新疆哈薩克族風(fēng)干肉中產(chǎn)蛋白酶乳酸菌的篩選及酶學(xué)特性研究
2020-06-06 04:55:06李丹陽(yáng)李宇輝高云云閆藝王俊鋼盧士玲
食品與發(fā)酵工業(yè) 2020年9期
關(guān)鍵詞:生長(zhǎng)
李丹陽(yáng),李宇輝,高云云,閆藝,王俊鋼*,盧士玲*
1(石河子大學(xué) 食品學(xué)院,新疆 石河子,832000) 2(新疆農(nóng)墾科學(xué)院農(nóng)產(chǎn)品加工研究所,新疆 石河子,832000) 3(新疆農(nóng)墾科學(xué)院農(nóng)產(chǎn)品加工重點(diǎn)實(shí)驗(yàn)室,新疆 石河子,832000)
新疆風(fēng)干肉是新疆游牧民族的特色肉制品之一,其中以塔城風(fēng)干肉最為有名。風(fēng)干肉又分為風(fēng)干牛肉、風(fēng)干馬肉、風(fēng)干羊肉[1]。新疆風(fēng)干肉的制作主要在秋冬季節(jié),以牛肉、馬肉或羊肉為原料,進(jìn)行切塊、短時(shí)腌制,再經(jīng)風(fēng)干后熟制作而成,屬于干腌發(fā)酵肉制品。風(fēng)干肉風(fēng)味獨(dú)特,其揮發(fā)性風(fēng)味物質(zhì)主要來(lái)源于脂肪氧化、香辛料添加及氨基酸Strecker降解[2]。氨基酸的Strecker降解主要是使肉中蛋白質(zhì)經(jīng)過(guò)降解形成多肽、小肽、氨基酸和胺,蛋白質(zhì)降解主要是由于肉中內(nèi)源性的酶以及產(chǎn)蛋白酶微生物的催化[3]。
產(chǎn)蛋白酶乳酸菌所產(chǎn)的蛋白酶可以在肉制品發(fā)酵過(guò)程中對(duì)蛋白質(zhì)進(jìn)行降解,是肉制品風(fēng)味形成的基礎(chǔ)。TODOROV等[4]研究表明,乳酸菌通過(guò)引起pH降低來(lái)影響蛋白質(zhì)降解,從而導(dǎo)致肌肉蛋白酶的活性增加。更重要的是,乳酸菌所產(chǎn)的蛋白酶吸收了由肌肉蛋白水解產(chǎn)生的肽,并將這些肽在細(xì)胞內(nèi)分裂成氨基酸,轉(zhuǎn)化為芳香成分。乳酸菌蛋白水解系統(tǒng)按其功能可分為3個(gè)部分:首先,細(xì)胞外蛋白酶作用于底物蛋白并將肉蛋白分解為肽;第二,肽酶水解肽;最終,分解產(chǎn)物穿過(guò)細(xì)胞質(zhì)膜[5]肽是肉味物質(zhì)的前體[6]。乳酸菌蛋白酶在發(fā)酵過(guò)程中促進(jìn)了游離氨基酸和短肽濃度的增加[7-8],對(duì)發(fā)酵肉的成熟具有積極作用[9],而其代謝產(chǎn)物則促進(jìn)了風(fēng)味的形成[10]。周才瓊等[11]對(duì)酸肉發(fā)酵過(guò)程中揮發(fā)性風(fēng)味物質(zhì)形成的研究得出結(jié)論,通過(guò)微生物生長(zhǎng)繁殖分泌胞外酶,導(dǎo)致脂肪及蛋白質(zhì)降解,產(chǎn)生與發(fā)酵風(fēng)味形成有關(guān)的醛、酮、醇、酸等物質(zhì),并發(fā)生酯化反應(yīng),形成了酸肉的特有風(fēng)味。ESSID等[12]從突尼斯傳統(tǒng)咸肉中分離出具有良好抗菌性和耐酸性的乳酸菌,并且所分離的乳酸菌都具有良好的水解酪蛋白的能力。林偉濤等[13]從中式發(fā)酵香腸中分離篩選出具有當(dāng)?shù)靥厣淖匀话l(fā)酵香腸風(fēng)味乳酸菌,它們都具有很好的產(chǎn)酸和產(chǎn)酶特性。SUN等[14]從哈爾濱干香腸中分離出1株乳酸菌產(chǎn)蛋白酶可以水解肌原纖維和肌漿蛋白,所產(chǎn)蛋白酶在pH 6和40 ℃下具有活性。有關(guān)新疆風(fēng)干肉中分離乳酸菌的研究主要集中在乳酸脫氫酶、亞硝酸還原酶和酯酶的特性方面[1],對(duì)新疆傳統(tǒng)風(fēng)干肉中產(chǎn)蛋白酶乳酸菌對(duì)其品質(zhì)影響的機(jī)制尚不明確。
