LED不同光質補光對日光溫室冬春茬番茄生長及光合特性的影響
2020-06-01 07:58:51盧純張亞紅李青
江蘇農業科學 2020年8期
盧純 張亞紅 李青


摘要:以番茄品種巴菲亞特為材料,研究不同LED光質(紅光、藍光、紅藍5 ∶1、紅藍7 ∶1)補光對冬春季溫室番茄生長指標及光合作用的影響。結果表明:番茄幼苗期到盛果期,藍光補光可有效促進番茄生長,不同光質在番茄生長期、坐果期對葉綠素合成具有一定的影響。在生長期,藍光、紅光補光處理可有效增強番茄的光合作用;在開花期,紅光、紅藍7 ∶1補光處理可有效增強番茄的光合作用;在坐果期,紅光、紅藍5 ∶1補光處理可有效增強番茄光合作用;番茄生長期、開花期、坐果期各補光處理在14:00—16:00光合作用最為活躍;紅光在開花期凈光合效率最大,紅光、紅藍5 ∶1在結果期凈光合效率最大。番茄盛果后期在PSⅡ反應中心開放程度依次是紅藍 7 ∶1>紅藍5 ∶1>紅光>藍光>CK,紅藍7 ∶1在盛果后期更有利于電子傳遞。番茄不同補光處理下產量排序是紅光>紅藍5 ∶1>紅藍 7 ∶1>藍光>CK。總體上本研究結果可為溫室冬春季番茄種植的光調控提供參考。
關鍵詞:LED光質;番茄;生長指標;光合作用
中圖分類號: S641.201;Q945.11文獻標志碼: A
文章編號:1002-1302(2020)08-0127-08
收稿日期:2019-03-19
基金項目:寧夏回族自治區重點研發計劃(編號:2016BZ0901)。
作者簡介:盧?純(1986—),男,寧夏銀川人,碩士,研究方向為蔬菜學。E-mail:917998377@qq.com。
光是促進植物生長發育進行光合作用、新陳代謝、基因表達和其他生理反應的重要能源。光在高等植物中表達可分為2個方面:(1)利用光信號提供植物光合作用,調節生長、分化和新陳代謝;(2)不同光照度和光質對植物生長、生物累積量、開花及功能性化合物的調控作用。光質是影響植物光合作用的重要因素之一,由不同輻射光譜組合而成,即使在同樣的可見光顏色下,其光譜的構成也有差異,光感受蛋白可以在特殊光譜波長范圍內促使小輔因子和發色團改變光感受器的調控,從而引起植物的生理響應[1]。不同光質對于植物不同生長發育期通過葉片光感色素接收光感信號的強度不同[2]。紅光和藍光是植物葉片光感色素吸收和利用最多和最重要的有效光源[3-4]。紅光通過刺激與調控植物光敏色素和光合器官,以此促進胚胎發育和植物化學物質合成[5]。藍光有助于葉綠體的形成、氣孔開閉以及葉綠素、花青素的生物合成[6]。研究表明,藍光可促進擬南芥幼苗莖的伸長同時降低生物量各項指標[7],紅光可促進植物莖葉生長并提高作物產量[8]。針對于紅藍光質配比的研究表明,在光質中加入紅藍配比甚至加入一些綠光可以有效促進植物生長[9]。
近年來,不同LED配比的人工光源在農業生產中推廣使用,LED光質、光照度和光周期及其對植物的影響被廣泛研究和探討。國內外學者研究了LED不同光質、光照度、光周期對番茄生長發育[10-11]、產量、品質[12]、葉綠素熒光動力學參數[13]和果實品質[14]的影響,但對于冬季日光溫室LED不同光質對番茄光合特性的影響少見報道。因此,本試驗以番茄為試驗材料,探究LED紅藍光質及其不同配比對番茄生長期、開花期、坐果期、盛果期生長指標和光合特性的影響并模擬光合速率曲線,以期為LED補光燈在冬季日光溫室栽培管理提供參考依據。
