α-淀粉酶對豬糞厭氧發酵產甲烷的影響及動力學分析
2020-05-14 09:26:24肖艷春
中國沼氣 2020年6期
陳 彪, 黃 婧, 肖艷春,2
(1.福建省農業科學院農業工程技術研究所, 福建 福州 350003; 2.福建技術師范學院 近海流域環境測控治理福建省高校重點實驗室, 福建 福清 350300)
利用厭氧發酵技術處理畜禽糞便是目前世界范圍內公認的最為有效的處理方式之一。厭氧發酵技術可將生物質等轉化為沼氣,在獲取能源物質的同時,實現減少環境污染的目的[1-2]。2015年我國畜禽糞便資源總量為20.08億t,畜禽糞便資源總量約合2.87億t干物質,具有963.77億m3沼氣潛力[3]。提升厭氧發酵產沼潛力可有效促進厭氧發酵技術在污水處理領域的工程實踐應用與發展,尤其對規模化養殖污水的治理及其資源化利用具有十分重要的意義。
厭氧發酵過程可分為有機物的水解、產酸和產甲烷3個階段,而發酵原料的不同性質決定了限制階段的不同。豬糞中主要大分子復雜有機物是淀粉、蛋白質和纖維素。因此,豬糞的水解階段是厭氧發酵過程的限速步驟。已有相關研究報道,利用水解酶提高發酵原料水解產物的濃度。水解酶是一類能夠催化促進有機物水解反應的酶,包括淀粉酶、纖維素酶、蛋白酶、脂肪酶等。Yang[4]等在厭氧發酵條件下添加淀粉酶和蛋白酶提高剩余污泥的生物水解效率,淀粉酶處理比蛋白酶處理具有更高的水解效率,淀粉酶處理比對照組顯著提高了54.24%;許美蘭[5]等利用α-淀粉酶和蛋白酶構成的復合酶對豬糞進行處理,可溶性化學需氧量(SCOD)提高106.9%;王婷婷[6]等通過添加酶對釀酒廢糟進行干式厭氧發酵,提高甲烷產率達36.8%。目前關于水解酶對豬糞厭氧發酵產甲烷的影響的動力學方面的研究鮮見報道。本研究采用升流式厭氧污泥床(UASB)工藝,以豬糞為發酵原料,應用Gompertz和一級動力學模型解析外源α-淀粉酶對厭氧發酵產甲烷的影響效應,以期提升發酵原料的產甲烷潛力,提高厭氧發酵技術在養殖污水處理領域的工程實踐應用效應。
1 材料與方法
1.1 材料和裝置
鮮豬糞取自福建某養殖公司,接種污泥取自常年正常運行的沼氣發酵裝置,豬糞及接種污泥成分組成見表1。α-淀粉酶購自上海源葉生物科技有限公司,酶活≥4000 U·g-1,pH值范圍為5.5~7.5,最適溫度為50℃~70℃。試驗采用UASB工藝,裝置示意見圖1。

表1 豬糞及接種污泥的成分