本實(shí)驗(yàn)以新疆風(fēng)干肉中高產(chǎn)蛋白酶乳酸菌為研究對(duì)象,系統(tǒng)研究了菌株產(chǎn)酶特性和蛋白酶性質(zhì),以期為風(fēng)干肉的工業(yè)化生產(chǎn)提供理論數(shù)據(jù)。
1 材料與方法
1.1 實(shí)驗(yàn)原料、培養(yǎng)基與試劑
1.1.1 實(shí)驗(yàn)原料
風(fēng)干牛肉和風(fēng)干馬肉樣品:購(gòu)于新疆塔城、額敏、富蘊(yùn)、托里地區(qū)。
1.1.2 培養(yǎng)基
MRS肉湯、MRS固體培養(yǎng)基:青島海博生物技術(shù)有限公司;MRS脫脂乳固體培養(yǎng)基:在MRS固體培養(yǎng)基中加入體積分?jǐn)?shù)為1%的脫脂乳。
1.1.3 試劑
福林試劑、無(wú)水Na2CO3、三氯乙酸、NaOH、濃HCl、H3PO4、乳酸、乳酸鈉、冰乙酸、干酪素、L-酪氨酸、無(wú)水乙醇、EDTA等,均為國(guó)產(chǎn)分析純;細(xì)菌DNA提取試劑盒,天根生化科技(北京)有限公司;2×TaqMaster Mix (Dye Plus),南京諾唯贊生物科技有限公司;瓊脂糖西班牙,Biowest公司。
1.2 儀器與設(shè)備
22331型高速冷凍離心機(jī),德國(guó)Eppendorf AG公司;HP1020凝膠成像系統(tǒng),美國(guó)BIO-RAD伯樂(lè)公司;T100型PCR儀,美國(guó)BIO-RAD伯樂(lè)公司;CX23LEDRFS1C生物顯微鏡,奧林巴斯(廣州)工業(yè)有限公司;LDZX-30KBS立式壓力蒸汽滅菌器,上海申安醫(yī)療器械廠;EONC酶標(biāo)儀,美國(guó)伯騰儀器有限公司;水平電泳槽(小型),美國(guó)Bio-Rad公司;核酸測(cè)定儀,美國(guó)賽默飛世爾科技公司;UB-7型pH計(jì),美國(guó)賽多利斯丹佛公司。
1.3 實(shí)驗(yàn)方法
1.3.1 乳酸菌的篩選分離、純化與鑒定
參考林偉濤等[13]的方法并作適當(dāng)修改。在無(wú)菌條件下取10 g風(fēng)干肉樣品充分剪碎,放入滅菌后的90 mL生理鹽水中,37 ℃、120 r/min培養(yǎng)24 h。吸取1mL菌液用生理鹽水進(jìn)行10倍梯度稀釋,每次稀釋均需用旋渦振蕩器儀充分混勻并更換無(wú)菌槍頭。分別吸取10-5~10-7梯度的稀釋液200 μL稀釋液均勻涂布于MRS培養(yǎng)基中,每個(gè)梯度做3個(gè)平行,于37 ℃恒溫培養(yǎng)箱中培養(yǎng)48 h。挑取不同形態(tài)的菌落進(jìn)行劃線培養(yǎng),重復(fù)劃線多次,直至用顯微鏡觀察至純種,記錄其菌落、細(xì)胞形態(tài)。挑選純化好的菌株進(jìn)行革蘭氏染色以及H2O2實(shí)驗(yàn),選擇革蘭氏染色呈陽(yáng)性,H2O2實(shí)驗(yàn)呈陰性的乳酸菌用體積分?jǐn)?shù)50%甘油/菌液(體積比為1∶1)于-80 ℃冰箱保存?zhèn)溆谩?/p>
1.3.2 乳酸菌菌株生物學(xué)鑒定
1.3.2.1 乳酸菌DNA的提取
