豐度,Shannon 指數(shù)和Simpson 指數(shù)衡量物種多樣性和均勻度[18]。由表2可知,發(fā)酵第1天的Chao1和ACE指數(shù)最大,表明發(fā)酵第1天的活性菌群豐富度最高,Shannon指數(shù)值和Simpson指數(shù)值也比較高,表明發(fā)酵第1天活性菌群多樣性較高,而發(fā)酵90 d所有α多樣性指數(shù)都最低,表明發(fā)酵后期活性菌群多樣性和豐富度較低,同時(shí)也說明在發(fā)酵過程中活性微生物群落演替逐漸趨于穩(wěn)定。
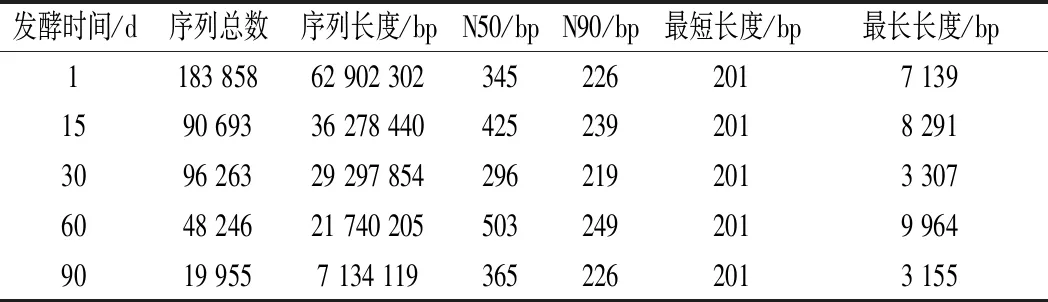
表1 序列組裝結(jié)果統(tǒng)計(jì)表Table 1 Statistics of sequence assembly results
注:N50/N90長度越長,說明組裝拼接效果越好,得到的宏轉(zhuǎn)錄組序列越完整;其中,N50長度尤為重要,是衡量宏轉(zhuǎn)錄組組裝拼接效果好壞的主要指標(biāo)之一

表2 α多樣性指數(shù)分析表Table 2 Analysis of α diversity index
2.3 物種注釋和豐度分析
工業(yè)豇豆泡菜發(fā)酵過程中活性微生物在門水平上群落結(jié)構(gòu)演替如圖2所示。發(fā)酵第1天,厚壁菌門(Firmicutes)的相對豐度高達(dá)88%,此外還檢出少量變形菌門(Proteobacteria, 7.9%)。發(fā)酵第15天,厚壁菌門的相對豐度下降而子囊菌門(Ascomycota)的相對豐度上升,分別為57.8%和41.7%。發(fā)酵30 d后,厚壁菌門以80%的相對豐度再次成為絕對優(yōu)勢菌門。隨著發(fā)酵進(jìn)行,厚壁菌門的相對豐度又逐漸降低而子囊菌門的相對豐度顯著增加,最終,子囊菌門分別以77%和98% 的相對豐度主導(dǎo)了發(fā)酵60 d和發(fā)酵90 d。隨著發(fā)酵時(shí)間的延長,原料中的營養(yǎng)物質(zhì)減少,代謝產(chǎn)物增加,微生物相互競爭,厚壁菌門中部分菌屬對泡菜液體系不適應(yīng)逐漸被淘汰,而子囊菌門中耐酸耐鹽的真菌開始在發(fā)酵后期發(fā)揮作用。
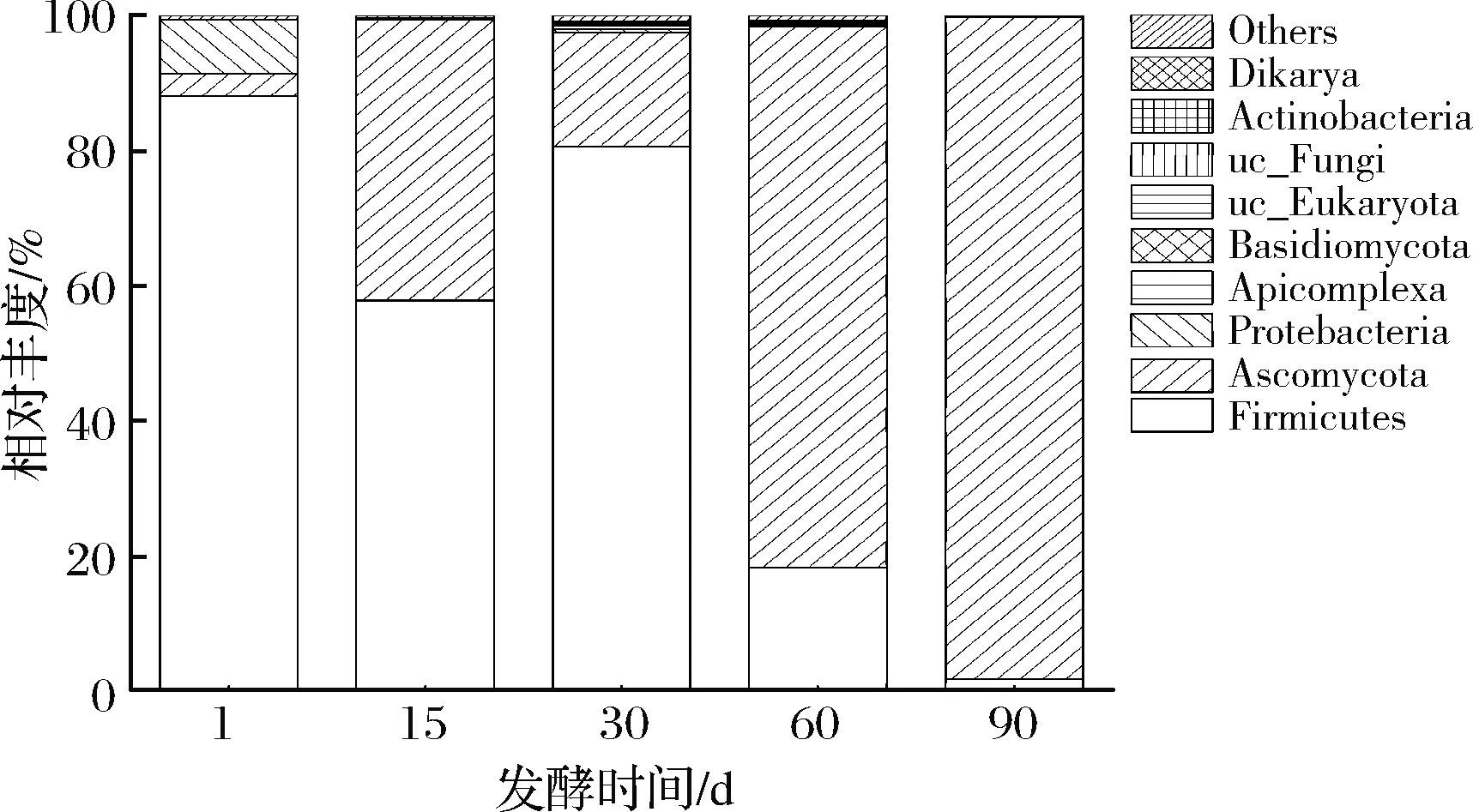
圖2 工業(yè)豇豆泡菜發(fā)酵過程中門水平上活性微生物群落結(jié)構(gòu)Fig.2 Structure of the active microbial community on phylum level during the industrial cowpea paocai fermentation
從屬水平來看(圖3),發(fā)酵第1天相對豐度最高的是乳桿菌屬(Lactobacillus, 61.8%),其次是厚壁菌門的片球菌屬(Pediococcus),其相對豐度為19.7%,此外還檢測到變形菌門的弧菌屬(Vibrio)和厚壁菌門的魏斯氏菌屬(Weissella),其相對豐度分別為7%和4.65%。