新疆傳統乳品中產胞外多糖乳酸菌的篩選及益生特性的研究
2020-06-08 07:55:36高云云李寶坤盧士玲蔣彩虹王慶玲
中國釀造 2020年5期
高云云,李寶坤,盧士玲,蔣彩虹,王慶玲,董 娟
(石河子大學食品學院,新疆石河子 832000)
乳酸菌(lactic acid bacteria,LAB)是指發酵碳水化合物產生乳酸的一類細菌的總稱,廣泛存在于自然界中,一般被認為是安全的(generally regards as safe,GRAS)[1]。乳酸菌胞外多糖(ex-polysaccharides,EPS)是微生物在生長代謝過程中分泌到細胞外的粘液或黏附在細胞表面的莢膜多糖[2],乳酸菌胞外多糖已被證明具有增強免疫調節[3]、抗腫瘤[4]、降膽固醇[5]、抗氧化[6]、改變發酵乳流變學特性[7]等作用。目前對與乳酸菌胞外多糖各種性質的研究方法較為成熟,所以將其應用于乳制品中來改變發酵乳品質和制作功能性乳品方面有廣闊的前景。
乳酸菌作為益生菌應具備多種對人體健康有益的生理功能,包括抑制有害菌的生長、延緩機體衰老、降低膽固醇、緩解乳糖不耐癥、調節腸道菌群平衡等[8]。乳酸菌發揮益生功效的前提是必須克服宿主體內的各種不利條件,能夠耐受胃液的低pH和腸液環境中的高濃度的膽鹽及具有附著在宿主腸壁細胞的能力等,這是乳酸菌發揮作用尤為重要的條件[9-10]。近年來,乳酸菌胞外多糖由于良好的功能特在食品和醫藥行業中廣泛應用,這使得乳酸菌受到更多的關注[11],乳酸菌胞外多糖作為增稠劑應用在酸奶生產中能夠起到改善酸奶質地、提高產品穩定性增強口感等作用[12]然而,國內對于乳酸菌EPS的研究起步較晚,但近年來已取得很大的進展[13]。關于如何準確篩選產EPS菌株和提高產量是目前國內學者研究的熱點。
新疆牧區少數民族在制作傳統乳品方面有著豐富的經驗,這為新疆傳統乳制品中優良乳酸菌菌種的開發利用提供了豐富的資源。本研究分析新疆傳統乳品中微生物多樣性,并篩選出了3株高產胞外多糖的植物乳桿菌,對菌株的潛在益生特性進行分析,以期為進一步利用此種菌株開發功能性食品提供良好的科學依據。
1 材料與方法
1.1 材料與試劑
1.1.1 原料和菌株
10份樣品均來自塔地區不同牧場。其中酸奶來源于二支河牧場四隊2份、二道橋鄉1份;酸馬奶來自霍吉爾特鄉和二道橋鄉2份;鮮馬奶來源于喀拉也木勒鄉1份、二道橋固牧場2份。樣品采集后裝入50 mL無菌離心管內旋緊蓋并做標記。將采集到的樣品至于低溫采樣箱中迅速運回實驗室,放置在-4 ℃冰箱中,為防止微生物損失,盡快對其中的微生物進行分離。