1?材料與方法
1.1?試驗設備
試驗補光材料采用深圳市一宸華節能照明有限公司訂做的LED植物生長光源,紅光與藍光比值分別為5 ∶1和7 ∶1(紅藍LED燈珠個數比),由波長為630 nm的紅色發光二極管與460 nm的藍色發光二極管組成,以自然光為對照。
1.2?試驗設計
試驗在寧夏園藝產業園科研示范基地(106°19′4.25″E、38°34′41.63″N)日光溫室大棚內進行。供試番茄品種為巴菲亞特。2017年9月20日育苗,10月20日4葉1心時定植。試驗設計為無補光、紅藍7 ∶1、紅藍5 ∶1、藍光、紅光共5個處理(表1、圖1),其中無補光作為對照(CK),具體每個處理種植5畦,3次重復,常規管理。定植后開始補光處理,隨植株生長實時調整補光燈高度,使到達葉片的光和光量子通量為80 μmol/(m2·s),各補光處理間用反光幕隔斷,每天分別在揭苫前和放苫后各補光3 h(07:00—10:00、18:00—21:00),從幼苗至拉秧持續補光。
分別于幼苗期(2017年10月29日)、生長期(2017年11月15日)、開花期(2017年11月22日)、坐果期(2017年12月20日)、盛果期(2018年1月5日)進行指標測定。
1.3?測試內容
1.3.1?生長指標的測定?每處理選取6株代表性植物掛牌標記,分別于番茄各生長階段測定株高、莖粗、葉綠素含量,葉面積參照文獻[15]計算。每個處理測定4株,每株測定3張葉片。
1.3.2?光合特性的測定?主要包括光合速率(Pn)、蒸騰速率(Tr)、胞間CO2濃度(Ci)和氣孔導度(Gs)的測定。凈光合速率的測定從 10:00 升棉被開始,整點時刻每隔2 h測定1次,至18:00降棉被結束,獲得LED不同光質補光對冬春溫室番茄不同生長階段凈光合速率日變化規律,采用Li-6400便攜式光合作用測定儀測定。番茄葉綠素熒光參數[在光適應下PSⅡ反應中心的光化學效率Y(Ⅱ)、光化學猝滅系數qP、非光化學猝滅系數NPQ、電子傳遞速率、ETR]使用便攜式光合儀測定。
1.3.3?番茄產量的測定?采收時按每小區稱質量,以3次重復的平均值代表該處理的平均產量。
1.4?數據統計與分析方法
采用DPS進行統計分析,用Excel 2007進行數據整理和作圖。
2?結果與分析
2.1?不同光質補光對番茄生長發育的影響
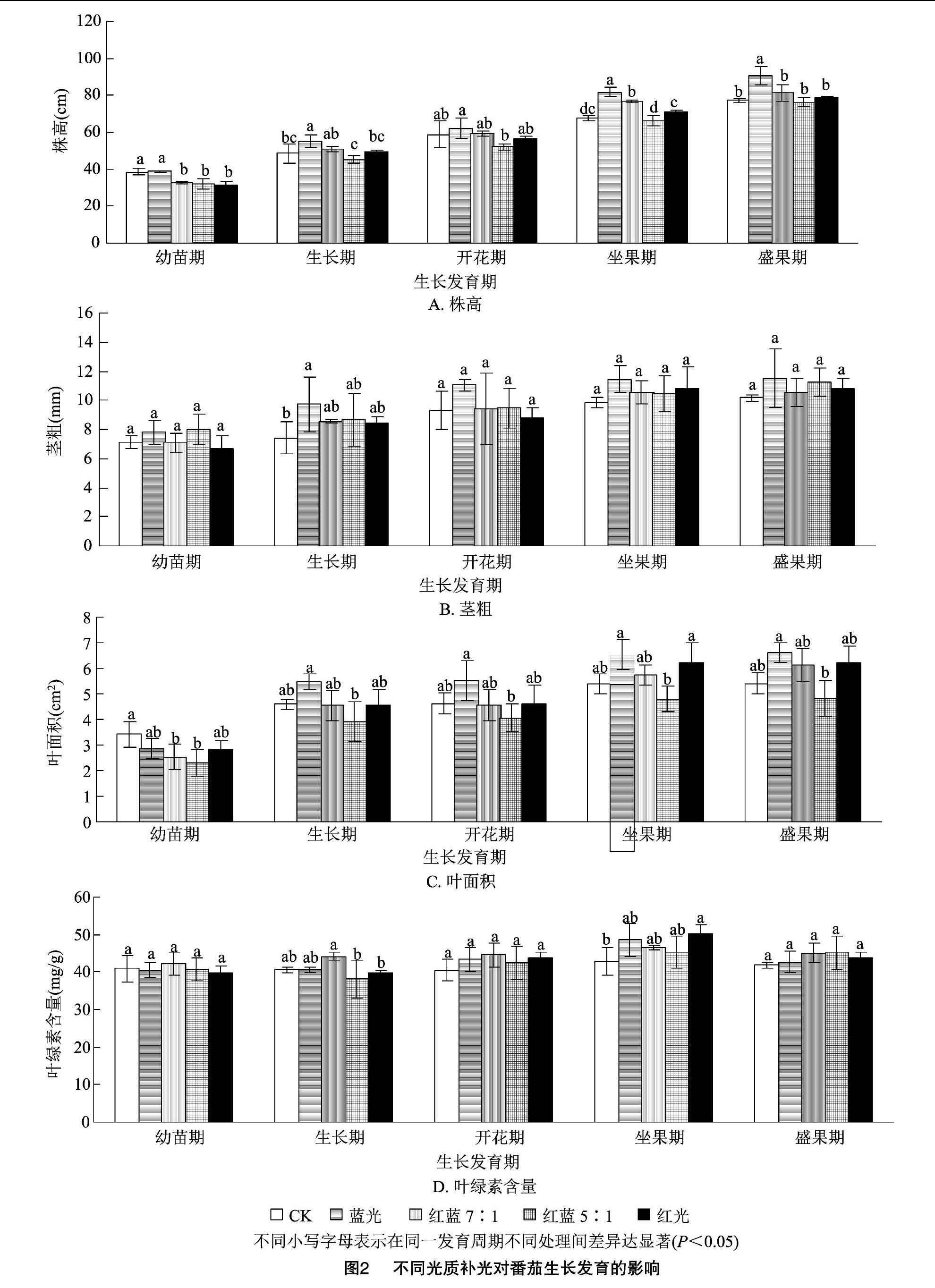
由圖2-A可見,藍光處理過的番茄株高最高,在生長期、開花期、坐果期、盛果期分別高于CK 14%、5%、21%、18%,尤其在生長期、坐果期和盛果期與其他4個處理相比基本差異達顯著(P<005)。
由圖2-B可見,藍光處理在番茄生長期的莖粗顯著高于CK(P<0.05)。在幼苗期、開花期、坐果期、盛果期各補光處理莖粗差異不顯著。
由圖2-C可見,藍光處理在生長期、開花期、坐果期、盛果期番茄葉面積高于其他處理,紅藍 5 ∶1 處理在幼苗期、生長期、開花期、坐果期、盛果期均低于其他處理。
由圖2-D可見,紅光處理葉綠素含量僅在坐果期顯著高于CK;5個處理葉綠素含量在幼苗期、開花期、盛果期差異不顯著,而在生長期、坐果期有一定差異,說明不同光質在番茄生長期、坐果期對葉綠素合成具有一定影響。
