不同群體刀鱭形態差異研究
2020-06-09 06:21:50張賽賽陳博錦成智麗孫鵬飛姜欣彤董安然姜志強
淡水漁業 2020年3期
張賽賽,羅 珺,陳博錦,成智麗,孫鵬飛,姜欣彤,董安然,姜志強,王 偉
(1.大連海洋大學水產與生命學院,遼寧省北方魚類應用生物學與增養殖重點實驗室,遼寧大連 116023; 2.大連市現代農業生產發展服務中心,遼寧大連 116023; 3.大連圣亞旅游控股股份有限公司,遼寧大連 116023)
刀鱭 (Coilianasus),隸屬鯡形目(Clupeiformes)鳀科(Engraulidae)鱭屬(Coilia),俗稱刀魚、鳳尾魚等,曾用名湖鱭,我國分布廣泛,北起遼寧遼河,南至廣東沿海及其與海相通的河流、湖泊,國外分布于朝鮮沿海以及日本有明海。刀鱭在我國主要產于黃渤海和東海沿岸各通海江河,其中以長江下游產量最高[1],為產區重要經濟魚類。近幾十年來由于過度捕撈和水域生態環境變化等因素,刀鱭野生資源銳減。郭弘藝等[2]對刀鱭漁汛集中的江蘇靖江段進行6年捕撈情況調查,發現其資源量有明顯下降趨勢。因此,隨著野生資源的銳減,研究不同地域刀鱭的形態差異,可為分析其種內的形態特征及分化規律提供數據支持。
形態學遺傳多樣性常用來反映魚類在形態方面的差異和作為種質選擇的重要參考。形態度量分析是傳統的魚類分類和種群鑒別常用的方法,但是往往無法鑒別同一物種的不同地理群體[3]。而多變量形態測量法較傳統的形態測量法能更有效區分種內不同群體的形態差異[4]。謝仲桂[5]采用多變量形態量法探討了不同居群華鳊屬(Sinibrama)物種的有效性。有關紅鰭東方鲀(Takifugurubripes)[6]、中國鱚(Sillagosinica)[7]和大瀧六線魚(Hexagrammosotakii)[8]的研究也說明了多變量形態測量能有效區分不同群體魚類。國外也常用多變量分析法研究魚類種群形態差異。Maruo等[9]利用形態學分析的方法確定了可以區分6種鮨科魚類的形態性狀。Gorgonio等[10]研究了墨西哥野生鱒(OncorhynchusSalmonidae)不同群體之間的形態差異。程起群等[11]對刀鱭和湖鱭形態差異的研究認為兩者為同一物種。