使用細(xì)菌基因組 DNA 提取試劑盒,北京天根生化科技限公司。按照說(shuō)明書(shū)操作步驟提取樣品中乳酸菌DNA。
1.3.2.2 16S rDNA測(cè)序鑒定
采用通用引物27f(5’-AGAGTTTGATCCTGGCTCAG-3’)和1492r:(5’CTACGGCTACCTTGTTACGA-3’),預(yù)變性95 ℃ 5 min;變性94 ℃ 30 s,退火50 ℃ 30 s,延伸72 ℃ 4 min共35個(gè)循環(huán);最后72 ℃延伸10 min[15]。PCR產(chǎn)物經(jīng) 1.0%瓊脂糖凝膠電泳檢測(cè)后,送往上海生工生物科技有限公司進(jìn)行測(cè)序,測(cè)序結(jié)果提交至 GenBank 數(shù)據(jù)庫(kù)中進(jìn)行序列同源性比對(duì)(BLAST),并用 MAGE 7.0 軟件構(gòu)建系統(tǒng)發(fā)育樹(shù)。
1.3.3 產(chǎn)蛋白酶乳酸菌的篩選
1.3.3.1 產(chǎn)蛋白酶乳酸菌的初篩
將保藏好的菌株以4.0%的接種量接入MRS液體培養(yǎng)基,于37 ℃恒溫培養(yǎng)箱中培養(yǎng)24 h后,在MRS固體培養(yǎng)基上進(jìn)行多次劃線至單菌落,挑單菌落到MRS液體培養(yǎng)基中于37 ℃恒溫培養(yǎng)箱中培養(yǎng)24 h。參考SUN等[14]方法并作適當(dāng)修改,采用牛津杯法初篩產(chǎn)蛋白酶高的菌株,配制好脫脂乳MRS固體培養(yǎng)基進(jìn)行倒板,將滅好菌的牛津杯放在培養(yǎng)基上,吸取已活化好的乳酸菌10 μL加入牛津杯中,37 ℃恒溫培養(yǎng)箱中培養(yǎng)24 h后觀察并測(cè)量透明圈的直徑。
1.3.3.2 產(chǎn)蛋白酶乳酸菌的復(fù)篩
參照張曉燕[16]的方法初篩,得到22株產(chǎn)透明圈大的菌株進(jìn)行蛋白酶活力的測(cè)定。
1.3.3.3 粗酶液的提取
參照FARHIDIAN等[17]的方法分離粗蛋白酶,進(jìn)行少量修改。菌液添加到干凈的離心管中,然后在4 ℃下以 10 000 r/min 離心10 min。所獲得的上清液是粗制的細(xì)胞外蛋白酶提取物,保存在4 ℃ 下直至后續(xù)使用(12 h內(nèi)使用)。
1.3.3.4 蛋白酶活力的測(cè)定[16]
按照國(guó)標(biāo)GB/T23527—2009規(guī)定的福林酚顯色法(Folin)測(cè)酶活力。蛋白酶活性定義:在pH 7.0和40 ℃下,每分鐘水解酪蛋白產(chǎn)生1 μg酪氨酸,定義為1個(gè)蛋白酶活力單位。同一個(gè)樣品測(cè)3次,取平均值。
酶液適當(dāng)稀釋,取4支試管編號(hào),1號(hào)為空白對(duì)照,分別加入1 mL酶液,立即加入0.4 mol/L TCA溶液2 mL,使酶失活。另3支試管中加入pH 7.0、質(zhì)量濃度為20 g/L的酪蛋白溶液1 mL,充分振蕩后,置于40 ℃恒溫水浴中保溫反應(yīng)10 min后,取出加入0.4 mol/L TCA溶液2 mL,終止反應(yīng)。同時(shí)在1號(hào)試管中加入20 g/L酪蛋白溶液1 mL,混勻后放入水浴中繼續(xù)保溫20 min,取出冷凍離心除去反應(yīng)沉淀,取離心后濾液1 mL移入干凈的4支新試管中,再加入0.4 mol/L Na2CO3溶液5 mL和稀釋度為1∶2的Folin試劑1 mL,充分搖勻,保溫顯色20 min后,迅速冷卻測(cè)定吸光值(OD680nm)。根據(jù)標(biāo)準(zhǔn)曲線,得到酪氨酸相當(dāng)量。按公式(1)計(jì)算:
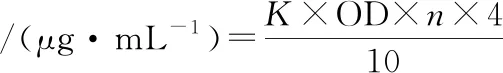
(1)
式中:K,標(biāo)準(zhǔn)曲線斜率的倒數(shù);4,4 mL反應(yīng)液取出 1 mL測(cè)定(即 4倍);n,酶液稀釋的倍數(shù);10,反應(yīng)10 min。
1.3.4 乳酸菌生物學(xué)特性分析
1.3.4.1 菌株最適生長(zhǎng)溫度
參照桑鵬等[18]的方法并做適當(dāng)修改。按4.0%的接種量接于MRS液體培養(yǎng)基中,置于不同的溫度(25、30、37、40、50、55、60 ℃)下?lián)u床培養(yǎng) 24 h,然后在波長(zhǎng)600 nm 處測(cè)定各個(gè)溫度下菌株生長(zhǎng)的吸光度值,以確定菌株的最適生長(zhǎng)溫度。
1.3.4.2 pH對(duì)乳酸菌生長(zhǎng)的影響
參考張大為等[19]的方法并作適當(dāng)修改。用冰乙酸將MRS液體培養(yǎng)基的pH值分別調(diào)節(jié)為1.5、2.0、2.5、3.0、3.5、4.0、4.5、5.0、5.5、6.0、6.5,以pH為6.0的自然培養(yǎng)作為對(duì)照,每個(gè)梯度3個(gè)平行。向液體培養(yǎng)基中接種分離純化的乳酸菌,放于 37 ℃的培養(yǎng)箱培養(yǎng) 24 h,測(cè)定600 nm處的吸光值。
1.3.4.3 乳酸菌耐鹽能力的評(píng)價(jià)
參考張大為等[19]的方法并作適當(dāng)修改。向質(zhì)量濃度為10、20、30、40、50、 60和70 g/L的 MRS 液體培養(yǎng)基中接種分離純化的乳酸菌,各留1支做空白試驗(yàn),放于37 ℃培養(yǎng)箱培養(yǎng)24 h,在波長(zhǎng)為600 nm處測(cè)量吸光值。
1.3.5 蛋白酶酶學(xué)性質(zhì)的研究
1.3.5.1 溫度對(duì)酶活力的影響[14]
以10 000 r/min,4 ℃,10 min提取粗酶液,分別在0~60 ℃條件下測(cè)定酶活力。
1.3.5.2 pH對(duì)酶活力的影響[14]
以10 000 r/min,4 ℃,10 min提取粗酶液,用不同緩沖液調(diào)整粗酶液的pH為0~11,在最適溫度下測(cè)定酶活力。
1.3.5.3 金屬離子和抑制劑對(duì)蛋白酶活性的影響
參照SUN等[14]的方法并作適當(dāng)修改。金屬離子和抑制劑對(duì)蛋白酶活性的影響的實(shí)驗(yàn)進(jìn)行3次平行,3次重復(fù)。將濃度為1和10 mmol/L不同的金屬離子(Na+、K+、Mg2+、Ca2+、Mn2+、Zn2+、Cu2+、Fe2+和Fe3+)和抑制劑EDTA溶解在 pH值為6.8的0.02 mol/L磷酸鹽緩沖液中的20 g/L酪蛋白溶液中,以測(cè)定其對(duì)蛋白酶活性的影響。隨后,將粗酶液與每種離子的氯化物鹽溶液(1 mL)或蛋白酶抑制劑在37 ℃孵育30 min,然后對(duì)蛋白酶的活力進(jìn)行測(cè)定。以沒(méi)有添加任何金屬離子或抑制劑的蛋白酶活性定義為 100%,作為對(duì)照。
1.4 數(shù)據(jù)與處理