1.三相分離器; 2.水封瓶; 3.氣體流量計; 4.出水口; 5.取樣口; 6.水泵; 7.蠕動泵; 8.恒溫水浴箱; 9.廢水進水池圖1 UASB發酵裝置示意
1.2 試驗方法
試驗設置1個對照組(CK組)、1個空白組和4個試驗組,每組設置3個重復,厭氧發酵周期為35 d。每組取豬糞1225 g,以未添加α-淀粉酶的處理為CK組,以只添加接種污泥與蒸餾水的處理為空白組,4個試驗組按照豬糞的總固體(TS)重量每克分別加入α-淀粉酶(酶添加量參考相關文獻[5, 7])10,20,30,40 mg,加入α-淀粉酶攪拌均勻后在50℃水浴鍋中恒溫震蕩處理4 h,充分發揮α-淀粉酶活性,待豬糞冷卻至常溫后每組接入1 L的接種污泥,經調配后發酵料液總體積為5 L(不足部分用蒸餾水補足、TS約為8%),料液通過蠕動泵導入UASB發酵裝置,進料結束后向發酵裝置中通N210 min保持厭氧環境,調節裝置溫度為35℃。厭氧發酵過程中每日定時測定沼氣產氣量、甲烷體積分數。
豬糞及接種物的TS、揮發性固體(VS)采用差重法測定;C,H,O,N,S元素的質量分數采用元素分析儀(Vario EL III,Elementar,德國)測定;沼氣日產氣量使用濕式流量計(LML-1,長春汽車濾清器有限責任公司)測定;沼氣中甲烷體積分數和總揮發性脂肪酸濃度采用氣相色譜法測定。
1.3 數據分析
1.3.1 發酵原料理論產甲烷潛力值
目前,發酵原料理論產甲烷潛力值的計算是通過分析物料中的C,H,O,N,S等元素含量進行估算,碳水化合物、蛋白質等有機物可用CxHyOzNnSs表示,通過方程式(1)估算出發酵原料的理論產甲烷潛力值Xe[8]。該研究中的甲烷產率均是基于豬糞中揮發性固體(VS)含量計算。
(1)
式中:Xe為發酵原料的理論產甲烷潛力值,mL·g-1;x,y,z,n,s分別代表了發酵原料C,H,O,N,S元素含量與C,H,O, N,S元素相對原子質量的比值(取整數)。
1.3.2 發酵底物產甲烷動力學
應用一級動力學和Gompertz模型擬合厭氧發酵過程并計算動力學反應常數,根據決定系數R2及均方根誤差RMSE對模型的擬合度進行評估。
一級動力學方程:
xt=x∞[1-exp(-kt)]
(2)
式中:X∞為整個發酵周期內豬糞甲烷產率,mL·g-1;Xt為t時刻豬糞甲烷產率,mL·g-1;k為反應速率常數,d-1。
Gompertz模型:
(3)
式中:Rm為產甲烷速率,mL·g-1·d-1;e為常數,2.718282;λ為遲滯時間,d。
1.3.3 發酵底物生物轉化產甲烷效率