大腸桿菌(Escherichia coli)、金黃色葡萄球菌(Staphylococcus aureus):石河子大學畜產品加工實驗室;鼠李糖乳桿菌(Lactobacillus rhamnosus):衡陽師范學院。
1.1.2 化學試劑
苯酚、濃硫酸、無水乙醇、三氯化鐵、牛膽鹽、膽固醇、葡萄糖、鐵氰化鉀、磷酸等均為國產分析純:國藥集團化學試劑有限公司;細菌基因組脫氧核糖核酸(deoxyribonucleic acid,DNA)提取試劑盒:北京天根生化科技限公司;1,1-二苯基-2-三硝基苯肼(1,1-diphenyl-2-trinitrophenylhydrazine,DPPH)(分析純):美國Sigma公司。
1.1.3 培養基
脫脂乳培養基(EMS)[14]:90 g/L脫脂乳,3.5 g/L酵母提取物,3.5 g/L蛋白胨,10 g/L葡萄糖。MRS肉湯培養基、MRS固體培養基、BHI培養基、Mueller Hinton Agar(MHA)培養基[15]:青島高科技工業園海博生物技術有限公司。
1.2 儀器與設備
DNP-9272電熱恒溫培養箱:上海精宏實驗設備公司;T100TTMhermal Cycler聚合酶鏈式反應(polymerase chain reaction,PCR)擴增儀:美國BIO-RAD公司;Universal Hood II凝膠成像儀:美國BIO-RAD公國藥集團化學試劑有限公司司;Nedfuge 15R臺式高速冷凍離心機:賽默飛世爾科技(中國)有限公司;EONC酶標儀:美國伯騰儀器有限公司;LDZX-30KBS立式壓力蒸汽滅菌鍋:上海申安醫療器械廠。
1.3 實驗方法
1.3.1 乳酸菌的分離純化
取酸奶、酸馬奶、鮮馬奶樣品各1 mL,用無菌生理鹽水10倍稀釋至10-5、10-6、10-7。取各濃度稀釋液100 μL涂布于MRS固體培養基上,37 ℃培養36 h后根據菌株在固體平板上的生長情況,挑選菌落形態不同的菌株進行革蘭氏染色及過氧化氫酶實驗。選擇革蘭氏染色陽性過氧化氫酶陰性結果的菌株進一步在MRS培養基中純化培養后,取適量菌液接入終濃度為30%甘油中,-80 ℃保藏。
1.3.2 菌株的生物學鑒定
使用試劑盒提取菌株DNA。采用通用引物27f(5'-AGAGTTTGATCCTGGCTCAG-3')和1492r:(5'-CTACGGCTACCTTGTTACGA-3'),PCR擴增程序參照文獻[16]。PCR產物經1.0%瓊脂糖凝膠電泳檢測后,送往上海生工生物科技有限公司進行測序,測序結果提交至GenBank數據庫中進行序列同源性比對,并用MAGE 5.0軟件構建系統發育樹。
1.3.3 產胞外多糖乳酸菌的篩選
用無菌接種環蘸取一定菌液四區劃線于MRS固體培養基上。另取200 μL接種于ESM培養基中。置于37 ℃培養箱中培養24 h后。觀察MRS瓊脂培養基上菌落形態和EMS培養基中的拉絲情況并做適當記錄。