向文殿[12]對不同定居型湖鱭形態差異的研究揭示了其定居演化的順序。王冰[13]以長江安徽段三個刀鱭群體為對象的研究表明三者形態差異小,主要差異出現在尾柄長與吻長。王丹婷等[14]對不同水域刀鱭群體形態差異的研究表明黃海群體與長江群體有關聯。眾多研究表明多變量分析法能有效判別魚類不同群體的差異,但是遼寧地區分布的刀鱭群體的形態學研究還是盲點。本研究采集了遼河、大洋河、鴨綠江和太湖4個不同地理群體的刀鱭樣品,通過對其中3個地理群體(遼河群體樣品因完整性被破壞無法測量框架結構)進行傳統形態測量和框架數據測量,以聚類分析、判別分析、主成分分析三種多元分析法對其進行綜合比較,以期了解群體間的差異及太湖群體與遼寧群體間差異,分析其種內的形態特征及分化規律,旨在為刀鱭的遺傳性狀改良研究提供數據支持。
1 材料與方法
1.1 試驗材料
試驗用魚來自4個地理群體,其中遼寧地區3個群體分別來自于遼河(LH)、大洋河(DYH)和鴨綠江(YLJ),江蘇1個為太湖群體(TH),合計112尾(表1)。所有樣品均采自天然水域野生群體,各群體樣品均為捕撈獲得,在船上低溫保存,上岸后冰箱冷凍。采集樣品時依據臀鰭鰭條數小于90和耳石形態鑒定刀鱭種類[15],遼河刀鱭野生分布稀少僅捕撈到20尾,其它采樣點在體型完整漁獲物中抽取樣品,保持冷凍狀態運輸回實驗室。
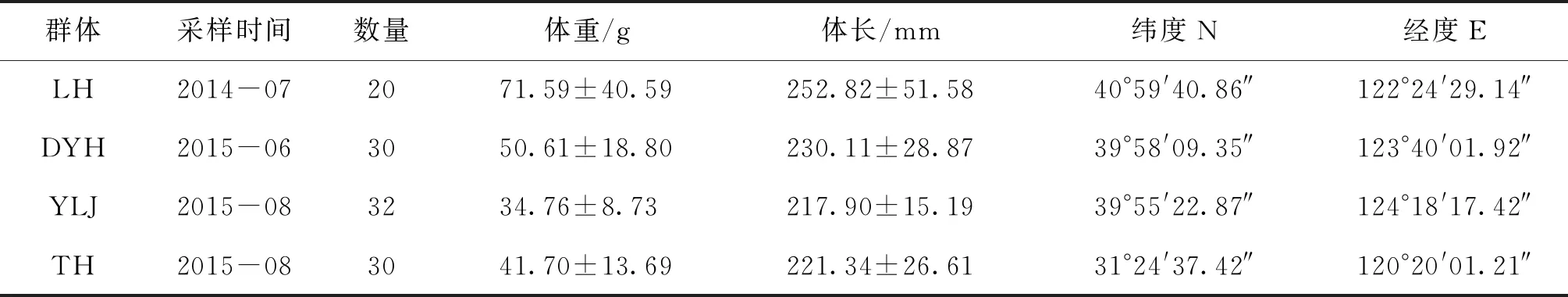
表1 四個刀鱭群體樣本的基本信息Tab.1 Basic information of C.nasus of the four populations
1.2 數據測量
魚體在低溫下緩慢解凍后平放在低溫托盤上,首先統計形態學的可數性狀,再進行可量性狀和框架結構參數的測量(遼河群體因完整性受破壞無法獲得部分可數性狀參數和框架結構參數)。所有可量性狀使用量魚板、兩腳規和電子數顯游標卡搭配使用進行測量,長度精確到0.01 mm,重量精確到0.01 g。
可數性狀,共11項:1.背鰭鰭條(DR),2.胸鰭鰭條(PTR),3.腹鰭鰭條(PVR),4.臀鰭鰭條(AR),5.尾鰭鰭條(TR),6.上鰓耙數(UR),7.下鰓耙數(LR),8.縱列鱗數(LS),9.橫列鱗數(RS),10.腹部棱鱗數(VS),11.脊椎骨數(NS)。