使用Origin 2017作圖,利用SPSS 25.0進(jìn)行差異性分析;實(shí)驗(yàn)數(shù)據(jù)均進(jìn)行3次平行3次重復(fù),以平均值±標(biāo)準(zhǔn)偏差表示,利用 MEGA 7.0軟件構(gòu)建系統(tǒng)發(fā)育樹(shù)。數(shù)據(jù)分析過(guò)程中,當(dāng)P<0.05,具有顯著性差異。
2 結(jié)果與分析
2.1 乳酸菌的分離與鑒定
從樣品中共分離得到161株純培養(yǎng)物,經(jīng)過(guò)革蘭氏染色和過(guò)氧化氫酶生理生化實(shí)驗(yàn),初步鑒定149株乳酸菌。通過(guò)16S rDNA基因檢測(cè)分析,確定149株乳酸菌。其中優(yōu)勢(shì)菌種為糞腸球菌39株約占26%,乳酸乳球菌28株約占19%,格氏乳球菌22株約占15%,植物乳桿菌18株約占12%,其余分別為戊糖片球菌10株、耐久腸球菌9株、腸系膜明串珠菌8株、路德維希腸桿菌6株、發(fā)酵乳桿菌5株、副干酪乳桿菌3株、羅伊氏乳桿菌1株。
2.2 產(chǎn)蛋白酶乳酸菌的篩選
根據(jù)制作酪氨酸標(biāo)準(zhǔn)曲線的方法,得出酪氨酸標(biāo)準(zhǔn)曲線線性回歸方程為y=0.005 2x-0.004 1,線性相關(guān)系數(shù)R2=0.999 2,表明線性關(guān)系良好。
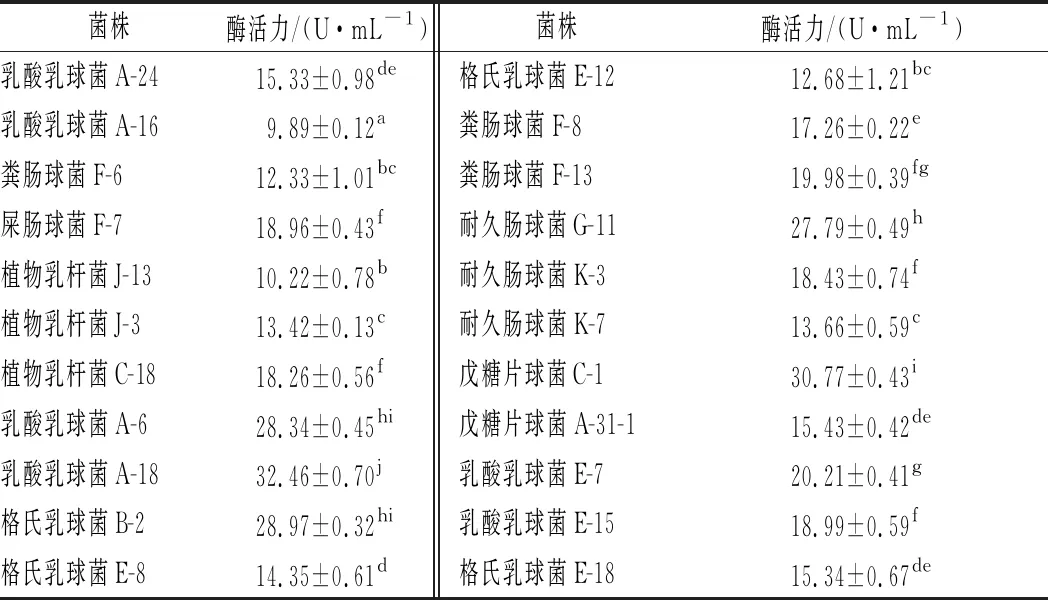
在所篩選的乳桿菌中初篩出22株在脫脂乳MRS瓊脂培養(yǎng)基中有明顯透明圈的菌株,如圖1所示。用福林酚法對(duì)初篩的22株菌株產(chǎn)蛋白酶活性進(jìn)行定量,實(shí)驗(yàn)結(jié)果顯示出5株產(chǎn)蛋白酶較高的菌株分別是乳酸乳球菌A-6、A-18,格氏乳球菌B-2,耐久腸球菌G-11,戊糖片球菌C-1。菌株產(chǎn)蛋白酶活力見(jiàn)表1。

a-空白;b-脫脂乳瓊脂培養(yǎng)基;c-脫脂乳MRS培養(yǎng)基圖1 乳酸菌水解脫脂乳產(chǎn)生透明圈的情況Fig.1 The transparent circles produced by the hydrolysisof skim milk by lactic acid bacteria
表1 菌株產(chǎn)蛋白酶的酶活力Table 1 Activity of protease produced by the strain
菌株酶活力/(U·mL-1)菌株酶活力/(U·mL-1)乳酸乳球菌A-2415.33±0.98de格氏乳球菌E-1212.68±1.21bc乳酸乳球菌A-169.89±0.12a糞腸球菌F-817.26±0.22e糞腸球菌F-612.33±1.01bc糞腸球菌F-1319.98±0.39fg屎腸球菌F-718.96±0.43f耐久腸球菌G-1127.79±0.49h植物乳桿菌J-1310.22±0.78b耐久腸球菌K-318.43±0.74f植物乳桿菌J-313.42±0.13c耐久腸球菌K-713.66±0.59c植物乳桿菌C-1818.26±0.56f戊糖片球菌C-130.77±0.43i乳酸乳球菌A-628.34±0.45hi戊糖片球菌A-31-115.43±0.42de乳酸乳球菌A-1832.46±0.70j乳酸乳球菌E-720.21±0.41g格氏乳球菌B-228.97±0.32hi乳酸乳球菌E-1518.99±0.59f格氏乳球菌E-814.35±0.61d格氏乳球菌E-1815.34±0.67de
注: 同一欄中不同字母表示有顯著性差異(P<0.05)
2.3 菌株生物學(xué)特性分析
2.3.1 菌株的最適生長(zhǎng)溫度