1.3.4 胞外多糖的提取及含量測定
參考LIU A等[17]的方法并做適當修改,將活化好的菌株接3%(V/V)于MRS肉湯中,37 ℃培養24 h得到發酵液,發酵液于4 ℃條件下8 000 r/min 離心20 min后收集上清液。加入3倍體積的冰乙醇4 ℃過夜放置。4 ℃條件下8 000 r/min離心20 min后,將沉淀物溶解在一定體積的去離子水中,加入終濃度為8%的三氯乙酸在4 ℃保持過夜去除蛋白質。再次離心除去沉淀,上清液置于4 ℃純水中透析48 h,每8 h換一次水。
多糖含量的測定采用苯酚-硫酸法,參照王輯等[8]的方法配制不同濃度的葡萄糖標準溶液,測定各個濃度的葡萄糖溶液在波長490 nm的吸光度值,分別以葡萄糖溶液濃度(x)和吸光度值(y)為橫坐標和縱坐標繪制葡萄糖標準曲線,得標準曲線回歸方程y=0.008x+0.004,相關系數R2=0.998 7,按照回歸方程計算樣品中胞外多糖的含量。
1.3.5 產糖菌株的耐受性
(1)耐酸能力評價
調節MRS培養基的pH值為3,接種3%(V/V)活化好的乳酸菌,37 ℃培養4 h后將所得菌液適當稀釋進行平板菌落計數[18]。
(2)膽鹽耐受性
參照DELGADO S等[19-20]的方法做部分調整。將活化好的菌液接種3%(V/V)于0、0.1%、0.2%、0.3%牛膽鹽的MRS培養基中,37 ℃培養,每小時測吸光度值(OD620nm值),直至OD620nm值上升0.3個單位,確定乳酸菌在不同濃度牛膽鹽中的生長滯后期,其計算公式如下:
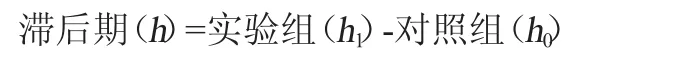
式中:h1為實驗組OD620nm值上升0.3個單位需要的時間;h0為對照組OD620nm值上升0.3個單位需要的時間。
(3)人工胃液耐受性
將活化好的菌液4 000 r/min離心10 min,吸取上清液,用磷酸鹽緩沖液(phosphate buffered saline,PBS)洗滌沉淀兩次后重懸于等體積生理鹽水中混勻制得菌液,采用稀釋涂布平板法計算此時的菌液濃度(CFU/mL)。取1 mL菌懸液于9 mL pH值為3的模擬人工胃液中(MRS肉湯中按3 g/L添加胃蛋白酶),37 ℃厭氧培養3 h后進行平板菌落計數[21],菌株存活率計算公式如下:
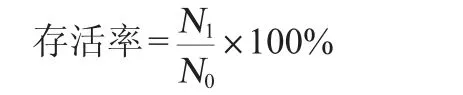
式中:N1為3h后的菌落數,CFU/mL;N0為0h的活菌數,CFU/mL。
(4)人工腸液耐受性