可量性狀,共18項:1.全長(TL),2.體長(BL),3.體高(BD),4.頭長(HL),5.吻長(SL),6.上頜骨長(JBL),7.眼徑(ED),8.眼間距(EI),9.眼后頭長(PL),10.尾柄高(CD),11.背鰭長(DL),12.胸鰭長(PTL:第一根鰭絲長),13.腹鰭長(PVL),14.臀鰭長(AL),15.尾鰭長(TFL),16.體厚(BT),17.體重(BW),18.凈體重(LBW)。
參考現有的框架結構[11-13]構建方式,刀鱭體側上下肌肉間的水平隔膜肉眼可見一條線(D13),和其它坐標點具有同源性,它是刀鱭生理結構的一部分。在水平隔膜線上分別選取了3個點:與D3-6的交點11,與D7-8的交點12,還有D13的1/2處點14,體兩側分別對應,點11′、點12′和點14′,增加三個反映魚體不同位置厚度的參數(D11-11′、D12-12′和D14-14′),建立刀鱭改進框架結構。魚體周身共16個坐標點,25個框架數據。框架結構圖(圖1)如下:

圖1 刀鱭的框架圖Fig.1 Truss network of C.nasus
1.下頜骨末端,2.吻前端,3.胸鰭基部前緣,4.頭背部末端,5.腹鰭基部前緣,6.背鰭基部前緣,7.臀鰭基部前緣,8.背鰭基部后緣,9.尾鰭基部后端,10.尾鰭背部起點,11.D3-6和體中軸線交點;11′.魚體右側D3-6和體中軸線交點,12.D7-8和體中軸線交點;12′.魚體右側D7-8和體中軸線交點,13.肌肉水平隔膜長,14.水平隔膜1/2處點。D1-2表示坐標點1到坐標點2之間的距離。
1.3 統計分析
由于刀鱭成熟雌雄魚個體無明顯第二性征,形態上基本無差異[14],雌雄個體統一測量分析。為校正因樣品規格差異和個體差異對形態參數造成的影響,將長度參數除以體長(BL)進行統一量綱校正。所有的數據采用Microsoft Excel 2010軟件工具進行統計,應用統計學軟件SPSS 20.0對可數性狀參數進行單因子方差分析,實驗數據以平均值±標準差表示,對40個可量性狀參數進行主成分分析,判別分析和聚類分析,其中P<0.05表示差異顯著。