以不同時刻甲烷累積產量與整個發酵周期甲烷總產量的比值表示發酵過程甲烷產率比(MPR)。以厭氧發酵不同時刻實際甲烷產率與理論甲烷產率的比值表示底物的生物產甲烷效率(BDA),計算公式:
(4)
式中:Xt為t時刻實際甲烷產率,mL·g-1;Xe為理論甲烷產率,mL·g-1。
1.3.4 數據處理
采用SPSS 19.0進行數據處理及方差分析,采用Origin 9.1軟件進行繪圖及方程擬合。各試驗組的總揮發性脂肪酸濃度、沼氣產氣量、甲烷體積分數取3次重復試驗的平均值。
2 結果與討論
2.1 α-淀粉酶對總揮發性脂肪酸的影響
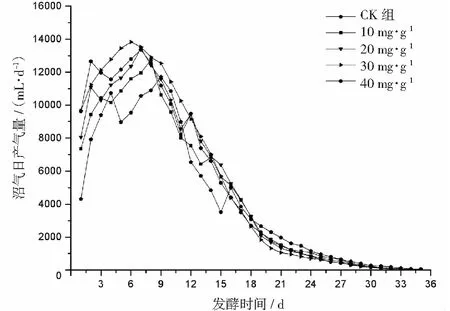
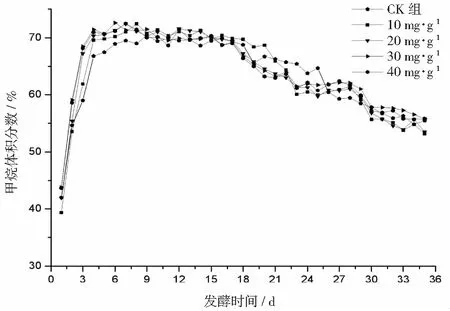
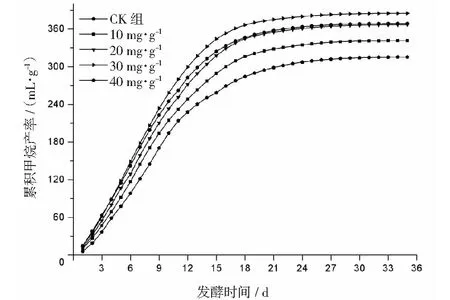
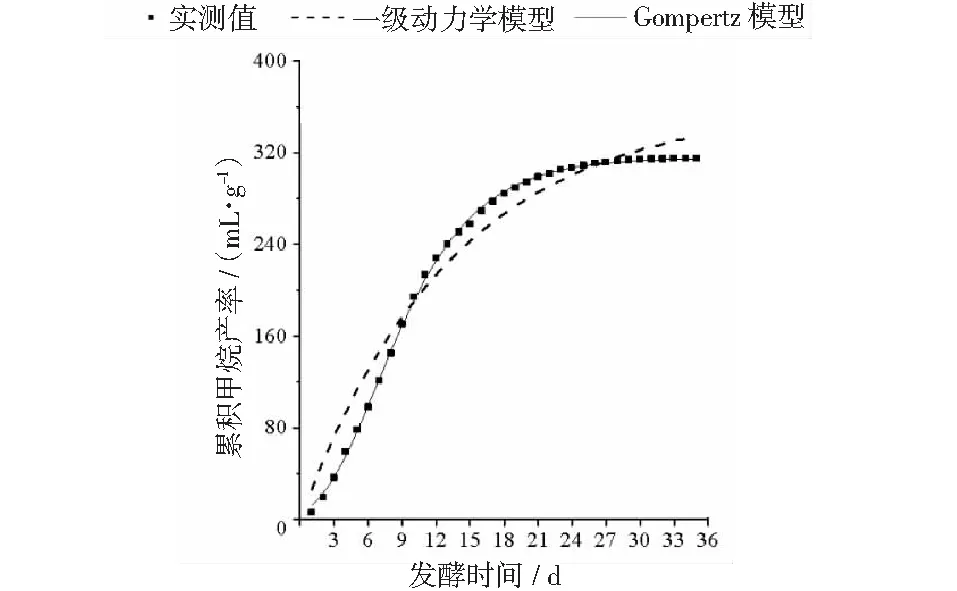
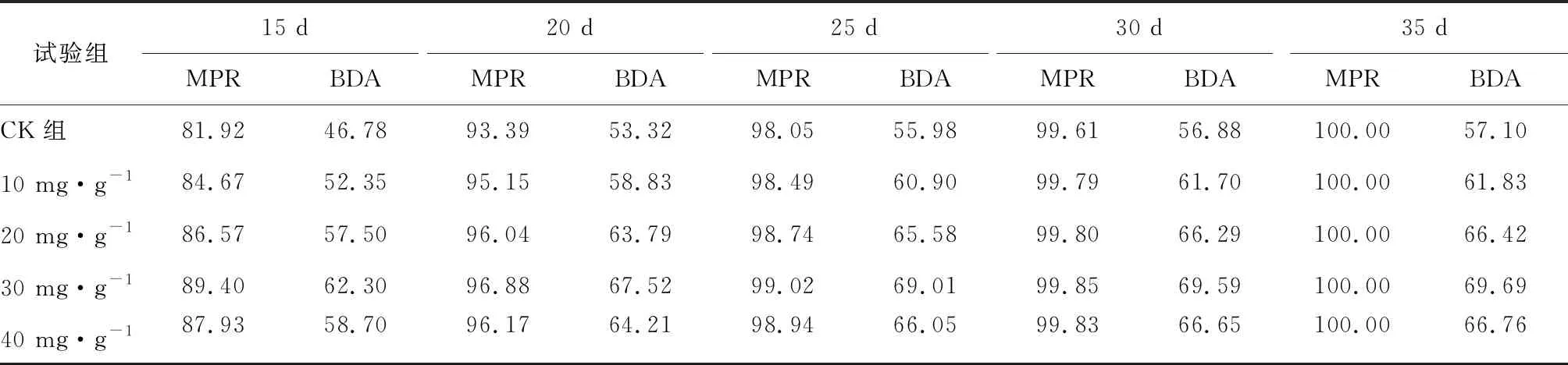
厭氧發酵過程各試驗組總揮發性脂肪酸(TVFAs)的濃度變化如圖2所示。由圖2可見:各試驗組和CK組的變化規律一致,均呈先升后降,最后趨于平穩的特征。CK組在第3天達到產酸高峰(TVFAs質量濃度為7030 mg·L-1);α-淀粉酶添加量為20,30,40 mg·g-1的試驗組均比CK組提前1天達到產酸高峰,其中添加量為40 mg·g-1的產酸峰值最大(8986 mg·L-1),比CK組提高了27.8%;其次是添加量為30 mg·g-1的試驗組(8308 mg·L-1),比CK組提高了18.2%。在厭氧發酵后期,各試驗組進入穩定期,至發酵結束時各試驗組的TVFAs累積量均低于CK組,其累積顯著度為30 mg·g-1<40 mg·g-1<20 mg·g-1<10 mg·g-1 圖2 總揮發性脂肪酸濃度變化曲線 2.2.1 α-淀粉酶對日產沼氣量的影響 各試驗組日產沼氣量變化情況見圖3,從圖3可以看出,在厭氧發酵的進程中,沼氣日產氣量總體呈現先上升后下降的趨勢。各試驗組的產氣啟動期均較短,在較短時間內就達到產氣高峰,這應與本試驗的接種污泥取自運行良好的沼氣工程厭氧發酵裝置,接種污泥中的功能微生物豐富且活躍有關。CK組,10 mg·g-1,20 mg·g-1和40 mg·g-1試驗組均出現了3個產氣高峰,CK組產氣高峰分別出現在第4,9,16天,30 mg·g-1試驗組只在第6天出現1個產氣高峰,峰值期比CK組提前3 d。