取1 mL菌懸液于9 mL模擬腸液中(在MRS肉湯里添加1 g/L胰蛋白酶和3 g/L牛膽鹽),37 ℃培養厭氧培養3 h后進行平板菌落計數。
1.3.6 產糖菌株DPPH自由基清除率測定
取入2 mL菌液于1 mL 0.2 mmol/L DPPH中,用無水乙醇補足2 mL,立刻渦旋搖勻,避光反應30 min,測定混合液在波長517nm處的吸光度值(OD517nm值)。以未加樣品為對照,無水乙醇作為空白[22]。DPPH自由基清除率計算公式如下:
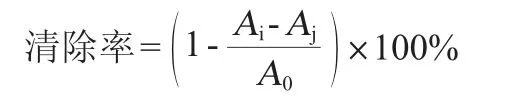
式中:Ai為1 mL的DPPH+1 mL樣品的吸光度值;Aj為1 mL溶劑+1 mL樣品的吸光度值;A0為1 mL DPPH+1 mL溶劑的吸光度值。
1.3.7 產糖菌株自凝聚率的測定
參考SON S H等[23]的方法作適當修改:將活化好菌液在4 000 r/min離心20 min,用PBS洗滌細胞兩次并重懸于相同的緩沖液中,調節OD600nm值=0.3±0.05。將細菌細胞懸浮液37 ℃孵育2 h,取樣測波長600 nm處吸光度值,自凝聚率計算公式如下:
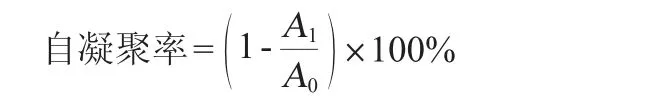
式中:A0為0 h的吸光度值;A1為2 h的吸光度值。
1.3.8 產糖菌株表面疏水性的測定
調節菌液的初始OD600nm值=0.5±0.02。將3 mL菌懸液和1 mL二甲苯于室溫條件下混勻混合。將混合物37 ℃靜置20 min后取水相,測定其在波長600 nm處的吸光度值(OD600nm值),表面疏水性計算公式如下:
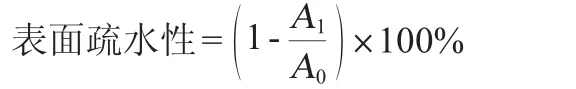
式中:A0為0 h的吸光度值;A1為加入二甲苯后的吸光度值。
1.3.9 產糖菌株膽固醇去除能力
參考吳云[24]的方法做適當修改。將活化好的菌液接3%(V/V)于膽鹽含量為0、0.1%、0.2%、0.3%且膽固醇終體積為100 μg/mL的MRS液體培養基中,搖床培養24 h后,取1 mL發酵液加入4 mL無水乙醇混合均勻靜置20 min,4 ℃、5 000 r/min離心10 min,取2.5 mL 上清液加入2.5 mL磷鐵硫試劑用漩渦混合器混勻冷卻后,測定波長560 nm處的吸光度值(OD560nm值),分別以膽固醇質量濃度(x)和吸光度值(y)為橫坐標和縱坐標繪制膽固醇標準曲線。得標準曲線回歸方程y=0.007 1x-0.009 4(相關系數R2=0.995 5),發酵液的膽固醇含量和菌株的膽固醇去除率計算公式如下:
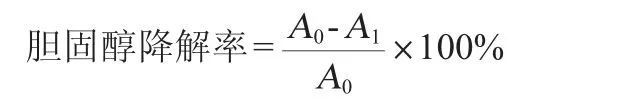
式中:A0為培養前膽固醇含量,mg/mL;A1為培養后膽固醇含量,mg/mL。