對4個地理群體刀鱭的可數性狀特征進行單因子方差分析。對40個形態參數做主成分分析。分別求出各群體40個參數的比例參數的均值,再用獲得的各項參數均值作聚類分析。對3個群體刀鱭形態差異貢獻較大的參數建立判別函數公式。然后進行交互驗證,檢驗判別公式可靠性。
2 結果
2.1 四個群體可數性狀方差分析
由表2可知,腹鰭條數和臀鰭條數在4個不同刀鱭群體中差異不顯著,可見刀鱭臀鰭條數較穩定。縱列鱗和脊椎骨數太湖群體顯著小于遼寧組群(LH、DYH、YLJ),上下鰓耙數和腹部棱鱗太湖群體顯著大于遼寧組群(DYH、YLJ),背鰭條數、胸鰭條數、尾鰭條數、橫列鱗數沒有出現明顯的南北方組群差異。分析大洋河、鴨綠江和太湖3個刀鱭群體肥滿度(condition factor)結果,大洋河群體最大,鴨綠江群體最小。
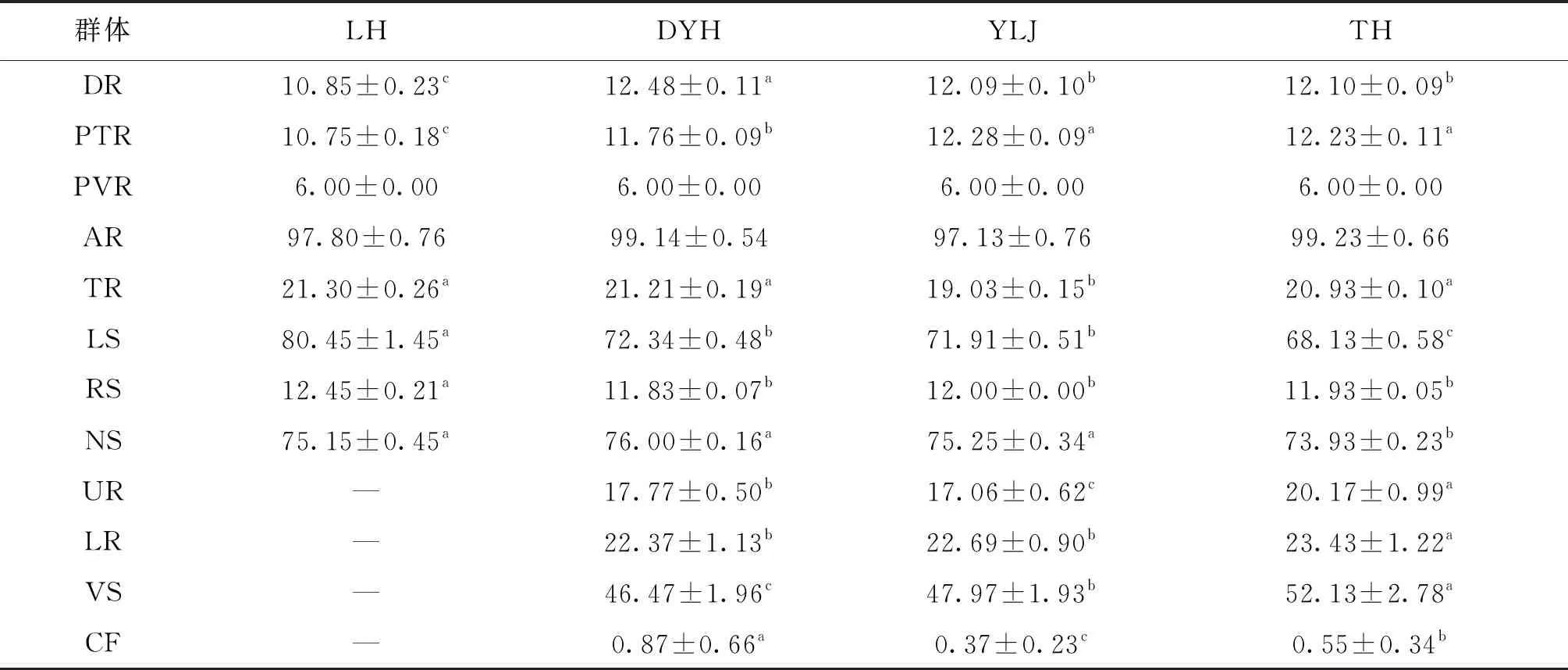
表2 四個刀鱭群體可數性狀參數方差分析Tab.2 Variance analysis of the countable character parameters of four C.nasus
注:同行上標不同小寫字母表示差異顯著(P<0.05)。下同。
2.2 三個群體刀鱭的主成分分析
由表3主成分分析結果可知,6個主成分對不同群體間總的變異的累積貢獻率積可達92.192%,超過了85%的要求,說明用幾個相互獨立的因子可以基本概括不同群體刀鱭的形態特征。進一步分析前兩個貢獻率較大的主成分發現,第一主成分主要歸納了D7-8/BL和D7-10/BL這2個形態參數特征,反映的是魚體縱向的特征;第二主成分主要歸納了TL/BL、D7-9/BL、D8-9/BL、D8-10/BL和D13/BL共5個形態參數的特征。由此可見刀鱭形態學差異主要由魚體縱向背鰭和尾鰭之間的差異造成的。
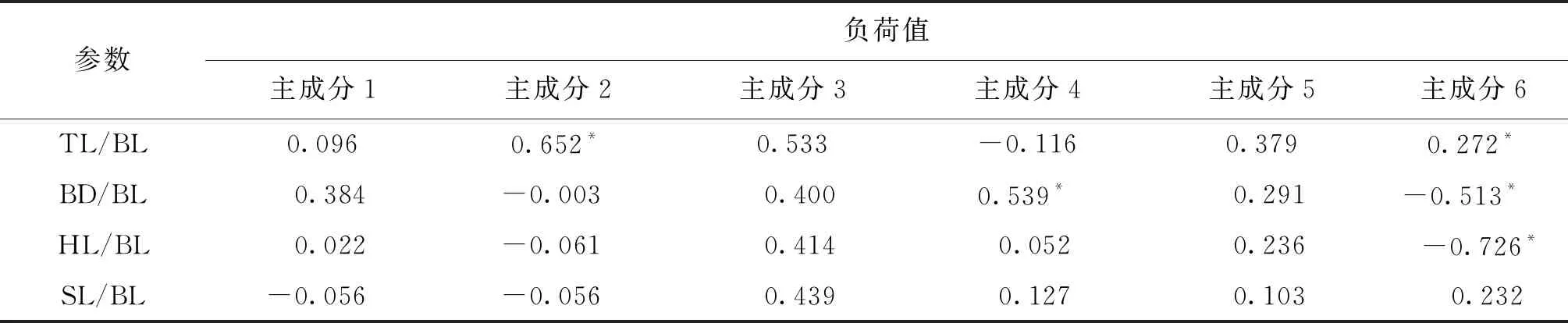
表3 3個群體刀鱭的40個形態參數對6個主成分的特征向量及主成分的貢獻率Tab.3 Eigenvectors and cumulative contribution rates of six principal components from 40 traits for three populations of C.nasus