圖2為溫度對(duì)菌株的影響。從圖2可知,5株菌在25~40 ℃,隨著溫度的上升生長(zhǎng)量也迅速上升(P<0.05),乳酸菌在40 ℃,MRS液體培養(yǎng)基中,600 nm下的吸光值達(dá)到最大,說(shuō)明5株菌的最適生長(zhǎng)溫度都為40 ℃左右。在40 ℃以后,菌株的生長(zhǎng)量隨著溫度的升高逐漸下降,到45 ℃之后5株菌幾乎都停止生長(zhǎng)(P>0.05)。在整個(gè)考察范圍內(nèi)菌株A-18具有較好的生長(zhǎng)活性。
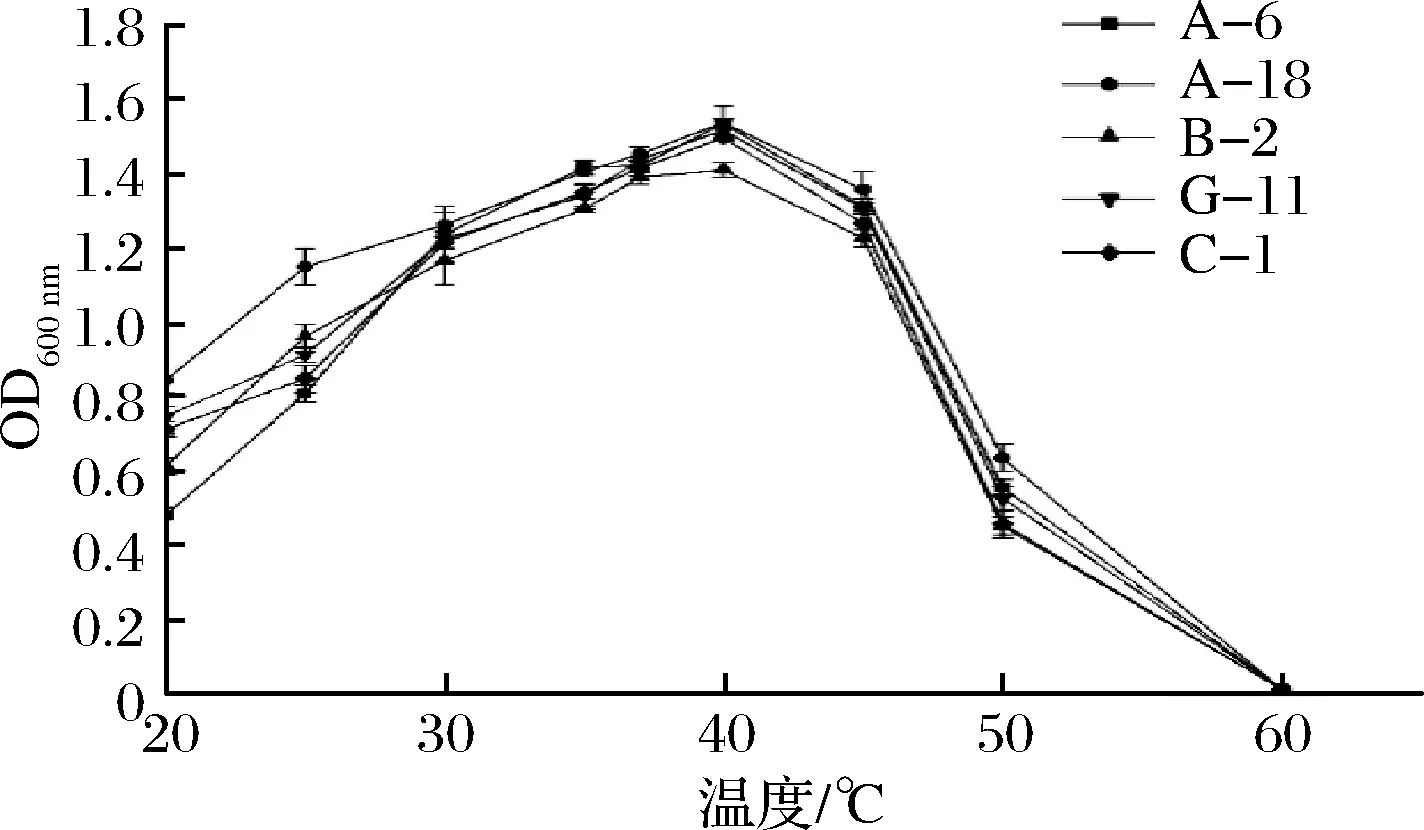
圖2 溫度對(duì)菌株生長(zhǎng)的影響Fig.2 Effect of temperature on strain growth
2.3.2 pH對(duì)菌株生長(zhǎng)的影響
乳酸菌的大量繁殖使得肉中pH降低[20],因此確定菌株的耐酸性十分重要。將5株菌株接種于不同pH的培養(yǎng)基中,培養(yǎng)24 h。從圖3可以看出,5株菌中G-11的耐酸能力較弱,其余4株菌A-6、A-18、B-2、C-1都具有較好的耐酸能力,在pH值為3.0~3.5的范圍內(nèi)仍能生長(zhǎng),在pH值為1.5~2.5時(shí)菌株基本不生長(zhǎng)。羅強(qiáng)等[21]在乳酸菌耐酸能力實(shí)驗(yàn)中對(duì)乳酸菌在pH 2.0條件下生長(zhǎng)情況進(jìn)行研究,發(fā)現(xiàn)pH 2.0時(shí)菌株存活率在70%,這可能是不同乳酸菌對(duì)酸的耐受性是不一致的。翟磊等[24]在乳酸菌耐酸能力實(shí)驗(yàn)中得出在pH低于2.0時(shí)菌株基本不生長(zhǎng),在pH值為3.0~3.5的范圍內(nèi)仍能生長(zhǎng)。這與本實(shí)驗(yàn)研究結(jié)果相似。通過(guò)分析比較得到5株菌在整個(gè)pH值考察范圍內(nèi),菌株A-18和菌株C-1的耐酸能力較強(qiáng)(P<0.05)。
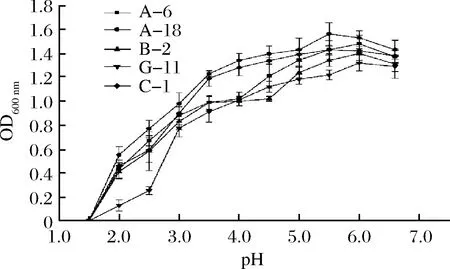
圖3 不同pH值對(duì)菌株生長(zhǎng)的影響Fig.3 Influence of different pH values on strain growth
2.3.3 菌株耐鹽能力
鹽脅迫引起的滲透壓變化會(huì)引起乳酸菌細(xì)胞結(jié)構(gòu)損傷,導(dǎo)致細(xì)胞生理代謝活動(dòng)紊亂甚至死亡,因此,乳酸菌在鹽脅迫條件下生存、生長(zhǎng)和代謝的能力在食品發(fā)酵過(guò)程中是非常重要的[23]。將5株菌分別接種于不同鹽濃度的MRS液體培養(yǎng)基中,37 ℃培養(yǎng)24 h考察各菌株對(duì)不同鹽濃度的耐受性,如圖4所示。
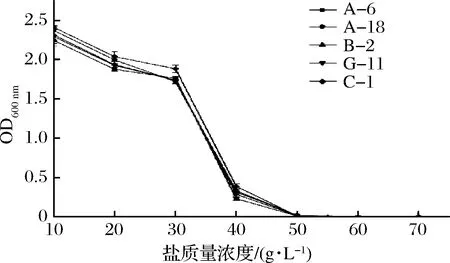
圖4 不同鹽濃度對(duì)菌株生長(zhǎng)的影響Fig.4 Effects of different salt concentrations on strain growth
5株菌隨著鹽質(zhì)量濃度和發(fā)酵液滲透壓的增加,生長(zhǎng)逐步受到抑制,在鹽質(zhì)量濃度為10~30 g/L的范圍內(nèi)時(shí),5株菌均生長(zhǎng)良好,在30~40 g/L的范圍內(nèi)時(shí),菌株生長(zhǎng)受到了抑制。鹽質(zhì)量濃度在40 g/L以上時(shí)菌株停止生長(zhǎng)(P<0.05)。張大為等[19]在乳酸菌耐鹽能力實(shí)驗(yàn)中得出鹽質(zhì)量濃度在10~3 g/L之間,菌株均生長(zhǎng)良好,鹽質(zhì)量濃度達(dá)到30~4 g/L時(shí),菌體能夠生長(zhǎng),但生長(zhǎng)受到抑制。在整個(gè)鹽濃度考察范圍內(nèi),5株菌中,菌株C-1和菌株B-2的耐鹽能力較優(yōu)(P<0.05)。