1.3.10 產糖菌株抑菌能力評價
將-80 ℃保藏金黃色葡萄球菌、大腸桿菌,在BHI液體培養基中培養24 h,Muelller-Hinton瓊脂培養基純化后,使其達到對數期備用。
采用牛津杯法測定乳酸菌的抑菌活性,指示菌使用Muelller-Hinton瓊脂培養基調整至106~107CFU/mL倒平板待其凝固后,將已滅菌的牛津杯放于瓊脂表面上。取200 μL乳酸菌菌液于牛津杯中,37 ℃培養24 h,觀察并測量抑菌圈直徑。通過探究菌株對大腸桿菌、金黃色葡萄球菌的抑制作用所形成的抑菌圈直徑大小來評估益生菌對體內致病菌感染的預防和抑制能力。
1.3.11 數據處理
使用Origin 2016進行圖表的制作;利用SPSS25.0進行差異性分析;實驗數據均進行3次重復,以平均值±標準偏差表示。
2 結果與分析
2.1 乳酸菌的分離及鑒定
從樣品中共分離得到153株純培養物,經過革蘭氏染色和過氧化氫酶實驗初步鑒定147株為疑似乳酸菌。通過16S rDNA基因檢測分析,確定145株乳酸菌。部分乳酸菌系統發育進化關系如圖1所示。

圖1 部分乳酸菌的系統發育樹Fig.1 Phylogenetic tree of some lactic acid bacteria
其中優勢菌種為屎腸球菌(Enterococcus)41株約占27.8%,發酵乳桿菌(Lactobacillus fermentum)36株約占24.5%,植物乳桿菌(Lactobacillusplantarum)23株約占15.6%,其余分別為短乳桿菌(Lactobacillus brevis)10株,戊糖片球菌(Pediococcus pentosaceus)9株,保加利亞乳桿菌(Lactobacillus bulgaricus)8株,食竇魏斯氏菌(Weissella cibaria)6株,瑞士乳桿菌(Lactobacillus helveticus)4株,副干酪乳桿菌(Lactobacillus paracasei)4株,耐久腸球菌(Enterococcus durans)3株,羅伊氏乳桿菌(Lactobacillus reuteri)1株。
2.2 產胞外多糖乳酸菌的篩選
由圖2可知,在優勢菌種屎腸球菌、發酵乳桿菌和植物乳桿菌中初篩出22株在MRS固體培養基和EMS培養基中有明顯產粘拉絲情況的菌株。由表1可知,用苯酚硫酸法對其所產多糖進行定量,實驗結果顯示出3株產胞外多糖含量較高的菌株1-3、1-6、4-1,均為植物乳桿菌。

圖2 產胞外多糖菌株的菌落形態Fig.2 Colony morphology of exopolysaccharides-producing strains