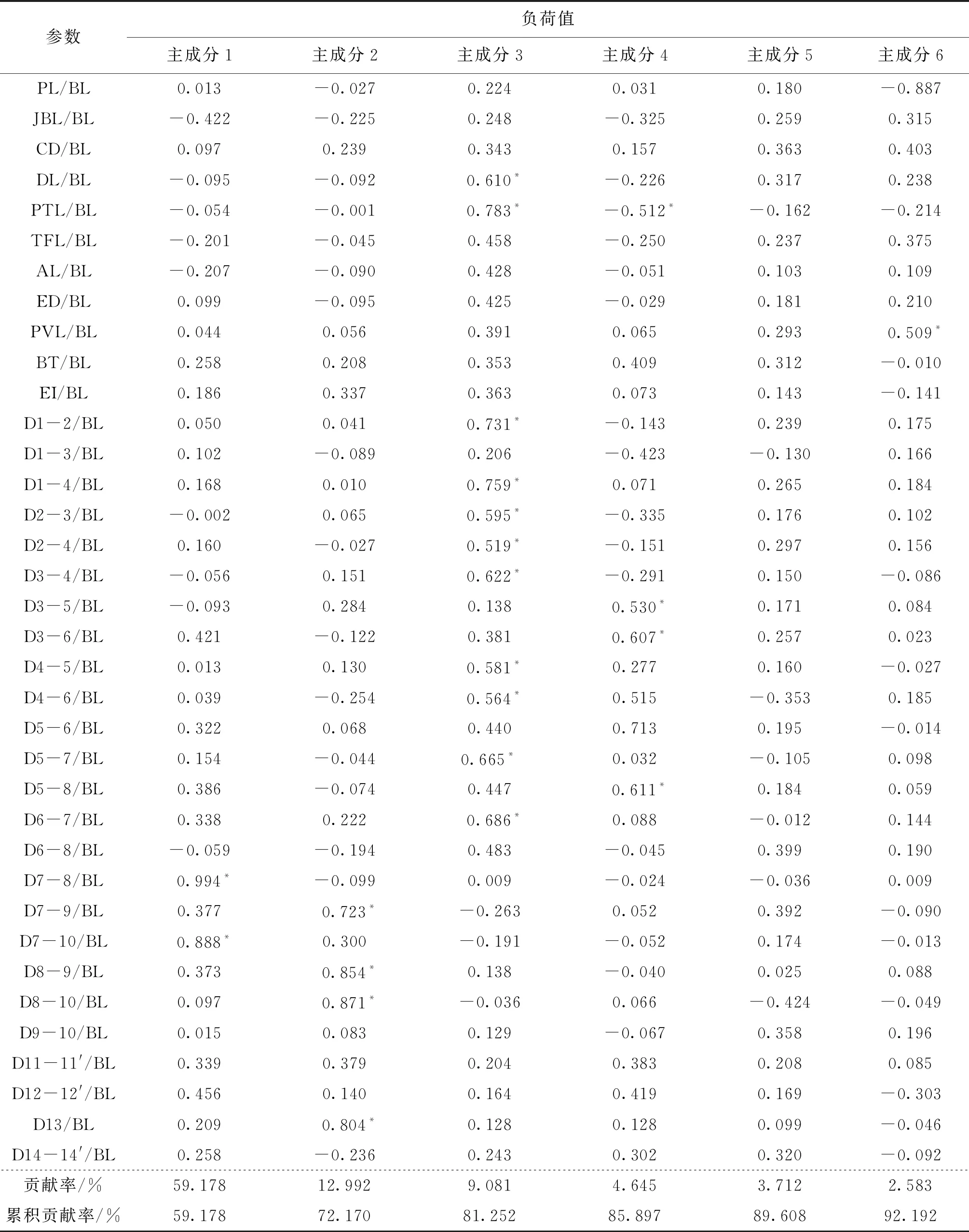
續表3
注:標*表示主要貢獻的參數(負荷值>0.500)
由圖2 刀鱭群體主成分1和主成分2的散點圖可知,分群體刀鱭在主成分1和主成分2兩方面基本可以分開,大洋河與鴨綠江和太湖群體分離較遠,鴨綠江和太湖群體有很小程度的重疊,3個群體基本可以分離開。
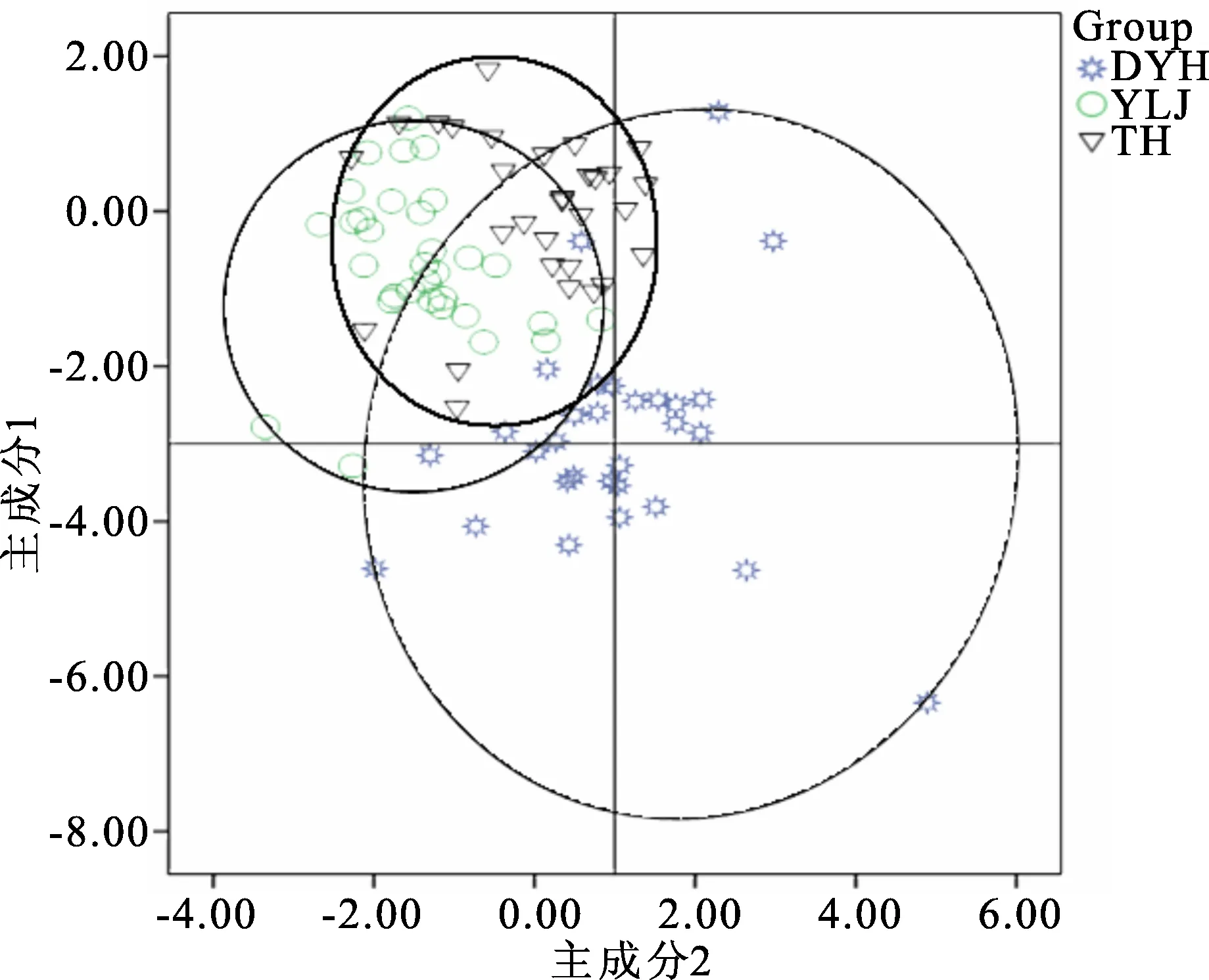
圖2 3個群體刀鱭主成分1和主成分2散點圖Fig.2 Scatter diagram for PC1 and PC2 of three populations of C.nasus
2.3 三個群體刀鱭的聚類分析
對三個群體40項形態參數取平均值通過平均連接法進行系統聚類分析。見圖3可知,3個群體聚成2支,大洋河單獨一支,鴨綠江和太湖聚成一支。

圖3 3個群體刀鱭聚類分析圖Fig.3 Cluster dengdrogram of three populations of C.nasus
2.4 三個群體刀鱭的判別分析
利用SPSS 20.0逐步判別法,對40個形態參數進行分析,選取ED/BL、JBL/BL、PTL/BL、EI/BL、D1-3/BL、D2-4/BL、D5-7/BL、D7-9/BL、D14-14′/BL共9個顯著貢獻的參數。用9個參數構建了3個群體的判別函數:
DYH=11 235.271×ED/BL+3 407.122×JBL/BL-101.743×PTL/BL+1 243.267×EI/BL+4 204.587×D1-3/BL-4 739.015×D2-4/BL+2 215.453×D5-7/BL+5 881.006×D7-9/BL+1 733.715×D14-14′/BL-2 306.080
YLJ=11 750.268×ED/BL+3 690.739×JBL/BL-23.868×PTL/BL+628.154×EI/BL+4 518.769×D1-3/BL-5 304.641×D2-4/BL+2 384.934×D5-7/BL+5 943.482×D7-9/BL-93.481×D14-14′/BL-2346.350