日產氣峰值最大值(13829 mL)出現在30 mg·g-1試驗組,與CK組(11710 mL)相比增加了18.1%。 有研究表明[9],乙酸是沼氣發酵過程中重要的中間代謝產物。由于微生物優先利用小分子糖類及淀粉類等易降解的有機物,水解作用生成的葡萄糖在厭氧條件下經糖酵解(EMP)途徑生成丙酮酸,丙酮酸是合成乙酸等揮發性脂肪酸的主要中間產物,生成的甲烷三分之二來源于乙酸,試驗組添加的α-淀粉酶促進了溶解性多糖濃度的增加[10],它可將淀粉類物質酶解成糊精、麥芽糖、葡萄糖等這些容易被產氫產乙酸或耗氫產乙酸細菌利用的營養底物,進而為產甲烷細菌提供了大量的乙酸鹽,CO2,氫等物質,促進沼氣產氣量提高,因此添加了α-淀粉酶的試驗組比CK組更快達到產氣峰值。而后產氣速率下降,出現下1個產氣高峰,這是因為容易降解的底物被消耗后,沼氣發酵微生物不得不利用纖維素等較難降解有機物,產氣峰值不如前期。由此,α-淀粉酶的適量添加有利于產氣峰值的提前到達。 圖3 沼氣日產氣量變化曲線 2.2.2 α-淀粉酶對甲烷體積分數的影響 圖4為沼氣中甲烷體積分數變化曲線圖,該體積分數的變化是厭氧發酵過程中不同微生物菌群之間相互作用的結果。由圖4可以看出,甲烷體積分數先呈現快速增長而后趨于穩定,末期則逐步降低。在厭氧發酵的第2~3天,各試驗組甲烷體積分數就已經大于50%,均進入正常產氣狀態。CK組的甲烷體積分數從第4天起就進入穩定期,至第26天結束,共保持了23 d,甲烷體積分數60.4%~70.4%;30 mg·g-1試驗組的穩定期最長,從第3天持續到第29天,甲烷體積分數61.6%~70.6%,比CK組略有增加,試驗組添加的α-淀粉酶使厭氧發酵系統中可利用的糖類物質增加,提高了系統中甲烷途徑中碳物質濃度,促進最終產物甲烷體積分數的提升。 圖4 沼氣中甲烷體積分數變化曲線 2.2.3 α-淀粉酶對甲烷產率的影響 圖5為各試驗組累積甲烷產率曲線圖。由圖5可知,CK組的累積甲烷產率最低(315.4 mL·g-1VS)。杜連柱[11]等研究發現在35℃條件下,TS為7.5%的豬糞進行厭氧發酵時,其甲烷產率為326 mL·g-1VS,與本試驗的結果較接近。隨著α-淀粉酶添加量從10 mg·g-1增加到30 mg·g-1時,甲烷產率亦隨之增加,在添加量為30 mg·g-1時達到最高的384.9 mL·g-1VS,比CK組提高了22.1%;而當添加量繼續增至40 mg·g-1時,其對應的甲烷產率相較30 mg·g-1試驗組反而降低。