2.4 菌株蛋白酶的酶學(xué)性質(zhì)
2.4.1 溫度對(duì)酶活力的影響
從圖5所示,在4~10 ℃低溫條件下,5株菌的蛋白酶活性均受到抑制,但是在低溫范圍內(nèi),蛋白酶仍具備一定的活性,但活性較低。在10~40 ℃的溫度范圍內(nèi),隨著溫度的升高,5株菌的酶活力都呈明顯上升趨勢(shì)(P<0.05),40 ℃時(shí)酶的活力達(dá)到最高,如圖5所示。在40~60 ℃范圍內(nèi),隨著溫度的升高,蛋白酶的活力逐漸下降。說(shuō)明40 ℃為蛋白酶的反應(yīng)活力最強(qiáng)。
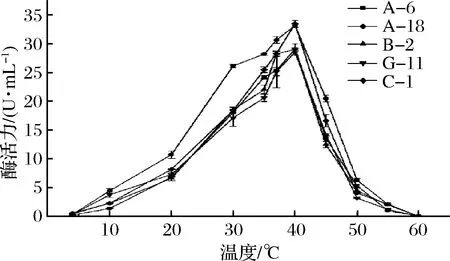
圖5 溫度對(duì)蛋白酶活性的影響Fig.5 Effect of temperature on the activity of protease
2.4.2 蛋白酶的最適反應(yīng)pH
培養(yǎng)基的初始pH會(huì)影響各種生長(zhǎng)因子在細(xì)胞膜上的轉(zhuǎn)運(yùn)過(guò)程,這可能會(huì)對(duì)微生物蛋白酶的生產(chǎn)過(guò)程產(chǎn)生不利影響。如果微生物處于直接或間接影響微生物蛋白酶活性的最佳pH,則代謝效率將很高[24],如圖6所示。
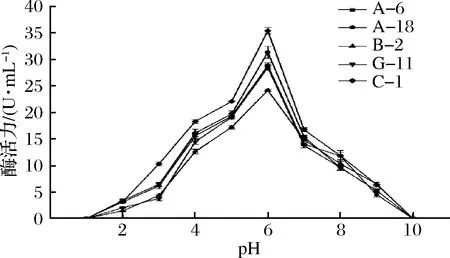
圖6 pH對(duì)蛋白酶活性的影響Fig.6 Effect of pH on protease activity of strain
在pH值為1~6的范圍內(nèi),5株菌株的蛋白酶活力隨著pH的上升均呈上升趨勢(shì)(P<0.05),在pH值為6時(shí)蛋白酶活性最強(qiáng)。在pH值為6以上時(shí),蛋白酶活力逐漸下降。說(shuō)明這5株菌株蛋白酶最適反應(yīng)pH值均為6,5株菌產(chǎn)生的蛋白酶為酸性蛋白酶。
pH是影響蛋白酶活性的重要指標(biāo)之一[25]。蛋白酶活性在pH 范圍內(nèi)的穩(wěn)定性對(duì)于發(fā)酵肉制品的質(zhì)量至關(guān)重要,并且嫩度和風(fēng)味前體也可以得到改善[26]。
2.4.3 金屬離子和抑制劑對(duì)蛋白酶活性的影響
肉品加工過(guò)程中會(huì)使用含Na+、K+、Mg2+的鹽,會(huì)影響乳酸菌的蛋白酶活性。金屬離子可能是蛋白酶活性位點(diǎn)的一部分,并直接參與催化過(guò)程[28]。在離子濃度為1 mmol/L的條件下研究了各種金屬離子對(duì)5株菌株蛋白酶相對(duì)活性的影響,如表2所示。在10 mmol/L的條件下研究了各種金屬離子對(duì)5株菌株蛋白酶相對(duì)活性的影響,如表3所示。以沒(méi)有添加金屬離子的蛋白酶用作對(duì)照(100%)。
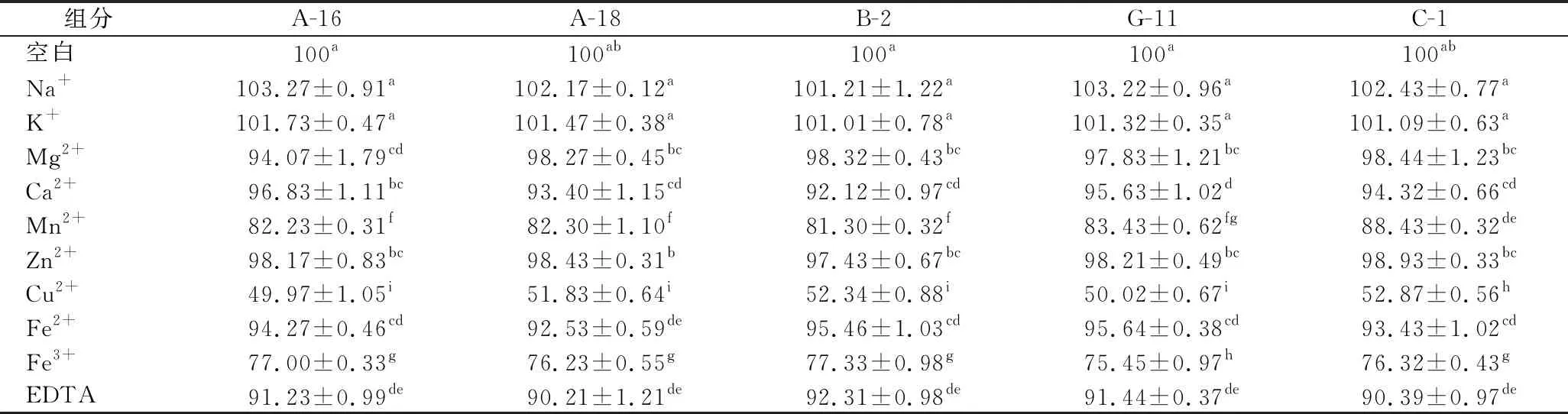
表2 1 mmol/L金屬離子和抑制劑對(duì)5株菌株蛋白酶活性的影響Table 2 Effect of 1 mmol/L metal ions and inhibitors on protease activity of five strains