TH=15 952.276×ED/BL+2 907.263×JBL/BL+1.813×PTL/BL-4.881×EI/BL+3 986.397×D1-3/BL-4 685.127×D2-4/BL+2 597.654×D5-7/BL+5 546.581×D7-9/BL+609.062×D14-14′/BL-2178.959
為驗證上述三個判別函數的實用性,對調查的3個群體刀鱭樣本采集的數據集使用判別函數進行驗證,判別結果見表4。由表可見,群體內的自身驗證判別準確率均為100.0%,群體間的交互判別率為93.8%~100.0%,綜合判別率96.74%,交互判別結果和群體內判別結果基本一致,由此可知3個群體的判別函數是可靠的,可以在實際鑒別刀鱭各群體時使用。
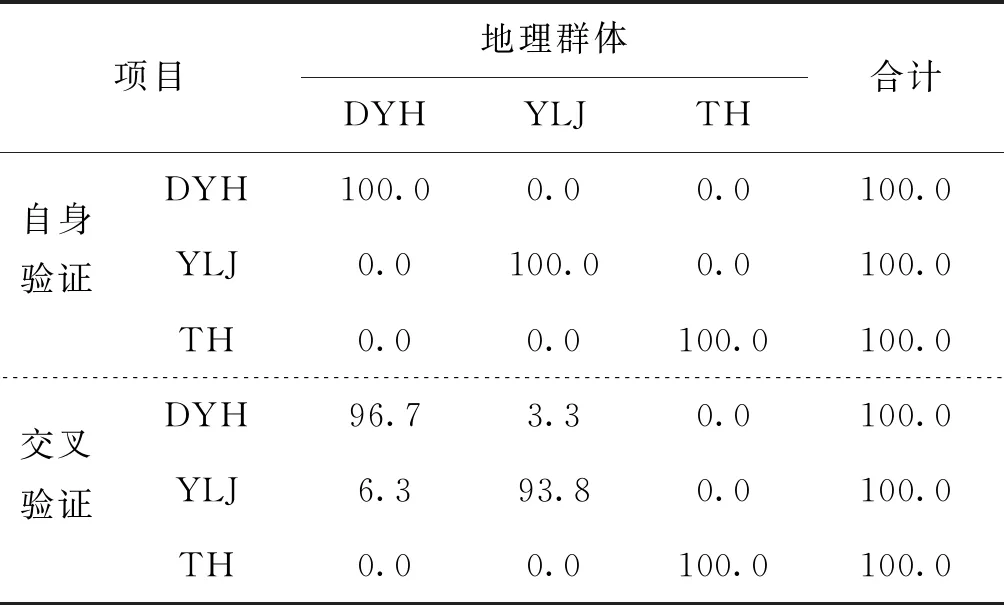
表4 3個群體刀鱭的判別結果Tab.4 The discriminant results of three populations of C.nasus %
2.5 主成分分析和判別分析篩選出的參數
主成分分析法第一主成分、第二主成分中貢獻率較大參數有7個,選出6個框架結構中參數,表5列出參數統一量綱后的數值。D7-9/BL、D7-10/BL、D8-9/BL、D13/BL、D8-10/BL五個參數遼寧兩群體顯著大于太湖群體,D7-8/BL參數遼寧群體顯著小于太湖群體。
判別分析法中軟件在40個參數中選取9個有顯著貢獻的參數用于構建判別函數。表6列出9個
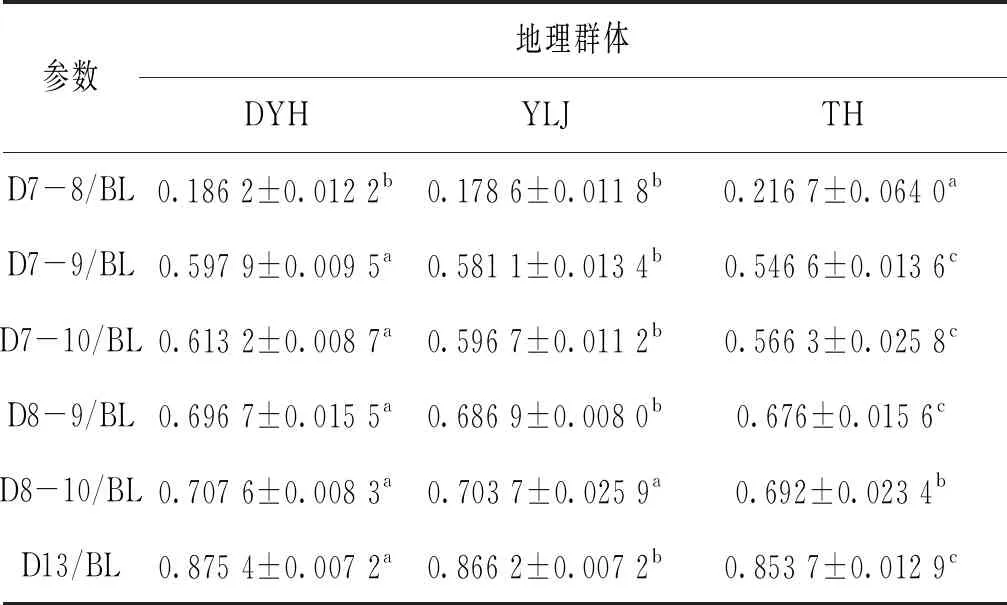
表5 主成分分析貢獻率較大參數Tab.5 The contribution rate of principal component analysis is large
參數的統一量綱后的數值。ED/BL、JBL/BL、EI/BL、D1-3/BL、D2-4/BL參數集中反映魚體頭部差異,D7-9/BL、D14-14′/BL反映尾部差異,D5-7/BL反映腹部差異。
