旋毛蟲肌幼蟲期外泌體的分離和小RNA鑒定
2020-05-08 00:47:58劉曉雷劉明遠
中國人獸共患病學報 2020年4期
高 欣,楊 勇,劉 蕾,劉曉雷,劉明遠,白 雪
2014年,世界衛(wèi)生組織(WHO)和聯(lián)合國糧農組織(FAO)根據(jù)寄生蟲對人體造成的損傷以及經(jīng)濟損失的綜合評估,將旋毛蟲列為食源性寄生蟲之首[1]。自Page和Owen于1835年首次在人體尸檢的肌肉中發(fā)現(xiàn),但到目前為止,旋毛蟲的具體感染機制仍不清楚[2]。研究表明,旋毛蟲蟲體粗提物和排泄分泌產物能夠誘導宿主強烈的Th2型免疫反應,引起宿主免疫抑制,最終建立慢性感染從而達到長期寄生[3]。大量實驗證明旋毛蟲蟲體粗提物或排泄分泌產物中存在可調控宿主免疫細胞、參與旋毛蟲與宿主的相互作用的關鍵分子[4]。外泌體(exosomes, EXO)是一種表面為磷脂膜的分泌小泡,幾乎所有細胞都能產生,存在于生物體液中,包括腹水、膽汁、母乳等[5]。EXO攜帶多種生物活性物質,如蛋白質、miRNA和mRNA,EXO與靶細胞融合后,其攜帶的蛋白質和RNA轉移到靶細胞,進而影響靶細胞的功能[6]。
研究表明,包括肝片吸蟲、蛔蟲、棘球絳蟲等在內的很多寄生蟲可以分泌外泌體作用于宿主細胞,調控宿主的基因表達和免疫反應,參與寄生蟲致病過程[7]。2018年,Li Y等[8]發(fā)現(xiàn)弓形蟲釋放的EXO能夠激活巨噬細胞,刺激促炎因子分泌,觸發(fā)宿主免疫反應,對弓形蟲感染進行局部保護。Fromm B等[9]發(fā)現(xiàn)肝片吸蟲釋放的EXO含有10個與宿主同源的miRNA,提示EXO可能干預宿主的轉錄機制。Eline P. Hansen等[10]發(fā)現(xiàn)蛔蟲EXO中的miRNA可以靶向宿主體內編碼免疫反應相關的細胞因子,從而調控宿主免疫反應。因此,EXO在細胞間通過傳遞效應分子進而介導寄生蟲與宿主之間的相互作用,這些研究提示,旋毛蟲也可能分泌EXO調節(jié)宿主免疫反應,從而成功寄生[11]。
本研究采用超速離心法從旋毛蟲肌幼蟲培養(yǎng)上清中成功獲得EXO,并應用透射電子顯微鏡、納米顆粒跟蹤分析、免疫印跡以及小RNA測序對所獲的EXO進行鑒定,為今后深入研究旋毛蟲肌幼蟲期EXO(ML-EXO)參與炎癥及其他生物學過程提供基礎,并為旋毛蟲病的致病機制及靶向治療提供相關依據(jù)。
1 材料與方法
1.1 材 料
1.1.1實驗動物與旋毛蟲蟲種 Wistar雌性大鼠8只,體重180~200 g(吉林大學醫(yī)學部動物中心),用于旋毛蟲傳代保種及旋毛蟲肌幼蟲期外泌體的收集。中國河南豬旋毛蟲分離株T.spiralis(ISS534)由本室傳代保種。
1.1.2主要試劑 胃蛋白酶、鹽酸均購于中國惠世生化試劑有限公司,RPMI-1640培養(yǎng)基、10 000 U/mL青霉素、10 000 U/mL鏈霉素、胎牛血清均購于Gibco公司,戊二醛、磷鎢酸均購于無錫市江原實業(yè)技貿總公司,BCA試劑盒購于上海碧云天生物技術有限公司,兔抗鼠CD63多克隆抗體、山羊抗鼠Enolase多克隆抗體均購于Abcam公司,辣根過氧化物酶標記的山羊抗兔IgG、驢抗山羊IgG均購于Cell Signaling公司,Tru Seq小RNA文庫制備試劑盒購于Illumina公司。
