轉(zhuǎn)錄因子Dlx1在HT22細(xì)胞中調(diào)控CRMP2轉(zhuǎn)錄活性的研究
2020-05-13 14:38:28束慧
科教導(dǎo)刊·電子版 2020年3期
束慧

摘 要 目的:觀察轉(zhuǎn)錄因子Dlx1對(duì)小鼠海馬神經(jīng)元細(xì)胞HT22細(xì)胞CRMP2( Collapsin response mediator protein 2)基因的調(diào)控作用。方法: 構(gòu)建pcDNA3.1-HA-Dlx1質(zhì)粒及pGL3.0-Basic-CRMP2 promoter質(zhì)粒,采用雙熒光素酶報(bào)告基因檢測系統(tǒng)檢測Dlx1對(duì)CRMP2基因啟動(dòng)子轉(zhuǎn)錄的影響; 檢測過表達(dá)Dlx1對(duì)內(nèi)源CRMP2基因表達(dá)的影響。結(jié)果:Dlx1直接結(jié)合在CRMP2啟動(dòng)子上,促進(jìn)CRMP2基因表達(dá)。結(jié)論: Dlx1促進(jìn)CRMP2基因表達(dá)。
關(guān)鍵詞 Dlx1 腦衰反應(yīng)調(diào)節(jié)蛋白-2 啟動(dòng)子 雙熒光素酶基因檢測
中圖分類號(hào):R780.2文獻(xiàn)標(biāo)識(shí)碼:A
腦衰反應(yīng)調(diào)節(jié)蛋白2 (Collapsin response mediator protein 2, CRMP2)是CRMPs家族成員之一,均由564-572個(gè)氨基酸殘基組成。人源CRMP2基因定位于8p21.2,轉(zhuǎn)錄本Ⅱ與小鼠CRMP2基因編碼的蛋白氨基酸殘基長度均為572aa,只有兩個(gè)氨基酸不同。CRMP2表達(dá)量的下降和活性的降低均會(huì)引起神經(jīng)系統(tǒng)疾病,如帕金森疾病、精神分裂癥和阿爾茨海默癥等。
Dlx1屬于Dlx基因家族成員之一,有研究表明Dlx家族基因在神經(jīng)元發(fā)育中有很重要的作用。本實(shí)驗(yàn)通過觀察Dlx1對(duì)CRMP2基因啟動(dòng)子轉(zhuǎn)錄活性的影響,以及轉(zhuǎn)錄活性之間的關(guān)系,為進(jìn)一步研究CRMP2基因表達(dá)的調(diào)控奠定了一定基礎(chǔ)。
1材料與方法
1.1主要試劑與儀器
HT22細(xì)胞株(實(shí)驗(yàn)室保留);CRMP2質(zhì)粒構(gòu)建引物(蘇州金唯智生物科技有限公司);二抗山羊抗兔(美國Abcam,ab6721);CRMP2 siRNA(吉瑪基因股份有限公司); CRMP2抗體(美國Abcam,ab129082);Lipofectamine 2000轉(zhuǎn)染試劑(美國Invitrogen, 11668019);蛋白濃度測定試劑盒(碧云天,p0012);胎牛血清(美國Hyclone,Sh30370.03);RPMI-1640培養(yǎng)基(美國Hyclone,Sh30809.01);Ripa裂解液(康為世紀(jì), ?CW2334);DNA質(zhì)粒小提試劑盒(天根生化科技有限公司);雙熒光素酶報(bào)告基因檢測系統(tǒng)(美國 Promega 公司)。
1.2質(zhì)粒構(gòu)建
采用PCR方法擴(kuò)增得到鼠源Dlx1 cDNA片段,使用EcoR1/BamH1限制性內(nèi)切酶處理后插入pcDNA3.1-HA載體,構(gòu)建成pcDNA3.1-HA-Dlx1質(zhì)粒,PCR引物序列:上游引物序列- CCG GAATTC ATGACCATGACCACCATGCCAG,下游引物序列- GCG GGATCC TCACATCAGTTGAGGCTGCTGC。采用PCR方法擴(kuò)增得到鼠源CRMP2啟動(dòng)子片段,使用Kpn1/Mlu1限制性內(nèi)切酶處理后插入pGL3.0-Basic載體,構(gòu)建成pGL3.0-Basic-CRMP2 promoter質(zhì)粒,PCR引物序列:游引物序列-CGG GGTACCAAATATAAATATATATATATTTTAA,下游引物序列-GCACGCGT TCTCTCCGGGGGGCGGGAGGAAG。
1.3 ?Dlx1對(duì)CRMP2啟動(dòng)子(-102- +1)103bp轉(zhuǎn)錄活性的影響
1.3.1不同濃度Dlx1對(duì)CRMP2啟動(dòng)子轉(zhuǎn)錄活性的影響