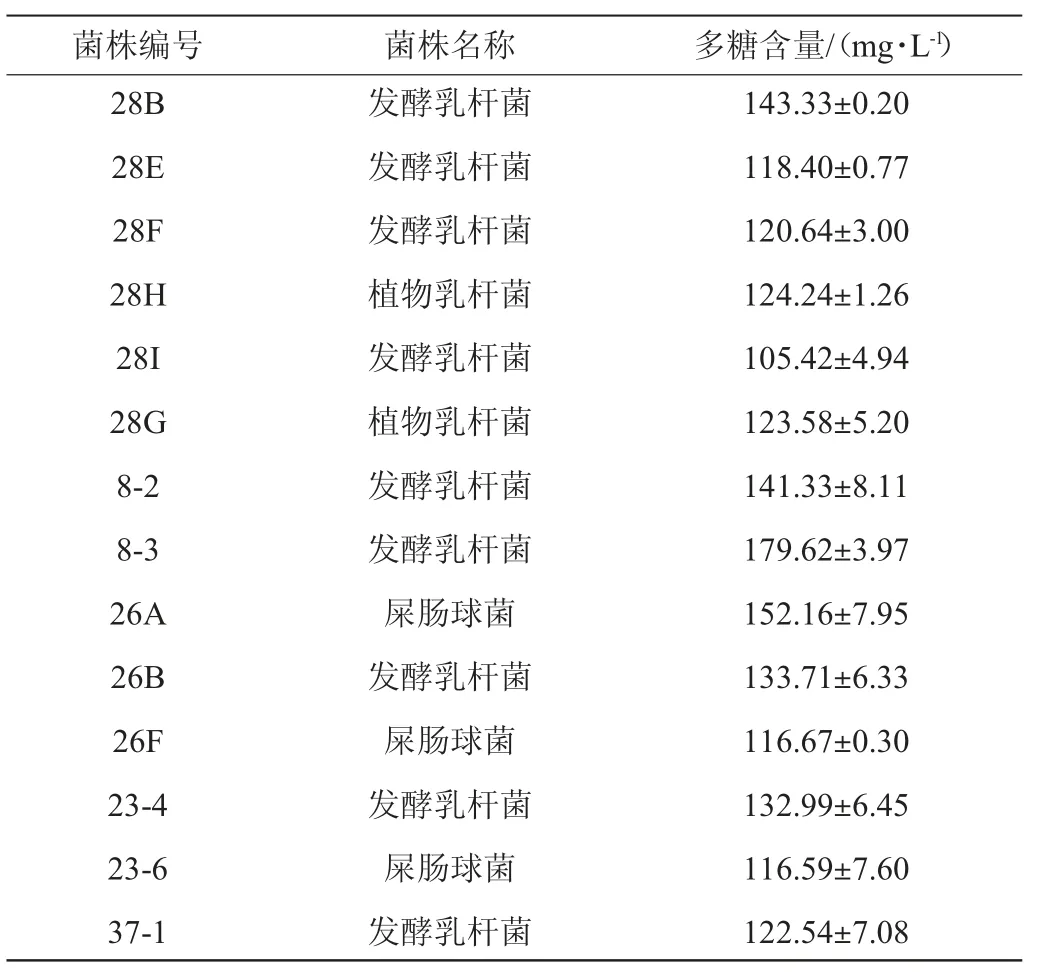
表1 菌株產胞外多糖的產量Table 1 Yield of exopolysaccharides produced by strains
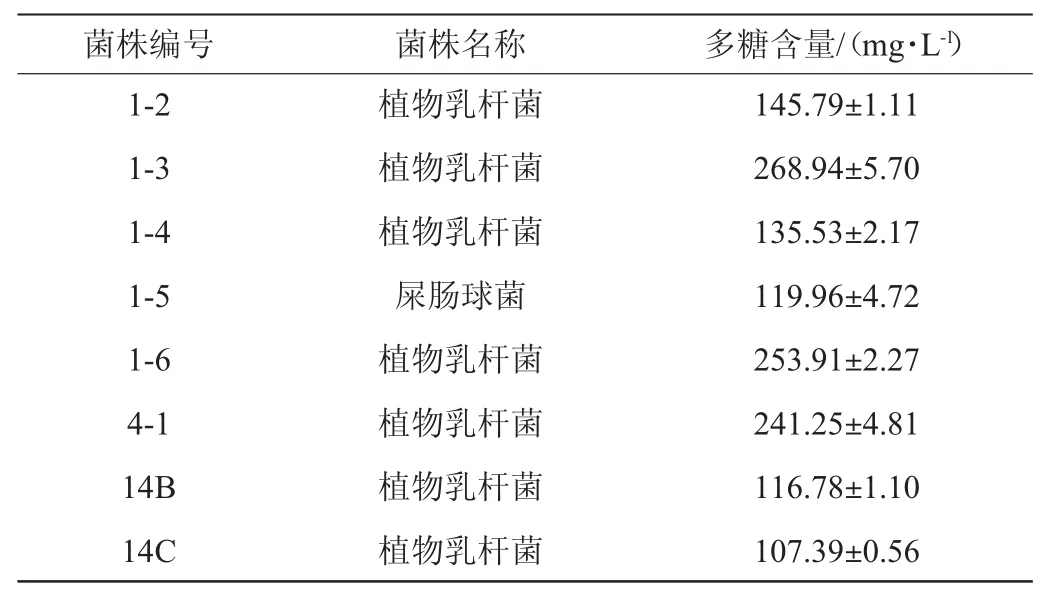
續表
2.3 產EPS植物乳桿菌的耐受性
2.3.1 耐酸能力評價
益生乳酸菌在pH值為3的環境中消化3 h后菌株的存活是作為評價菌株耐酸性的標準[25]。本實驗中用菌株在pH值為3的環境中消化4 h來進一步保證其在3 h的活菌數。如圖3所示,3株植物乳桿菌對低pH的生長環境都有一定的耐受能力,在pH值為3的條件下處理4 h后,菌株4-1活菌數最多顯著高于鼠李糖乳桿菌(P<0.05),但菌株1-3和1-6也表現出一定的耐酸能力,預示該菌株進入消化道后具有對胃液中低pH的不利條件有較強的抵抗能力。
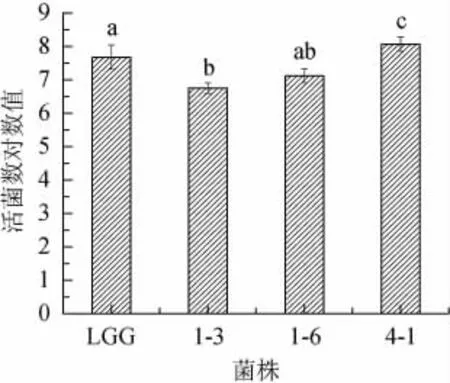
圖3 菌株耐酸能力評價Fig.3 Evaluation of acid resistance of strains
2.3.2 膽鹽耐受性
微生物在有膽鹽的環境中生長會受到強烈抑制。因此,只有那些對高濃度膽鹽有較強耐受性的菌株才能在人體腸道內存活下來并發揮作用[26]。如圖4所示,植物乳桿菌1-3相對于其他兩株菌有較強的膽鹽耐受性,在含量為0.1%、0.2%、0.3%的膽鹽環境中生長滯后時間約為1 h、2 h、4 h。菌株4-1的生長滯后時間約為2 h、4 h、7 h。菌株1-6的生長滯后時間分別為2 h、6 h在膽鹽含量為0.3%的環境中生長10 h后OD620nm值仍然沒有達到上升0.3,說明菌株濃度為0.3%的膽鹽對菌株1-6的生長產生強烈的抑制作用。
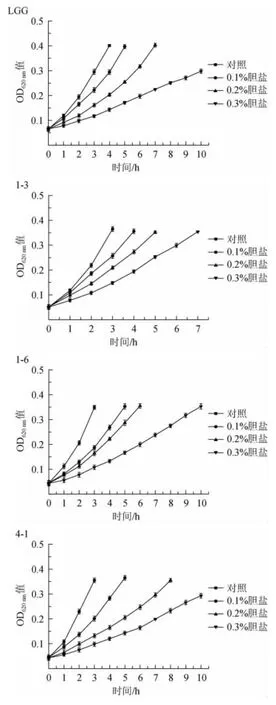
圖4 菌株膽鹽耐受性評價Fig.4 Evaluation of bile salt tolerance of strains
2.3.3 菌株模擬腸胃耐受性
人體胃液酸性一般在pH 3左右,但由于受到飲食等因素的影響。pH值在1.5~5.0之間波動,胃液的酸性會激活胃蛋白酶原,殺死進入胃中的微生物。所以益生菌對胃液中的酸性環境和胃蛋白酶有一定的耐受性是其在人體內發揮益生作用的前提[27]。人體小腸是益生菌發揮作用的場所,但小腸的高膽鹽環境會抑制益生菌的活性,所以益生菌對腸液也要有一定的耐受性才能發揮其益生作用[28]。3株菌的胃腸液耐受性結果如圖5所示。由圖5可知,菌株4-1在人工胃液中的存活率最高達到68.74%,菌株1-6存活率最低53.07%。但菌株1-3在人工腸液中有最大的存活率達到69.97%,菌株4-1存活率最低50.01%。3株植物乳桿菌在人工胃腸液中的存活率均>50%,說明3株植物乳桿菌對模擬人工胃腸均有較高的耐受性。