由此分析,與CK組相比,在適當的范圍內添加α-淀粉酶可以促進發酵料液中的淀粉水解為小分子糖類,即丙酮酸的前體物質,產氫產乙酸細菌將丙酮酸轉化為如乙酸、丙酸、丁酸、甲醇等揮發性有機酸,反應底物的增加會提升產甲烷菌轉化速率,提高甲烷產率;但過量添加的α-淀粉酶可導致過快的有機物水解及后續過快的揮發性有機酸的產生,一旦超過產甲烷菌甲烷轉化的速率,揮發性有機酸的過量積累將抑制產甲烷菌的活性,此外,底物中部分細菌的新陳代謝活動獲取營養物質的速度高于酶促反應速度,導致可溶性物質濃度下降,使甲烷產率下降。試驗結果表明,在豬糞厭氧發酵進程中,溶解性碳源物質的濃度決定了厭氧發酵的甲烷產率,α-淀粉酶的添加閾值為30 mg·g-1。 圖5 累積甲烷產率曲線 本試驗采用經典的一級動力學以及Gompertz模型對不同試驗組的累積甲烷產率曲線進行擬合,具體見圖6~圖10。表2列出了2種模型的擬合結果,其中,產甲烷動力學參數包括動力學常數(k)、產氣速率(Rm)、發酵遲滯期時長(λ)等。根據決定系數R2及均方根誤差RMSE對模型擬合的表現進行評估,從擬合結果可以看出,采用Gompertz模型擬合各試驗組產甲烷過程,擬合度較好,R2均大于0.998,模型擬合的累積甲烷產率最大值與實測值接近,說明該模型可以較為準確的反應各試驗組的產氣情況;一級動力學模型的擬合精確度較低,擬合結果與實測值存在偏離,因此,該模型不適用于本研究中產甲烷過程擬合。 從Gompertz模型的擬合結果可以看出,CK組的Rm最慢(26.18 mL·g-1d-1),30 mg·g-1試驗組的Rm最快(32.63 mL·g-1d-1),比CK組提高24.6%。隨著α-淀粉酶的增加λ隨之縮短,CK組λ最長(2.29 d),40 mg·g-1試驗組λ較短(1.08 d);30 mg·g-1試驗組λ遲滯期次之(1.52 d)。這是由于添加的酶增加了發酵底物中小分子糖類的濃度和可吸附位點數量,促進產甲烷微生物菌群繁殖生長,進而提升甲烷產率。試驗結果表明,α-淀粉酶的添加不僅可以縮短遲滯期,而且還能提高反應速率。 圖6 CK組實測累積產甲烷及一級動力學, Gompertz模型擬合曲線 圖7 10 mg·g-1試驗組實測累積產甲烷及 一級動力學,Gompertz模型擬合曲線 圖8 20 mg·g-1試驗組實測累積產甲烷及 一級動力學,Gompertz模型擬合曲線 圖9 30 mg·g-1試驗組實測累積產甲烷及 一級動力學,Gompertz模型擬合曲線 圖10 40 mg·g-1試驗組實測累積產甲烷及 