HT22細(xì)胞株培養(yǎng)于RPMI-1640培養(yǎng)基(含10%胎牛血清,1%霉素/鏈霉素雙抗),置 5% CO2,37℃細(xì)胞培養(yǎng)箱培養(yǎng)。取生長狀態(tài)穩(wěn)定HT22細(xì)胞鋪板,培養(yǎng)過夜,待細(xì)胞豐度達(dá)到大約70%時(shí),按照Lipofectamine 2000轉(zhuǎn)染試劑說明書進(jìn)行轉(zhuǎn)染。將pcDNA3.1-HA載體,pcDNA3.1-HA-Dlx1質(zhì)粒分別按不同的濃度(50ng,100ng,200ng)和pGL3-Basic -CRMP2 promoter質(zhì)粒(300ng),Renilla質(zhì)粒(10ng)共轉(zhuǎn)染到12孔板 HT22細(xì)胞,各轉(zhuǎn)染3個(gè)孔。48 h后收集細(xì)胞,分別使用100L 細(xì)胞裂解液裂解細(xì)胞,充分裂解,離心后取20L上清液用于雙熒光素酶報(bào)告基因試劑盒檢測,具體步驟參考說明書。
1.3.2小鼠CRMP2基因啟動(dòng)子特征性序列的定點(diǎn)突變
將小鼠CRMP2啟動(dòng)子序列進(jìn)行轉(zhuǎn)錄因子結(jié)合位點(diǎn)分析(http://jaspar. genereg.net/cgi-bin/jaspar_db.pl),結(jié)果顯示轉(zhuǎn)錄因子Dlx1在預(yù)測得到的CRMP2啟動(dòng)子區(qū)域具有潛在結(jié)合位點(diǎn)(tataaatata)。設(shè)計(jì)突變引物,將tataaatata序列突變掉構(gòu)建突變質(zhì)粒,將得到的突變質(zhì)粒送至蘇州金唯智生物科技有限公司測序,將所得結(jié)果使用NCBI。
1.3.3檢測Dlx1對(duì)CRMP2野生型和突變型基因啟動(dòng)子轉(zhuǎn)錄活性的影響
將pcDNA3.1-HA載體,pcDNA3.1-HA-Dlx1質(zhì)粒分別和pGL3.0-Basic -CRMP2 promoter質(zhì)粒及突變質(zhì)粒,Renilla質(zhì)粒共轉(zhuǎn)染到12孔板HT22細(xì)胞,各轉(zhuǎn)染3個(gè)孔。按照Lipofectamine 2000轉(zhuǎn)染試劑說明書進(jìn)行轉(zhuǎn)染。將pcDNA3.1-HA-Dlx1質(zhì)粒按不同的濃度(50ng,100ng,200ng)分別和pGL3.0-Basic -CRMP2 promoter質(zhì)粒/突變質(zhì)粒(300ng),Renilla質(zhì)粒(10ng)共轉(zhuǎn)染到12孔板 HT22細(xì)胞,各轉(zhuǎn)染3個(gè)復(fù)孔。48 h后收集細(xì)胞,分別使用100L 細(xì)胞裂解液裂解細(xì)胞,充分裂解,離心后取20L上清液用于雙熒光素酶報(bào)告基因試劑盒檢測,具體步驟參考說明書。
1.4 ?Dlx1促進(jìn)內(nèi)源性CRMP2基因表達(dá)
將pcDNA3.1-HA載體和pcDNA3.1-HA-Dlx1質(zhì)粒分別轉(zhuǎn)染到12孔板HT22細(xì)胞中,每孔轉(zhuǎn)染500ng,各轉(zhuǎn)染6個(gè)復(fù)孔;在HT22細(xì)胞中分別轉(zhuǎn)染10ul NC siRNA和siRNA CRMP2(濃度均為1nM)。48h后,提取mRNA和蛋白質(zhì),分別做Q-PCR和Western Blot檢測CRMP2的mRNA和蛋白變化。Q-PCR引物序列:上游引物序列-GGGATCTGATGCTGACTTGGTC,下游引物序列-GGGAATGTAGCGTCCTGAGC。將收集的蛋白質(zhì)采用BCA試劑盒定量后,濃度調(diào)至一致,加入5咨涎撼逡海?5 ℃煮沸15 min。將30ug蛋白樣品加到10%聚丙烯酰胺凝膠電泳(SDS-PAGE)凝膠上樣孔中,80V電泳20min,120V電泳120min,使用PVDF膜轉(zhuǎn)膜 (恒定電流:330 mA,70min)。室溫下5%的小牛血清封閉1 h,孵育CRMP2及GAPDH抗體(1:5000)室溫2h,TBST洗膜4次,每次5 min,孵育山羊抗兔二抗(1:10000)室溫下孵育1 h,洗膜4次,每次5 min,ECL發(fā)光液曝光收集結(jié)果。
1.5統(tǒng)計(jì)學(xué)處理
實(shí)驗(yàn)數(shù)據(jù)采用單因素方差分析和雙因素方差分析來進(jìn)行顯著性分析,p<0.05 表示差異具有統(tǒng)計(jì)學(xué)意義,用Graph Pad Prism進(jìn)行統(tǒng)計(jì)學(xué)分析,數(shù)據(jù)均以均值北曜嘉蟊硎盡?