1.2 方 法
1.2.1旋毛蟲肌幼蟲的收集 取8只Wistar大鼠,經(jīng)口感染3 500條旋毛蟲肌幼蟲,于感染后35 d(dpi)脫頸處死,去其皮膚、脂肪、四肢、內臟等。在顯微鏡下觀察膈肌,確定有旋毛蟲肌幼蟲后,將胴體和膈肌絞碎,放入人工消化液(10 mL/L濃鹽酸和10 g/L胃蛋白酶)中,在37 ℃下恒溫消化2 h。液體過篩網(wǎng)并自然沉淀1 h,加入清水洗滌直至液體清亮,收集旋毛蟲肌幼蟲[12]。
1.2.2旋毛蟲肌幼蟲期外泌體的提取 本實驗采用超速離心法提取旋毛蟲肌幼蟲期EXO(ML-EXO)。將旋毛蟲肌幼蟲用含5%雙抗(10 000 U/mL青霉素、10 000 U/mL鏈霉素)的無血清RPMI-1640培養(yǎng)基清洗5次后,置于37 ℃、5% CO2培養(yǎng)箱中用800 mL含2%雙抗的RPMI-1640培養(yǎng)基培養(yǎng)12 h。收集旋毛蟲肌幼蟲培養(yǎng)液,分裝于50 mL離心管中,首先于4 ℃下800 g離心15 min,去除蟲體。取上清置于新的離心管中,于4 ℃下5 000 g離心15 min以去除雜質。收集上清,使用0.22 μm過濾器過濾后轉移到10 kDa超濾管中進行超濾,4 ℃下5 000 g離心15 min。所得上清于4 ℃下120 000 g離心2 h,棄去上清,沉淀即為ML-EXO[13]。100 μL PBS重懸EXO,—80 ℃保存,以備后續(xù)使用。
1.2.3透射電子顯微鏡觀察旋毛蟲肌幼蟲期外泌體形態(tài) 采用透射電子顯微鏡的方法對ML-EXO形態(tài)進行檢測。將純化后的EXO在PBS中重懸后,取10 μL ML-EXO懸浮液置于銅網(wǎng),靜置1 min后,用濾紙將液體吸去,采用3%磷鎢酸鈉溶液室溫負染2 min,室溫風干,置于透射電子顯微鏡(Hitachi H-7650)下觀察ML-EXO形態(tài),在80 kV下調節(jié)亮度及焦距進行圖像采集[14]。
1.2.4納米顆粒跟蹤分析法檢測旋毛蟲肌幼蟲期外泌體大小 將ML-EXO沉淀用1 mL PBS重懸制成1 mL 0.5 g/L的溶液,經(jīng)0.22 μm濾器過濾后置于冰上。按照納米顆粒跟蹤分析儀(Nanno Sight NS3000)操作流程,調整參數(shù),檢測EXO的粒子數(shù)目、濃度以及粒徑分布,收集并保存數(shù)據(jù)[15]。
1.2.5蛋白免疫印跡法檢測旋毛蟲肌幼蟲外泌體的CD63和Enolase蛋白水平 通過BCA測定ML-EXO濃度,將其充分裂解,進行10% SDS-PAGE電泳分離蛋白質,每孔上樣30 μg蛋白質。電泳結束后轉膜,5%脫脂奶粉封閉。然后加入兔抗鼠CD63(1∶1 000)、山羊抗鼠Enolase多克隆抗體(1∶200),4 ℃孵育過夜,使用1×TBST緩沖液洗3次。再分別加入辣根過氧化物酶標記的山羊抗兔IgG(1∶2 000)、驢抗山羊IgG(1∶50 000),室溫孵育2 h,使用1×TBST緩沖液洗3次,加入化學發(fā)光顯色液,應用凝膠成像分析進行分析[16]。