乳桿菌屬(Lactobacillus)、片球菌屬(Pediococcus)和魏斯氏菌屬(Weissella)主要來自泡菜母水和新鮮豇豆,通過同型或異型發(fā)酵產(chǎn)生乳酸和乙酸,使得發(fā)酵前期總酸含量迅速上升,對豇豆泡菜酸爽口感的形成有重要作用,這些菌屬在腌菜[19]、家庭泡菜[20]和東北酸菜[21]等發(fā)酵蔬菜中也都有發(fā)現(xiàn)。發(fā)酵第15天時(shí),乳桿菌屬(Lactobacillus)的相對豐度下降至55.6%,而子囊菌門的Millerozyma的相對豐度上升至33%,Millerozyma是一種罕見的酵母病原體[22],但也有研究報(bào)道Millerozyma的某些菌種具有降解生物胺的作用[23],Millerozyma在豇豆泡菜發(fā)酵過程中的作用有待進(jìn)一步分析。另外,還發(fā)現(xiàn)了少量子囊菌門的Starmerella(4.34%),而發(fā)酵初期相對豐度較高的片球菌屬(Pediococcus)、弧菌屬(Vibrio)和魏斯氏菌屬(Weissella)的相對豐度均低于0.05%,說明這些菌屬是工業(yè)豇豆泡菜發(fā)酵的啟動微生物,參與發(fā)酵初始階段,對發(fā)酵中后期作用較小,這與之前佟婷婷等[24]發(fā)現(xiàn)魏斯氏菌屬是泡菜母水作引子發(fā)酵蔬菜過程中的啟動菌結(jié)果保持一致。隨著發(fā)酵進(jìn)行,發(fā)酵30 d時(shí)乳桿菌屬(Lactobacillus)再次成為絕對優(yōu)勢菌屬,其相對豐度為77.6%,另外,Millerozyma的相對豐度快速下降至7%。發(fā)酵第60天,Starmerella的相對豐度迅速增加到69.3%,成為主導(dǎo)菌屬,同時(shí)乳桿菌屬(Lactobacillus)的相對豐度明顯下降,但仍以16.6%的相對豐度成為第二優(yōu)勢菌屬,在發(fā)酵前中期乳桿菌屬生長代謝活躍產(chǎn)生大量乳酸,由于乳酸積累過多且發(fā)酵后期鹽度上升,乳桿菌生長代謝受到抑制。到發(fā)酵90 d時(shí),Starmerella以76.6%的相對豐度成為絕對優(yōu)勢菌屬,其次是子囊菌門的Sugiyamaella(7.1%)和Millerozyma(6.05%),僅有1.14%的乳桿菌屬(Lactobacillus)在發(fā)酵末期被檢測出。發(fā)酵后期,Starmerella逐漸占據(jù)主導(dǎo)地位,有關(guān)Starmerella報(bào)道較少,其中Starmerellabacillaris產(chǎn)甘油和揮發(fā)性酸的能力較強(qiáng),被用于發(fā)酵葡萄酒[25-26]。與本研究結(jié)果不同,李恒等[27]發(fā)現(xiàn)Kazachstania和假絲酵母屬(Candida)是泡菜母水中的主要真菌屬,可能是檢測方法、泡菜品種、發(fā)酵工藝、取樣時(shí)間及取樣地點(diǎn)不同導(dǎo)致。

圖3 工業(yè)豇豆泡菜發(fā)酵過程中屬水平上活性微生物群落結(jié)構(gòu)Fig.3 Structure of the active microbial community on genus level during the industrial cowpea paocai fermentation
2.4 KEGG功能注釋