2.2?不同光質補光對番茄葉片光合特性日變化的影響
由圖3-A可知,生長期番茄葉片各光照處理下凈光合速率均大于CK,其中藍光處理呈“雙峰”值,分別在14:00和18:00達到峰值,表明藍光處理下番茄葉片光合有效時間得到延長。由圖3-B可知,各處理氣孔導度呈先下降后上升的趨勢。由圖3-C可知,各光照處理的蒸騰速率在午間12:00均達到最大值且大于CK,16:00之后均小于CK。由圖3-D可知,胞間CO2濃度呈下降趨勢。
由圖4-A可知,開花期番茄葉片各處理下凈光合速率在14:00—16:00達到峰值,表明番茄開花期凈光合速率在日間光照度最大時最為活躍。由圖4-B可知,各處理氣孔導度呈先升后降的趨勢,且在14:00—16:00達到峰值,表明番茄花期生殖生長期間須要通過光合作用積累更多的能量促進開花。由圖4-C可知,紅光處理的蒸騰速率在16:00達到峰值,其余各光質補光處理在14:00達到峰值,且14:00時所有光質處理的蒸騰速率均小于CK,表明在14:00時番茄葉片大量吸收有機物和礦物質確保其生殖生長的進行。由圖4-D可知,各處理的番茄胞間CO2濃度在12:00時明顯低于CK,表明在此期間各處理氣孔完全關閉。
由圖5-A可知,番茄葉片在各光照處理下凈光合速率均在14:00時達到最大值。由圖5-B可知,各處理氣孔導度呈先降后升的趨勢。由圖5-C可知,各光照處理蒸騰速率呈“雙峰”變化,且在 14:00 顯著低于CK,說明在14:00時光合作用最強。由圖5-D可知,番茄胞間CO2濃度呈先降后升再降的趨勢,各處理間差異不明顯。
2.3?不同光質補光對番茄凈光合速率峰值的對比分析
為了更好地比較番茄不同生長階段各光照處理凈光合速率活躍程度,選取14:00—16:00光合作用活躍階段的凈光合速率平均值進行比較,具體見圖6。其中,在番茄不同生長發育時期,各光照處理總體大于CK,表明不同LED光質處理均不同程度地增強了番茄的凈光合速率。紅光處理在開花期凈光合速率最大,藍光、紅藍5 ∶1處理在坐果期凈光合速率最大。
2.4?補光對番茄果實產量和熒光特性的影響
2.4.1?LED不同光質補光對番茄果實產量的影響?由表2可知,藍光、紅藍7 ∶1、紅藍5 ∶1、紅光處理下的番茄平均單果質量顯著高于CK,紅藍5 ∶1處理的果型指數最優。產量排序是紅光>紅藍 5 ∶1>紅藍7 ∶1>藍光>CK。
2.4.2?不同光質補光對番茄花期和果期熒光特性的影響
由圖7-A、圖7-B可見,隨不同LED光源照射時間的延長,實際光化學效率、光化學猝滅系數變化各不相同, 總體呈先降后升的趨勢。盛果后期,紅光、紅藍5 ∶1、紅藍7 ∶1光化學猝滅系數明顯高于其他處理,表明番茄盛果后期在紅光、紅藍 5 ∶1、紅藍7 ∶1處理下具有更高的實際光化學效率和更高比例還原態的電子受體質體醌A,番茄開花期、坐果期、盛果期光化學猝滅系數均無顯著差異,盛果后期各處理光化學猝滅系數差異顯著,其在 PSⅡ 反應中心開放程度依次是紅藍7 ∶1>紅藍 5 ∶1>紅 光> 藍光>CK。
由圖7-C、圖7-D可見,不同光質補光ETR和NPQ在番茄花期、果期表現均有不同,紅藍7 ∶1在盛果后期更有利于番茄的電子傳遞,且使其熱耗散水平更高。