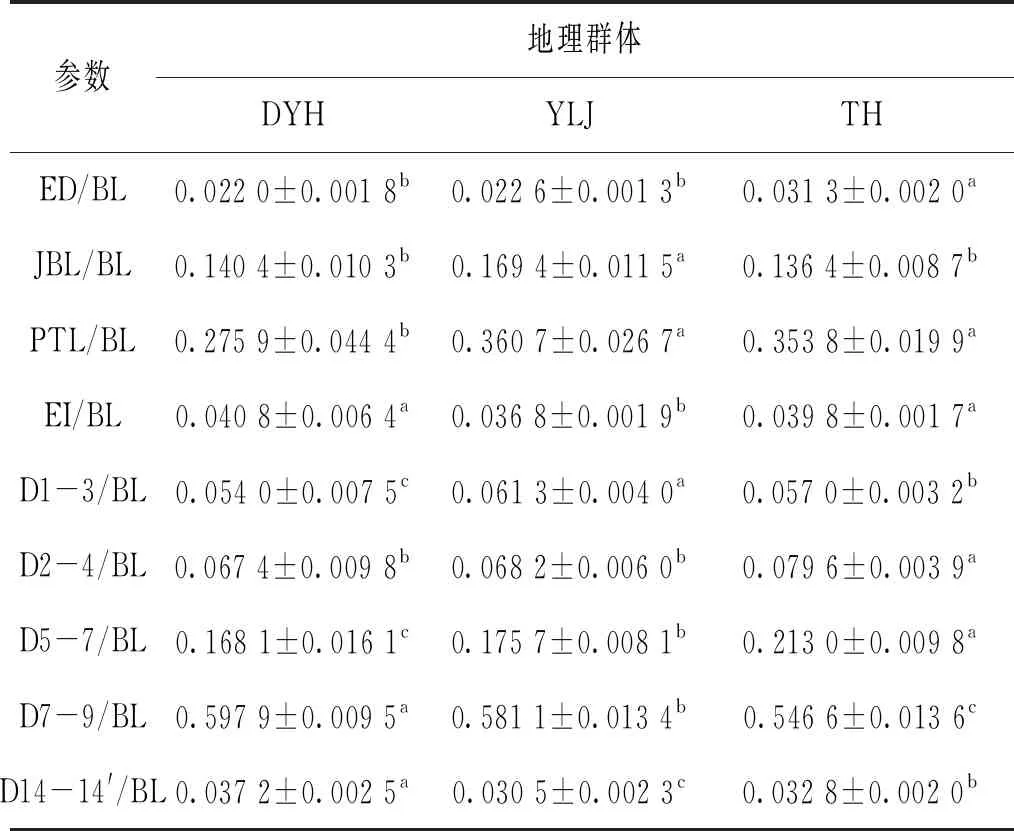
表6 判別分析貢獻大參數Tab.6 Discriminant analysis contributes large parameters
3 討論
3.1 不同群體刀鱭形態差異
20世紀90年代開始,幾何形態測量法在魚類形態學研究中逐漸得到推廣[16]。刀鱭是游泳速度較快的魚類,在游動時尾部快速擺動呈S型,尾部擺動產生的推力提供刀鱭快速的游動。大洋河群體和鴨綠江群體刀鱭為洄游生態型會做長距離洄游,太湖刀鱭為內陸生態型只在淡水生活不做長距離洄游,結合表5信息猜測長距離洄游的生態因素引起刀鱭D7-8較體長比值變小,D7-9、D7-10、D8-9、D13、D8-10較體長比值變大。王冰[13]對長江安徽段3個不同刀鱭群體形態差異在尾柄長/體長結果一致。王偉等[17]對游泳速度快的翹嘴鲌的7個群體研究發現,各群體的頭部和尾部發生了變異,刀鱭與翹嘴鲌都屬于側扁體型,尾部變異觀點與本研究一致。曾聰[18]選取不同湖泊的團頭魴研究發現尾部也出現了變異。類似的研究結果在Cadrin等[19]對大西洋黃蓋鰈(Limandaferruginea)的形態差異分析中也見到。因生態環境不同是造成刀鱭尾部形態差異分化的主要因素。判別分析選出參數主要集中在刀鱭頭部,由表6顯示上頜骨長、眼徑、眼間距、D1-3/BL和D2-4/BL存在顯著差異,但無明顯南北方差異,猜測可能與攝食行為和水環境有關。刀鱭在游動時張開上下頜通過鰓耙過濾水中的枝角類、橈足類和小蝦等,上下頜張開的狀態長期受到游動速度和水生態環境的影響,這可能影響刀鱭頭部比例的變化。這個猜測與張秀霞等[20]有關唐魚(Tanichthysalbonubes)不同群體頭部形態變異的研究結果一致。王冰[13]的研究也表明長江安徽段不同刀鱭群體“吻長/體長”有差異。其它研究也發現魚類生態形態的關聯,眼徑大小與食物在水中的位置相關[21]。由此可見刀鱭頭部和尾部易受生態環境影響發生差異分化。聚類分析較直觀的顯示出群體間的關系遠近,分析結果把鴨綠江和太湖兩群體聚為一支,大洋河群體單獨一支,結果和主成分分析接近。這與王丹婷等[14]的研究刀鱭黃海群體與長江群體有關聯的結果一致。判別分析中交叉驗證時出現3個體判別失誤,鴨綠江群體2個體判別為大洋河群體,大洋河群體1個體判別為鴨綠江群體,兩群體間存在個別體型接近情況。