注: 同一欄中不同字母表示有顯著性差異(P<0.05)(下同)
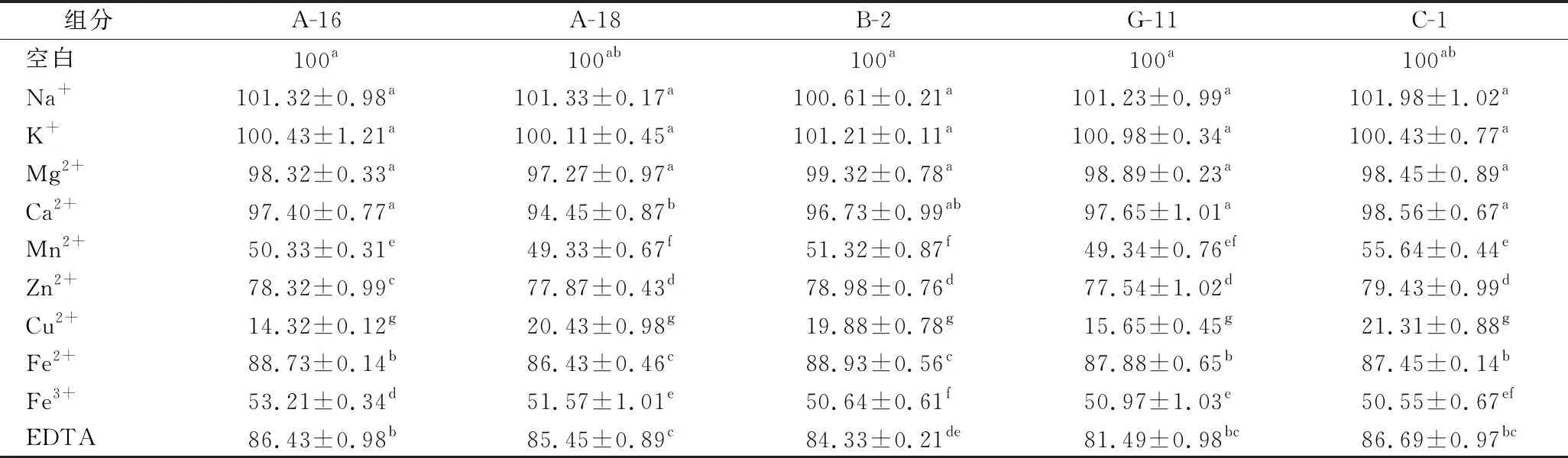
表3 10 mmol/L金屬離子和抑制劑對(duì)5株菌株蛋白酶活性的影響Table 3 Effect of 10 mmol/L metal ions and inhibitors on protease activity of five strains
Na+和 K+在1和10 mmol/L 濃度下誘導(dǎo)5株菌株蛋白酶活性略有增加(P>0.05)。Mn2+、Cu2+和Fe3+對(duì)蛋白酶的活性有抑制作用。與對(duì)照相比,5株菌株在Cu2+為1和10 mmol/L的條件下蛋白酶的相對(duì)活性抑制明顯(P<0.05)。這可能是由于這些離子與5株菌的蛋白酶?jìng)?cè)鏈基團(tuán)之間的牢固結(jié)合,從而降低了蛋白酶與其底物之間的結(jié)合[29]。SUN等[14]研究表明Cu2+確實(shí)會(huì)顯著降低蛋白酶的活性。EDTA在濃度為1和10 mmol/L時(shí),對(duì)蛋白酶活性降低顯著(P<0.05),這與YU等[30]報(bào)道類似。
3 結(jié)論
本研究從新疆哈薩克族風(fēng)干肉中分離出149株乳酸菌,通過(guò)表型鑒定及脫脂乳MRS瓊脂培養(yǎng)基初篩出22株在脫脂乳MRS瓊脂培養(yǎng)基中有明顯透明圈的菌株,用福林酚法對(duì)初篩的22株菌株產(chǎn)蛋白酶活性進(jìn)行定量,實(shí)驗(yàn)結(jié)果顯示出5株產(chǎn)蛋白酶活力較高的菌株,5株菌株為:乳酸乳球菌A-6、A-18、格氏乳球菌B-2、耐久腸球菌G-11、戊糖片球菌C-1。
菌株生物學(xué)特性分析研究結(jié)果顯示,所有菌株的最適生長(zhǎng)溫度為40 ℃,5株菌中菌株A-18具有較高的生長(zhǎng)活性。菌株耐酸特性分析表示5株菌中株菌中G-11的耐酸能力較弱,其余4株菌A-6、A-18、B-2、C-1都具有較好的耐酸能力在pH值為3~3.5的條件下均能生長(zhǎng),整個(gè)pH考察范圍內(nèi)菌株A-18和菌株C-1的耐酸能力較強(qiáng)。菌株耐鹽特性分析表示在鹽濃度為10~30 g/L的范圍內(nèi)時(shí),5株菌均生長(zhǎng)良好,具有一定的耐鹽能力,在整個(gè)鹽濃度考察范圍內(nèi),5株菌中,菌株C-1和菌株B-2的耐鹽能力較優(yōu)。
對(duì)蛋白酶酶學(xué)特性分析表示菌株產(chǎn)生的蛋白酶在40 ℃的反應(yīng)活力最強(qiáng)并且5株菌所產(chǎn)的蛋白酶均為酸性蛋白酶。Na+和K+在1和10 mmol/L濃度下誘導(dǎo)蛋白酶活性略有增加(P>0.05)。Mn2+、Zn2+、Cu2+和Fe3+對(duì)蛋白酶的活性有抑制作用。相比較之下Cu2+的存在對(duì)蛋白酶抑制作用明顯(P>0.05)。
根據(jù)研究結(jié)果乳酸乳球菌A-18、戊糖片球菌C-11產(chǎn)蛋白酶含量較高,具有較好的耐酸耐鹽特性,可作為潛在益生菌用于后續(xù)發(fā)酵肉制品品質(zhì)控制的研究。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(jí)(2020年3期)2020-03-17 09:24:33
瘋狂英語(yǔ)·新悅讀(2019年11期)2019-12-18 05:14:16
華人時(shí)刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