3?結論與討論
3.1?討論
光是日光溫室內冬春番茄生長發育的重要組成部分,一方面促進植物有機物和干物質的積累,另一方面均衡調控植物營養生長、生殖生長、器官分化的周期,使能量得到有效積累和釋放。在冬季溫室低溫寡照的條件下, LED補光延長了光合速率有效時間,促進植物生理代謝,為植物的正常生長發育提供有利條件。本研究表明,番茄在不同生長階段對光質的要求不同,LED光對植物生理發育的影響與日光照射有著明顯的差異。光合作用的光源主要依靠可見光,藍光和紅光充當植物光合作用的獨立光反應者和光接收者,一些研究表明,植物體內的光接受器可能與植物的生理反應有關,而藍光和紅光在植物生理機能響應與信號傳導之間也會有干擾[16],這也充分說明該試驗不同補光處理在番茄不同生長階段和日間不同時段的生長和光合特性不同。
番茄是在果實成熟后持續采收的作物,在其果實成熟期,葉綠素含量逐漸下降,類胡蘿卜逐漸合成,尤其是番茄紅素是其果實轉色表達的重要環節[17]。植物利用光能進行光合作用,并通過調節不同光質改善植物生長周期和果實產量品質[18]。紅光和藍光在番茄采收期果實生理代謝和生物色素合成起到了至關重要的作用,紅光和藍光也影響果實大小和產量[14]。番茄花期果期補光處理下,各光照處理均不同程度對番茄產量和熒光特性產生了積極的影響。單色光譜(紅光、藍光)和混合光譜(紅藍 5 ∶1、紅藍7 ∶1)對番茄不同花期和果期各階段的作用也不同。
3.2?結論
本試驗用不同LED光質(紅光、藍光、紅藍 5 ∶1、紅藍7 ∶1)補光處理冬春季溫室番茄,研究其生長指標及光合作用。結果表明,不同LED光質處理對番茄生長特性具有不同的影響。從幼苗期到盛果期,藍光對番茄生長發育起到了至關重要的作用,藍光補光可有效促進番茄生長,且不同光質在番茄生長期、坐果期對葉綠素合成具有一定的影響。從不同LED光質處理對番茄光合特性的影響可知,番茄不同生長階段對光源補光的需求不同,各補光處理相比CK都能不同程度地提高番茄的凈光合速率。其中,生長期藍光、紅光補光處理可有效提高番茄的凈光合速率;開花期紅光、紅藍7 ∶1補光處理可有效提高番茄的凈光合速率;坐果期紅光、紅藍5 ∶1補光處理可有效提高番茄的凈光合速率;番茄生長期、開花期、坐果期補光處理在 14:00—16:00光合作用最為活躍。說明不同LED光質處理在不同生長發育時期均不同程度地提高番茄的凈光合速率,其中,紅光在開花期凈光合速率最大,藍光、紅藍5 ∶1在結果期的凈光合速率最大。番茄不同補光處理下產量排序是紅光>紅藍 5 ∶1> 紅藍7 ∶1>藍光>CK。番茄盛果后期在紅光、紅藍5 ∶1、紅藍7 ∶1處理下具有更高的實際光化學效率和更高比例還原態的電子受體質體醌A,其在PSⅡ反應中心開放程度依次是紅藍7 ∶1>紅藍5 ∶1>紅光>藍光>CK,紅藍7 ∶1在盛果后期更有利于番茄的電子傳遞。
參考文獻:
[1]Shcherbakova D M,Verkhusha V V. Chromophore chemistry of fluorescent proteins controlled by light[J]. Current Opinion in Chemical Biology,2014,20:60-68.