1.2.6旋毛蟲肌幼蟲外泌體中小RNA的高通量測序 采用Trizol方法從ML-EXO中提取總RNA,采用Agilent 2100系統(tǒng)分析RNA質量[17]。使用15%聚丙烯酰胺凝膠電泳(PAGE)分離18~30 nt之間的小RNA,使用Tru Seq小RNA文庫制備試劑盒構建小RNA文庫。經(jīng)檢測合格后的小RNA,在Illumina HiSeq2000測序儀上進行測序分析。本研究的小RNA文庫構建和測序均由深圳華大基因公司完成。
對原始測序數(shù)據(jù),首先去除測序質量較低、不確定堿基大于10%、5′接頭污染、無插入片段、無3′接頭、包含polyA、小于18 nt的片段,得到clean reads。通過blast或bowtie將clean reads和miRBase、GenBank、Rfam數(shù)據(jù)庫比對,鑒定出與數(shù)據(jù)庫中完全匹配的小RNA。將測序后的小RNA進行分類注釋,包括rRNA、snRNA、tRNA、miRNA等。對于miRNA的分析,通過targetscan和miRanda軟件,預測miRNA的靶基因,應用GO功能和ggplot2分析研究靶基因的生物學功能。
2 結 果
2.1旋毛蟲肌幼蟲期外泌體形態(tài)學觀察 將超速離心法分離純化獲得的ML-EXO,置于40 000倍透射電子顯微鏡下觀察,可見EXO呈典型的橢圓形或圓形的杯狀形態(tài),具有明顯的膜結構,其腔內有低密度顆粒,背景清晰,無污染,符合EXO在透射電子顯微鏡下的特征(圖1)。利用納米顆粒跟蹤分析法對所提取ML-EXO的粒徑進行檢測,做出分析報告(表1)。其中直徑80~200 nm約占83%,200~300 nm約占12%,300~500 nm約占5%,可以看出ML-EXO大多數(shù)處于80~200 nm直徑范圍。
2.2Western blotting檢測旋毛蟲肌幼蟲期外泌體特異性標記物 采用免疫印記法,對旋毛蟲肌幼蟲及其EXO進行檢測,結果顯示旋毛蟲肌幼蟲及其EXO均表達特異性標記蛋白CD63和Enolase,大小分別是43 kDa和49 kDa(圖2)。

圖1 透射電子顯微鏡觀察旋毛蟲肌幼蟲期外泌體的形態(tài)特征Fig.1 Morphological characterization of T. spiralis muscle larvae exosomes under TEM
表1 旋毛蟲肌幼蟲期外泌體不同大小的百分比
Tab.1 Percentage ofT.spiralismuscle larvae EXO in various size range
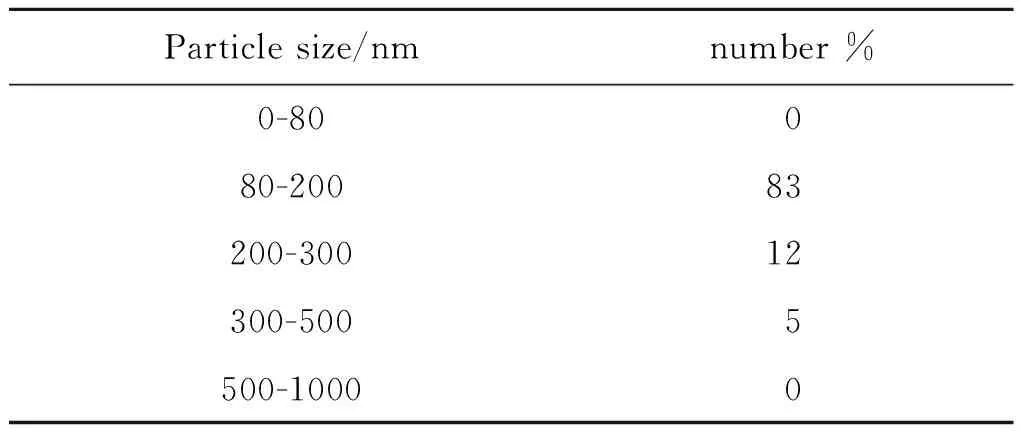
Particle size/nmnumber %0-80080-20083200-30012300-5005500-10000
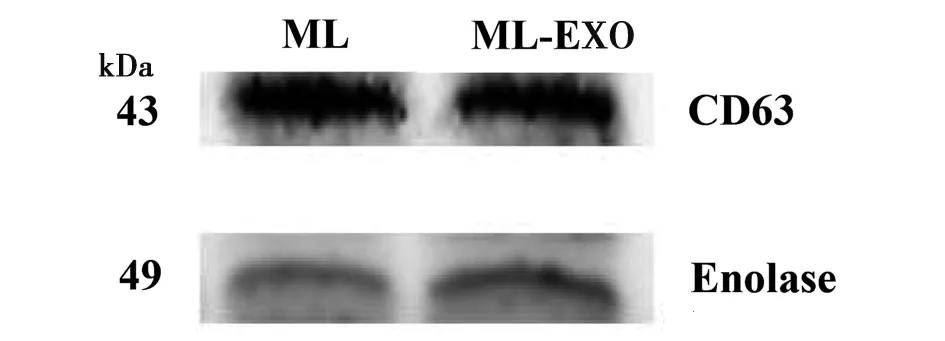
圖2 蛋白質免疫印跡法檢測旋毛蟲肌幼蟲期外泌體特異性標記蛋白CD63和Enolase蛋白表達Fig.2 Expression of specific markers CD63 and Enolase in T. spiralis muscle larvae EXO by western blotting