2結(jié)果
2.1不同濃度Dlx1對(duì)CRMP2啟動(dòng)子轉(zhuǎn)錄活性的影響
檢測不同濃度的轉(zhuǎn)錄因子Dlx1質(zhì)粒與pGL3.0-Basic -CRMP2 promoter質(zhì)粒共轉(zhuǎn)染HT22細(xì)胞時(shí)發(fā)現(xiàn),隨著Dlx1濃度的增高,pGL3.0-Basic -CRMP2 promoter質(zhì)粒的熒光素酶活性顯著增高,差異有統(tǒng)計(jì)學(xué)意義( P <0.001) ,結(jié)果如圖1所示。
圖1:pcDNA3.1-HA載體,不同濃度的轉(zhuǎn)錄因子Dlx1質(zhì)粒分別與pGL3.0-Basic -CRMP2 promoter質(zhì)粒共轉(zhuǎn)染HT22細(xì)胞,以Renilla為內(nèi)參,使用雙熒光素酶報(bào)告基因試劑盒檢測熒光酶的活性,結(jié)果顯示,隨著轉(zhuǎn)錄因子Dlx1濃度的增高,熒光素酶的活性也隨之增強(qiáng),p<0.001。
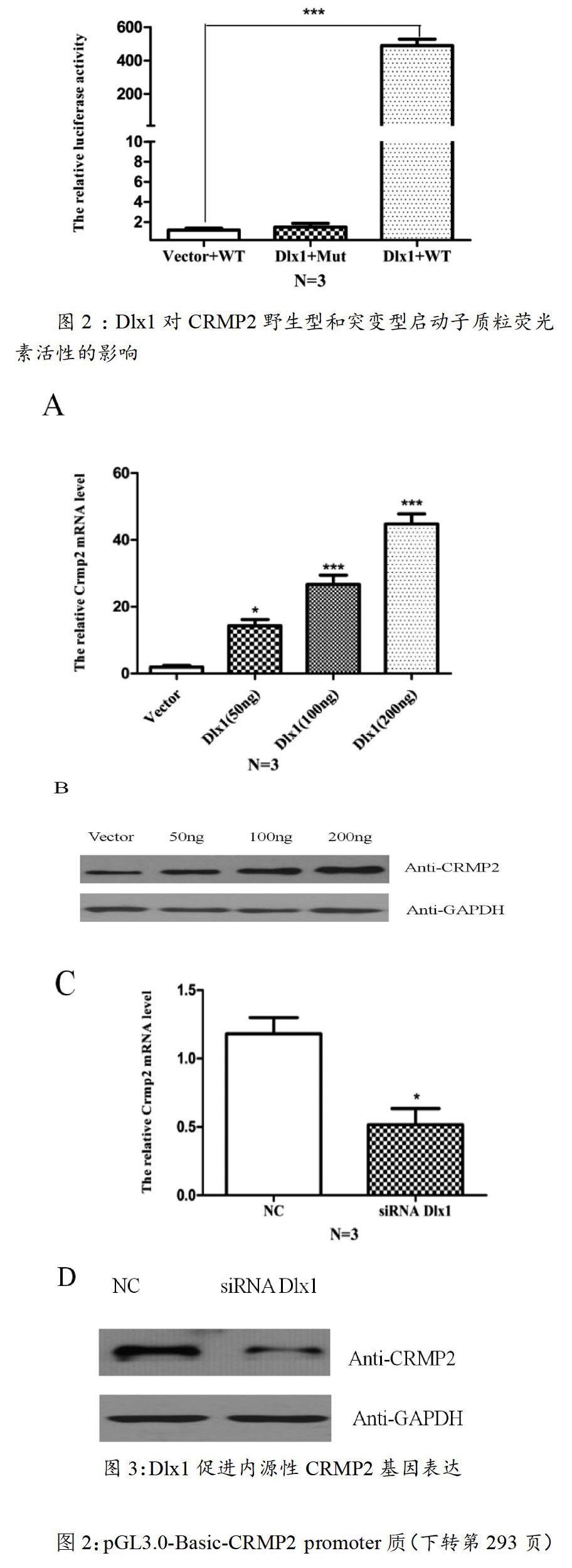
2.2 ?Dlx1對(duì)CRMP2野生型和突變型啟動(dòng)子質(zhì)粒熒光素活性的影響
通過生物信息學(xué)的分析,我們將Dlx1在預(yù)測得到的CRMP2啟動(dòng)子區(qū)域具有潛在結(jié)合位點(diǎn),通過PCR定點(diǎn)突變的方法去掉,構(gòu)建突變質(zhì)粒。然后將Dlx1分別與CRMP2野生型和突變型啟動(dòng)子質(zhì)粒轉(zhuǎn)染,檢測熒光素酶的活性。Dlx1對(duì)野生型CRMP2啟動(dòng)子質(zhì)粒熒光素活性有激活作用,差異有統(tǒng)計(jì)學(xué)意義(p<0.001);對(duì)CRMP2突變型啟動(dòng)子質(zhì)粒熒光素活性無明顯激活作用,差異無統(tǒng)計(jì)學(xué)意義(p>0.05),結(jié)果如圖2所示。
圖2:pGL3.0-Basic-CRMP2 promoter質(zhì)(下轉(zhuǎn)第293頁)(上接第288頁)粒中的Dlx1結(jié)合位點(diǎn)突變后,Dlx1不能促進(jìn)光素酶的活性增強(qiáng),差異無統(tǒng)計(jì)學(xué)意義(p>0.05),說明Dlx1可以直接結(jié)合在CRMP2啟動(dòng)子,并促進(jìn)其轉(zhuǎn)錄。
2.3 ?Dlx1促進(jìn)內(nèi)源性CRMP2基因表達(dá)