一級動力學,Gompertz模型擬合曲線 韓婭新、李超等對不同有機廢棄物的產甲烷過程進行動力學分析[8, 12-13],不同發酵物料的性質及接種比對動力學擬合模型的選擇有顯著性影響,一級動力學模型適用于擬合接種比大于3:1的厭氧發酵產甲烷過程,其他條件下適宜采用Gompertz模型進行擬合,與本研究的結果一致。 表2 擬合方程對試驗組累積甲烷產率曲線的擬合結果 表3為各試驗組厭氧發酵過程中MPR及BDA。本研究中以MPR等于90%表示厭氧發酵反應過程結束,根據該指標可以確定各試驗組的厭氧發酵水力停留時間(HRT)。CK組的HRT為17.9 d,30 mg·g-1試驗組的HRT最短(15.3 d),比CK組縮短了14.5%。在工程實踐中,HRT的適當縮短有利于降低厭氧發酵系統的建設和運行成本,提高容積甲烷產率。 分析發酵原料的組成成分,得到豬糞的化學組成為C270H485O141N18S2,根據方程式(1)計算得出其理論產甲烷潛力值為552.3 mL·g-1。BDA是衡量發酵物料轉化產甲烷是否徹底的重要指標,其值越高,說明生物質轉化的程度越高。適當添加α-淀粉酶可以提高BDA,在第15~35天,各試驗組BDA值由大到小排序為30 mg·g-1﹥40 mg·g-1﹥20 mg·g-1﹥10 mg·g-1﹥CK組。30 mg·g-1試驗組在第13天的BDA值已超過CK組第35天的值;反應結束時,與CK組相比提高了22.1%,說明α-淀粉酶的添加對BDA促進作用顯著。 表3 厭氧發酵過程部分時間點的MPR和BDA (%) (1)α-淀粉酶的適量添加有利于促進有機物厭氧發酵底物的水解,提高甲烷產率。當α-淀粉酶添加量≤30 mg·g-1時,添加量與甲烷產率增加呈正相關。 (2)α-淀粉酶添加量為30 mg·g-1的試驗組對甲烷產率的促進作用最為顯著,其日產氣峰值、甲烷產率、BDA分別為13 829 mL,384.9 mL·g-1VS,69.69%,與對照組比分別提高了18.1%,22.1%,22.1%。 (3)添加α-淀粉酶的豬糞厭氧發酵過程比較符合Gompertz模型,添加量為30 mg·g-1的試驗組最大反應速率為32.63 mL·g-1d-1,比CK組提高24.6%,決定系數R2為0.9995。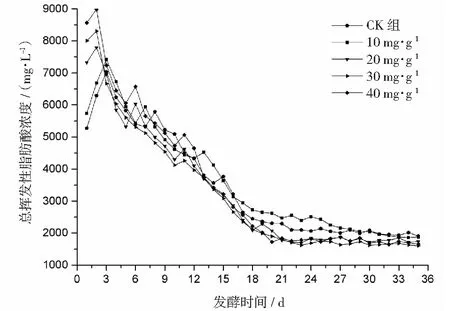
2.2 α-淀粉酶對沼氣產氣性能的影響
2.3 擬合模型的動力學分析





2.4 α-淀粉酶對BDA的影響
3 結論