各樣本在KEGG第一等級上的表達(dá)量分布如圖4所示。發(fā)酵第1天,有關(guān)代謝(metabolism) 的基因豐度最高,占比 48.7%,其次是遺傳信息處理(genetic information processing) 和環(huán)境信息處理 (environmental information processing),分別占17.3%和11.4%。在發(fā)酵15~30 d的轉(zhuǎn)錄本中注釋到有關(guān)代謝的基因最多,分別占 41%和48%,其次是人類疾病(human diseases),分別占23.9%和13%。隨著發(fā)酵進(jìn)行,代謝相關(guān)基因的表達(dá)量逐漸下降,發(fā)酵60 d時(shí),代謝相關(guān)基因的相對表達(dá)量為32.9%,仍維持較高的表達(dá)量。但到發(fā)酵結(jié)束時(shí),人類疾病相關(guān)基因的相對表達(dá)量達(dá)46%,生物體系統(tǒng)(organismal systems)次之,占比31%,而有關(guān)代謝的基因表達(dá)量僅占19%。在豇豆泡菜發(fā)酵前中期,以乳桿菌屬為主的微生物利用新鮮豇豆中的營養(yǎng)物質(zhì)迅速生長并通過不同代謝途徑生成各種影響豇豆泡菜品質(zhì)的代謝產(chǎn)物,使得此發(fā)酵期間代謝相關(guān)基因的表達(dá)量較高。在發(fā)酵后期,人類疾病相關(guān)基因的表達(dá)量增加,在此階段真菌豐度也迅速增加,由此推測部分真菌可能與人類疾病密切相關(guān)。
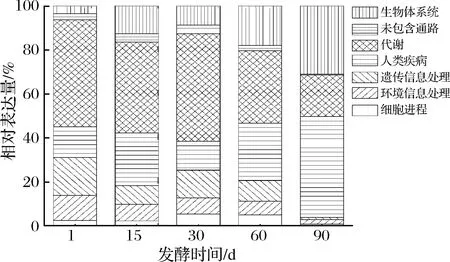
圖4 工業(yè)豇豆泡菜發(fā)酵過程中基因表達(dá)的 KEGG第一功能等級富集分析Fig.4 KEGG analysis of gene expression on first level during the industrial cowpea paocai fermentation
對代謝子功能進(jìn)行具體分析,如圖5所示,共注釋到11種代謝功能。發(fā)酵第1天,有關(guān)碳水化合物代謝(carbohydrate metabolism)和能量代謝(energy metabolism)的基因表達(dá)量較高,分別為17.9%和16.9%,此外還注釋到少量的核酸代謝(nucleotide metabolism)和氨基酸代謝(amino acid metabolism)。發(fā)酵第15天,碳水化合物代謝和能量代謝相關(guān)基因的相對表達(dá)量分別為18.9%和11.8%,另外還注釋到3%的氨基酸代謝。發(fā)酵第30天,有關(guān)碳水化合物代謝和能量代謝的基因相對表達(dá)量分別下降至18.2%和7.5%,而氨基酸代謝相關(guān)基因的表達(dá)量上升至7.4%。發(fā)酵60 d,碳水化合物代謝相關(guān)基因表達(dá)量繼續(xù)下降至11.1%,能量代謝相關(guān)基因表達(dá)量開始增加,占9.5%。到發(fā)酵90 d時(shí),能量代謝的相關(guān)基因表達(dá)量占15.9%,而碳水化合物代謝僅注釋到1.2%。