3.2 關于改進框架測量法的探討
現有魚類形態框架測量法構建的網狀框架只覆蓋了魚體側面,缺失了魚體厚度方面的信息,傳統形態中魚體厚參數單一而且與框架法的網狀結構無法構成一個整體結構。本研究中在構建刀鱭的框架結構時做了改進,增加了魚體厚度方面的參數和水平隔膜長度(D13),使之與框架結構形成一體,成為三維檢測構架。改進后的刀鱭框架結構由全身16個坐標點形成25個框架參數。在主成分分析中,D13對主成分2的負荷值為0.804>0.500 具有顯著的貢獻。根據SPSS 20.0逐步判別法,由40個形態參數篩選出的9個有顯著貢獻的參數,D14-14′在其中,說明增加的參數對判別起到了貢獻作用。表4結果表明各群體刀鱭自身驗證準確率100%,綜合判別準確率高達96.74%。明顯高于王丹婷等[14]對不同水域刀鱭形態研究中,自身驗證準確率93.10%和綜合判別準確率71.55%;向文殿等[22]不同湖泊刀鱭形態研究中綜合判別準確率80.6%;王偉等[17]多個翹嘴紅鲌群體形態差異綜合判別率為90%。由此可知,增加D13 和D14-14′參數對分析魚類形態差異有很大的幫助,能更可靠穩定地判別不同群體魚類。在框架測量法中增加不同位置魚體體厚的數據,建立更全面反映魚體體型的框架,這是對現有框架測量法的改進,值得在其它魚類的形態學研究中進一步研究和探討。
猜你喜歡
結構工程師(2022年2期)2022-07-15 02:23:50
音樂探索(2022年2期)2022-05-30 21:01:37
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
建材發展導向(2021年14期)2021-08-23 00:56:34
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
工程建設與設計(2016年2期)2016-08-22 09:12:46