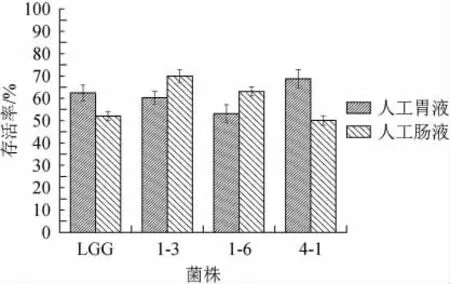
圖5 菌株的人工胃腸液耐受性評價Fig.5 Evaluation of artificial gastrointestinal tolerance of strains
2.3.4 菌株DPPH自由基清除率
DPPH在有機溶劑中是一種穩定的自由基,對DPPH自由基的清除可反應出菌株具有降低過氧化自由基、烷自由基或脂質自由基連鎖反應的特性[29]。三株菌對DPPH自由基清除能力如圖6所示。由圖6可知,菌株1-3對DPPH自由基清除率(50.03%)明顯高于對照菌株LGG(32.77%)和菌株1-6、菌株4-1(P<0.05),菌株4-1的DPPH自由基清除率基本與參考菌株LGG相同。目前已有研究表明植物乳桿菌DPPH自由基清除率為(8.39%~45.97%)這與目前的研究結果相類似[30-31]。XU Y等[32]研究發現L.plantarumKX041產生的EPS具有較好的清除DPPH自由基和羥基自由基的能力。本研究中3株產胞外多糖EPS的植物乳桿菌對DPPH自由基有較強的清除能力,推測其抗氧化活性與EPS活性有一定的關系,但還需進行實驗驗證。
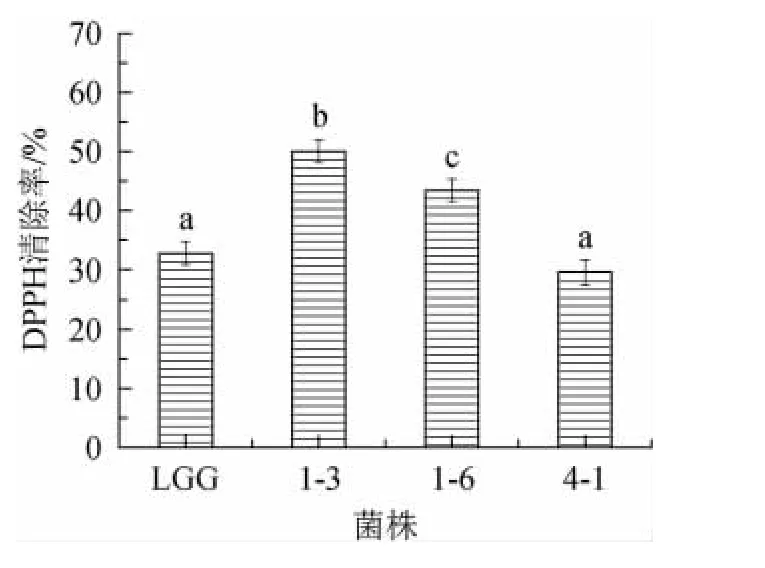
圖6 菌株對DPPH自由基的清除率Fig.6 Scavenging rate of DPPH free radicals by strains
2.3.5 菌株的表面疏水性和自凝聚率
乳酸菌菌株的細胞表面疏水性和自聚集能力與粘附能力和腸道的定植有關[33]。自凝集是同一種菌株間相互凝集形成多細胞簇的現象,益生菌通過自凝集作用相互凝集到一定的數量時粘附才牢固,進而達到菌株發揮益生功效所需要的數量。而疏水性是指非極性溶液(溶質)在極性水中所呈現的不穩定狀態,從而引起一系列熱能(熵)和分子重新分布及排列的變化。疏水性較高的菌株,對小腸組織有較高的粘附能力。菌株的自凝聚和疏水性如表2所示。由表2可知,不同菌株之間自凝聚能力和表面疏水性存在一定差異,菌株1-3的表面疏水性和自凝聚能力顯著高于其他菌株(P<0.05)。3株植物乳桿菌都表現出一定自凝聚能力和疏水性,說明該菌株有黏附在人體腸道上皮細胞和黏膜表面的可能性。

表2 菌株自凝聚及表面疏水性Table 2 Self-aggregation and surface hydrophobicity of strains