除發(fā)酵第90天外,碳水化合物代謝相關(guān)基因表達(dá)量在整個(gè)發(fā)酵過程中都是最高的。發(fā)酵第1天,新鮮豇豆含大量以糖類為主的碳水化合物,乳桿菌屬,片球菌屬和魏斯氏菌屬等乳酸菌迅速生長,并通過糖酵解等代謝途徑利用葡萄糖產(chǎn)生丙酮酸等代謝產(chǎn)物,使得碳水化合物代謝相關(guān)基因表達(dá)量較高。發(fā)酵15~30 d時(shí),乳桿菌屬等微生物繼續(xù)活躍參與碳水化合物代謝,并進(jìn)一步將發(fā)酵初期積累的丙酮酸及其他中間代謝產(chǎn)物生成乳酸和乙酸等風(fēng)味成分,豇豆泡菜中乳酸和乙酸含量在此發(fā)酵階段迅速增加,葡萄糖含量顯著下降,大量乳酸和乙酸的積累使得發(fā)酵前30 d的總酸含量迅速增加,pH快速下降。發(fā)酵60 d后,隨著葡萄糖等糖類基本耗盡且乳桿菌屬豐度明顯降低,碳水化合物代謝相關(guān)基因表達(dá)量逐漸降低,尤其是乳桿菌屬含量很低的發(fā)酵90 d, 說明豇豆泡菜發(fā)酵過程中乳桿菌屬是參與碳水化合物代謝的主要菌屬。發(fā)酵前30 d還有少量氨基酸代謝,這可能是因?yàn)槲⑸飳⑿迈r豇豆中的蛋白質(zhì)分解成氨基酸,部分氨基酸可能在微生物氨基酸脫羧酶的作用下生成生物胺。此外,在整個(gè)發(fā)酵過程中能量代謝相關(guān)基因表達(dá)量較為穩(wěn)定,這是因?yàn)槲⑸镄枰S持正常生理活動,能量代謝是細(xì)胞進(jìn)行其他生理活動的基礎(chǔ)。
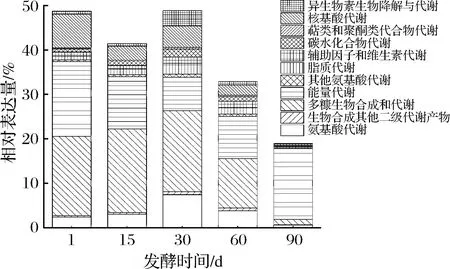
圖5 工業(yè)豇豆泡菜發(fā)酵過程中代謝途徑的基因表達(dá)富集分析Fig.5 KEGG analysis of gene expression of metabolism during the industrial cowpea paocai fermentation
3 結(jié)論
本研究利用宏轉(zhuǎn)錄組學(xué)技術(shù)對工業(yè)豇豆泡菜活性微生物群落結(jié)構(gòu)進(jìn)行了解析,結(jié)果表明乳桿菌屬(Lactobacillus)和片球菌屬 (Pediococcus) 是發(fā)酵初期主要的活性菌屬,發(fā)酵中期(15~30 d)的優(yōu)勢活性菌屬是乳桿菌屬(Lactobacillus),隨著發(fā)酵進(jìn)行,細(xì)菌豐度逐漸下降,以Starmerella為主的真菌豐度逐漸增加,最終主導(dǎo)了發(fā)酵末期(60~90 d)。同時(shí)對其進(jìn)行了KEGG功能注釋,工業(yè)豇豆泡菜液中活性微生物主要參與代謝功能,集中在碳水化合物代謝和能量代謝,碳水化合物代謝相關(guān)基因表達(dá)量占比最高。本研究結(jié)果加深了對工業(yè)豇豆泡菜微生物群落組成和多樣性的認(rèn)識,發(fā)現(xiàn)了發(fā)酵過程中起關(guān)鍵作用的菌屬,為解析泡菜發(fā)酵過程中微生物代謝機(jī)理提供參考,后續(xù)可進(jìn)一步分析活菌群落在發(fā)酵過程中具體的代謝途徑來揭示其對工業(yè)豇豆泡菜風(fēng)味和品質(zhì)的貢獻(xiàn),以期為發(fā)酵蔬菜工業(yè)化發(fā)展提供借鑒。