2.3旋毛蟲肌幼蟲期外泌體的小RNA分析 將所有ML-EXO中的小RNA與各類RNA的比對、注釋情況進行總結。結果發(fā)現(xiàn),在ML-EXO小RNA文庫中,來源于編碼蛋白質的內含子和外顯子共6 764種,約占小RNA的0.55%,表明總RNA質量完好,基本沒有降解。各類非編碼小RNA共206 120種,占小RNA的16.64%,其中rRNA的比例最高,占12.80%。未注釋的小RNA共959 200種,占小RNA的77.45%,這部分序列可能是新小RNA序列,其分類和功能值得進一步研究(圖3A)。通過將小RNA和miRBase數(shù)據(jù)庫比對,鑒定出已知miRNA共66 352種,占小RNA的5.36%。對已知miRNA用多個軟件進行靶基因預測,發(fā)現(xiàn)靶基因數(shù)量為34 437個(表2)。為了確定預測的靶基因行使的主要生物學功能,本研究通過GO富集分析靶基因的功能分類體系,發(fā)現(xiàn)在細胞、生物學調節(jié)、代謝等過程中,靶基因顯著富集(圖3B)。進一步利用ggplot2對豐度前50的miRNA的靶基因進行功能預測,發(fā)現(xiàn)其參與了多種免疫調節(jié)如MAPK級聯(lián)反應、JNK級聯(lián)反應和B細胞活化等過程(圖3C)。

A為小RNA的種類;B為miRNA的GO功能分析;C為miRNA對免疫相關靶基因的調節(jié)分析圖3 旋毛蟲肌幼蟲期外泌體中的小RNA分析Fig.3 Small RNA analysis in T. spiralis muscle larvae EXO
表2 旋毛蟲肌幼蟲期外泌體中的miRNA靶基因預測統(tǒng)計
Tab.2 Summary ofT.spiralismuscle larvae EXO miRNA target prediction
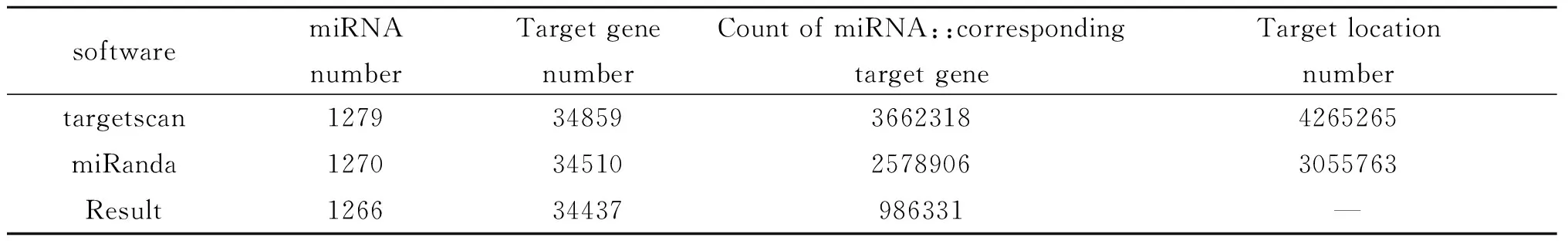
softwaremiRNA numberTarget gene numberCount of miRNA::corresponding target geneTarget location numbertargetscan12793485936623184265265miRanda12703451025789063055763Result126634437986331—
3 討 論
2013年,Rothman、Schekman和Sudhof對外泌體(EXO)運輸調節(jié)機制的研究獲得諾貝爾生理或醫(yī)學獎。自此,EXO成為研究熱點[18]。旋毛蟲是一種重要的食源性寄生蟲,嚴重危害公共衛(wèi)生安全,但關于旋毛蟲肌幼蟲期EXO的研究尚未報道[19]。目前分離EXO的方法很多,常用的有超速離心法、蔗糖密度梯度離心法、超濾法等[20]。超速離心法是分離EXO的常用方法,已被廣泛應用于各種EXO的分離,主要依據(jù)粒子密度和大小設置離心力,一般為高速離心和低速離心配合,可獲得較高純度的EXO,具有操作簡單、不易污染等優(yōu)點,因此是EXO提取的金標準[21]。我們通過超速離心法分離旋毛蟲肌幼蟲培養(yǎng)12h后的上清,在透射電子顯微鏡觀察發(fā)現(xiàn),旋毛蟲肌幼蟲期外泌體(ML-EXO)呈球形,與EXO的經(jīng)典形態(tài)相似。