2.3.6 膽固醇降解率
高膽固醇血癥是導致人類發病和死亡的一個重大的健康問題。近年來大量的體內外實驗證明植物乳桿菌能夠有效地降低膽固醇。由圖7可知,在添加膽鹽的培養基中3株植物乳桿菌降解膽固醇的能力均有所提高,表明膽鹽存在對膽固醇的降解率有一定的促進作用。但隨著膽鹽濃度的升高其膽固醇降解率有所下降。可能是因為過高濃度的膽鹽抑制乳酸菌的生長。3株植物乳桿菌都在0.1%的膽鹽濃度中有最高的膽固醇降解率。但菌株1-3的膽固醇降解率達到36.98%。這與李妮等[34]的研究結果相類似。
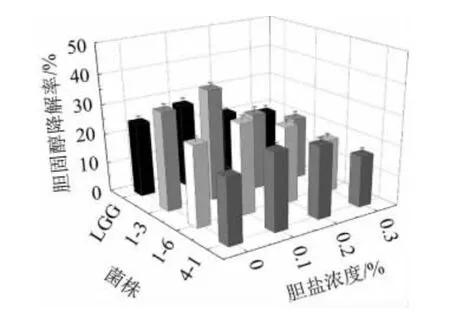
圖7 菌株的膽固醇降解率Fig.7 Cholesterol degradation rate of strains
2.3.7 抑菌能力評價
如圖8所示,植物乳桿菌1-3、1-6對大腸桿菌和金黃色葡糖球菌的抑制作用顯著大于菌株4-1(P<0.05)。對大腸桿菌的抑菌圈直徑都達到19 mm。對金黃色葡糖球菌的抑菌圈直徑都達到18 mm。而菌株4-1對大腸桿菌和金黃葡糖球菌的抑制作用較菌株1-3、1-6弱。抑菌圈直徑在15 mm左右。宋瑩龍[35]研究發現植物乳桿菌12產生的胞外多糖對大腸桿菌,金黃色葡糖球菌等6種致病菌生物膜的形成有抑制作用。本研究中3株植物乳桿菌的高抑菌能力除了與其本身的作用之外可能也和其產生的胞外多糖的活性有關。

圖8 菌株1-3和1-6對大腸桿菌和金黃色葡糖球菌的抑制作用Fig.8 Inhibition of strains 1-3 and 1-6 against Escherichia coli and Staphylococcus aureus
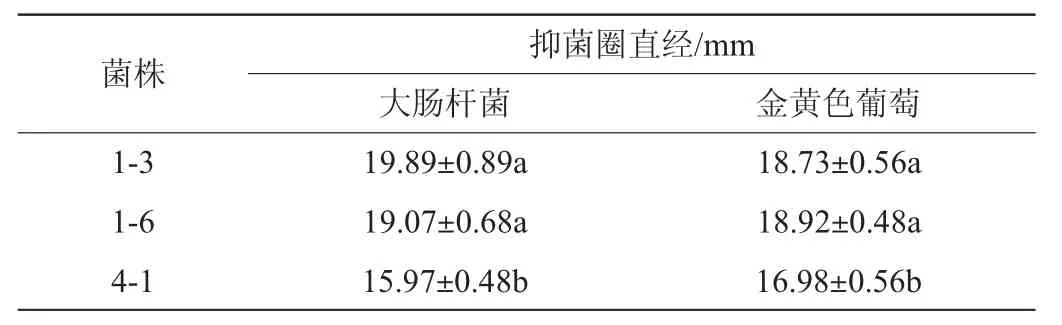
表3 菌株對致病菌的抑菌活性Table 3 Inhibitory activities of strain against pathogens
3 結論
從新疆傳統乳品中分離出147株乳酸菌,通過表型鑒定及苯酚硫酸篩選出22株產胞外多糖含量較高的菌株,其中三株植物乳桿菌1-3、1-6、4-1的產量達到240 mg/L。潛在益生特性研究結果顯示,菌株1-3、1-6、4-1在酸性環境中的活菌總數都達到107CFU/mL,在模擬人工胃腸液中的存活率都大于50%。菌株1-3表現出最高的膽鹽耐受性,對DPPH自由基的清除能力達到50%、自凝聚力最大為59.3%及對二甲苯的粘附性為68.3%,在膽鹽濃度為0.1%時膽固醇去除率最高達到36.98%,且對大腸桿菌和金黃色葡萄球菌有較強的抑制能力。因此植物乳桿菌1-3可作為潛在益生菌用于功能性產品的開發。