將不同濃度pcDNA3.1-HA-Dlx1質(zhì)粒瞬時(shí)轉(zhuǎn)染到HT22細(xì)胞,采用Q-PCR和Western Blot方法檢測CRMP2 mRNA和蛋白水平的變化,CRMP2 mRNA和蛋白水平均升高,有統(tǒng)計(jì)學(xué)意義(p<0.05,p<0.001),結(jié)果如圖3-A和3-B所示;在HT22細(xì)胞中Knock-down Dlx1,CRMP2 mRNA和蛋白水平均下降,有統(tǒng)計(jì)學(xué)意義(p<0.05)結(jié)果如圖3-C和3D所示,說明Dlx1可以促進(jìn)CRMP2基因表達(dá)。
圖3-A:不同濃度過表達(dá)Dlx1組與對(duì)照組相比,CRMP2 mRNA水平顯著升高,差異有統(tǒng)計(jì)學(xué)意義(p<0.05,p<0.001);圖3-B:不同濃度過表達(dá)Dlx1組與對(duì)照組相比,CRMP2 蛋白水平顯著升高,差異有統(tǒng)計(jì)學(xué)意義(p<0.05);圖3-C:Knock-down Dlx1組與對(duì)照組相比,CRMP2 mRNA水平下降,差異有統(tǒng)計(jì)學(xué)意義(p<0.05);圖3-D:Knock-down Dlx1組與對(duì)照組相比,CRMP2 蛋白水平下降,差異有統(tǒng)計(jì)學(xué)意義(p<0.05)。
3討論
近年來,蛋白質(zhì)組學(xué)已得到廣泛應(yīng)用,為一些疾病發(fā)病機(jī)制的研究帶來了更寬廣的思路。諸多實(shí)驗(yàn)已證明,CRMP2在神經(jīng)系統(tǒng)疾病中起到重要作用,其表達(dá)貫穿神經(jīng)元生命始終,發(fā)育階段達(dá)到高峰。同時(shí)CRMP2的表達(dá)量和神經(jīng)元位置也有相關(guān)性,例如,在小腦、海馬和嗅球等處CRMP2 表達(dá)較多。
本實(shí)驗(yàn)通過采用生物信息學(xué)的方法,預(yù)測出轉(zhuǎn)錄因子Dlx1可能調(diào)控CRMP2的基因表達(dá),并通過相關(guān)實(shí)驗(yàn)加以驗(yàn)證,首次發(fā)現(xiàn),轉(zhuǎn)錄因子可以直接結(jié)合在CRMP2啟動(dòng)子上,并促進(jìn)CRMP2基因表達(dá)。基因轉(zhuǎn)錄過程十分復(fù)雜,通常情況下,許多轉(zhuǎn)錄因子形成復(fù)合物共同起調(diào)控作用,所以我們不能排除還有其他轉(zhuǎn)錄因子也參與這一調(diào)控過程,具體還有哪些轉(zhuǎn)錄因子參與調(diào)控有待我們進(jìn)一步研究探索。
綜上所述,轉(zhuǎn)錄因子Dlx1與CRMP2啟動(dòng)子相互結(jié)合從而調(diào)控其表達(dá)水平,為一些疾病發(fā)病研究和治療提供一定的理論支持。