納米顆粒追蹤技術原理是對每個顆粒的布朗運動進行跟蹤和分析,結合Stockes-Einstein方程式計算出納米顆粒的流體力學直徑和濃度,該技術的樣本處理簡單,能保證EXO原始狀態(tài),檢測速度快,已成為鑒定EXO的方法之一[22]。采用納米顆粒追蹤技術檢測到ML-EXO直徑在80~200 nm之間。EXO的標記物有CD9、CD63、CD81、Enolase等,其中CD63是一種保守的四聚體跨膜蛋白,直接參與了EXO內容物的分選,Enolase則參與EXO的合成過程[23]。因此,本研究檢測了ML-EXO CD63和Enolase的表達,均為陽性。這些結果表明我們成功分離獲得了旋毛蟲肌幼蟲期EXO。
miRNA可以通過EXO從寄生蟲傳遞到宿主細胞,從而調節(jié)靶基因的表達,參與寄生蟲與宿主的相互作用,因此,鑒定EXO中的miRNA為研究寄生蟲病的發(fā)病機制和藥物靶點開辟了新的途徑[24]。Let-7是蠕蟲中發(fā)現(xiàn)的第一個miRNA,同時也出現(xiàn)在寄生蟲EXO中,可下調宿主的免疫反應,miRNA還與寄生蟲的生長發(fā)育有關,miR-277和miR-4989可通過介導信號通路參與血吸蟲的發(fā)育[25]。本研究通過targetscan和miRanda軟件,在ML-EXO中鑒定了1 266種miRNA。研究miRNA功能的前提是確定其靶基因,然而,由于互補的序列較少,使得對miRNA靶基因預測的準確性較低[26]。但是對miRNA與靶基因的研究發(fā)現(xiàn)它們的作用有一定規(guī)律,預測一個靶基因的決定因素為miRNA的5′端有6~8個核苷酸可以與靶基因mRNA的3′端精確互補,這6~8個核苷酸稱為“種子序列”[27]。因此,miRNA靶基因預測主要用種子序列的互補程度和miRNA與相應靶基因結合的穩(wěn)定性作為基礎[28]。基于這些原則,人們開發(fā)miRNA靶基因預測軟件有Targetscan、miRanda等[29]。Targetscan軟件基于miRNA的種子序列與靶基因序列互補,而miRanda軟件基于miRNA與相應靶基因結合的穩(wěn)定性。因此,這些軟件的算法和側重點有區(qū)別,對預測同一個miRNA靶基因也有偏差[30]。所以,我們用Targetscan和miRanda軟件進行預測,取交集作為預測結果,結果表明,miRNA的靶基因數(shù)量有34 437個,靶基因預測結果還需進一步驗證。在miRNA靶基因的GO富集分析中,ML-EXO中miRNA的靶基因富集到細胞過程、生物學調節(jié)、代謝過程等過程中,這表明EXO中miRNA的靶基因在細胞過程、生物學調節(jié)、代謝過程中發(fā)揮重要作用。寄生蟲對宿主的免疫調節(jié)作用是通過寄生蟲和宿主之間遺傳信息的傳遞形成的,因此,寄生蟲miRNA在調節(jié)宿主免疫系統(tǒng)中發(fā)揮了重要作用,我們通過ggplot2分析,發(fā)現(xiàn)一些miRNA調節(jié)免疫相關的靶基因,可能是旋毛蟲逃避宿主免疫防御的重要機制,這將為旋毛蟲與宿主的相互作用提供新的思路。
綜上所述,本研究首次證實了旋毛蟲肌幼蟲可分泌EXO,同時,證明了EXO中含有多種功能性小RNA,為旋毛蟲EXO分離提供實驗依據(jù),也為后續(xù)ML-EXO發(fā)揮免疫調節(jié)作用等相關研究提供基礎。
利益沖突:無