[2]Chen X L,Yang Q C,Song W P,et al. Growth and nutritional properties of lettuce affected by different alternating intervals of red and blue LED irradiation[J]. Scientia Horticulturae,2017,223(15):44-52.
[3]Abidi F,Girault T,Douillet O,et al. Blue light effects on rose photosynthesis and photomorphogenesis[J]. Plant Biology,2013,15(1):67-74.
[4]Massa G D,Kim H H,Wheeler R M,et al. Plant productivity in response to LED lighting[J]. HortScience,2008,43(7):1951-1956.
[5]Choi H G,Moon B Y,Kang N J. Effects of LED light on the production of strawberry during cultivation in a plastic greenhouse and in a growth chamber[J]. Scientia Horticulturae,2015,189(25):22-31.
[6]Li Q,Kubota C. Effects of supplemental light quality on growth and phytochemicals of baby leaf lettuce[J]. Environmental and Experimental Botany,2009,67(1):59-64.
[7]Folta K M. Green light stimulates early stem elongation,antagonizing light-mediated growth inhibition[J]. Plant Physiology,2004,135(3):1407-1416.
[8]Zhang Y T,Zhang Y Q,Yang Q C,et al. Overhead supplemental far-red light stimulates tomato growth under intra-canopy lighting with LEDs[J]. Journal of Integrative Agriculture,2019,18(1):62-69.
[9]Kim H H,Goins G D,Wheeler R M,et al. Stomatal conductance of lettuce grown under or exposed to different light qualities[J]. Annals of Botany,2004,94(5):691-697.
[10]何?蔚,陳丹艷,胡曉婷,等. 不同光周期與光質配比對番茄植株生長發育的影響[J]. 西北農業學報,2018,27(4):116-124.
[11]何偉明,劉龐源,武占會,等. 不同光強與光質處理下對番茄幼苗生長的影響[J]. 北方園藝,2017(19):72-75.
[12]錢舒婷. 不同補光燈對設施草莓、番茄光合生長及產量品質的影響[D]. 楊凌:西北農林科技大學,2018
[13]謝?鑫,王俊玲,段立肖,等. 光質對番茄葉片葉綠素熒光動力學參數的影響[J]. 河北農業大學學報,2013,36(6):48-51,88.
[14]Nájera C,Guil-Guerrero J L,Enríquez L J,et al. LED-enhanced dietary and organoleptic qualities in postharvest tomato fruit[J]. Postharvest Biology and Technology,2018,145:151-156.
[15]吳遠藩. 量葉片的長和寬計算番茄葉面積[J]. 農業科技通訊,1980(12):20-21.
[16]Chen X L,Yang Q C,Song W P,et al. Growth and nutritional properties of lettuce affected by different alternating intervals of red and blue LED irradiation[J]. Scientia Horticulturae,2017,223(15):44-52.
[17]Liu L H,Zabaras D,Bennett L E,et al. Effects of UV-C,red light and sun light on the carotenoid content and physical qualities of tomatoes during post-harvest storage[J]. Food Chemistry,2009,115(2):495-500.
[18]Wang Y,Folta K M. Contributions of green light to plant growth and development[J]. American Journal of Botany,2013,100(1):70-78.
