葉片光合生理參數變化特征與小麥受旱狀態的關系
2020-05-06 01:14:22趙福年王潤元王鶴齡
干旱地區農業研究 2020年1期
關鍵詞:模型
趙福年,王潤元,張 凱,齊 月,陳 斐,趙 鴻,王鶴齡
(中國氣象局蘭州干旱氣象研究所,甘肅省干旱氣候變化與減災重點實驗室,中國氣象局干旱氣候變化與減災重點實驗室,甘肅 蘭州 730020)
因水分供給不足而引發的干旱是限制全球作物產量增加的重要因素。然而,由于缺乏定量描述作物受旱階段性特征的指標,目前依然無法準確刻畫作物受旱進程[1],這導致我們無法精確模擬干旱脅迫下作物生長發育的動態過程[2],進而影響更加合理的灌溉制度的制定。盡管研究者給出了大量反映作物遭受干旱脅迫的指標(諸如土壤水分、土壤水勢、葉片相對含水量、葉水勢、冠層溫度以及作物水分脅迫指數等[3]),但是這些指標在確定作物受旱程度時,往往以最終產量為標準,即用受旱影響作物與供水充足作物最終產量之間的偏差作為作物受旱嚴重程度分類的依據[4]。這種劃分方法使我們無法有效地掌握干旱對作物生長發育影響的動態過程,阻礙了我們對農業干旱致害機理的認識[5]。因此,有必要在作物生長發育時期,對其受干旱脅迫影響的生理生態參數變化特征進行分析,以量化作物受旱程度。
作物本身具有較強的適應能力及應對干旱的調節能力,作物遭受輕度的干旱脅迫并不會對作物生長發育造成很大的影響[6]。在干旱脅迫發生初期,作物可通過產生大量脫落酸(有促進氣孔關閉的功能),降低細胞分裂素(有加劇作物水分虧缺的功能)的含量,調節作物體內細胞分裂素和脫落酸水平及其比例關系能夠在一定程度上維持植物體的水分平衡[7]。只有干旱脅迫超過一定的臨界值,作物才會遭受明顯的干旱影響,即作物生理參數對干旱脅迫存在閾值反應[8],而且不同生理參數的水分閾值不完全相同,這是由作物對干旱脅迫過程的適應和調劑機制不同而決定的[9]。以光合生理過程為例,在遭受干旱脅迫時,作物葉片凈光合速率(Pn)的下降并不是唯一一種調節機制在控制,而是既有氣孔因素又有非氣孔因素的作用[10]。在干旱脅迫相對較輕時,Pn受氣孔因素的影響呈緩慢的下降趨勢[10];當干旱脅迫繼續發展,作物葉片的光合能力受到損害,Pn將受到明顯抑制[11];而當嚴重的干旱脅迫造成葉綠體變形,葉綠體片層膜系統受損,使希爾反應減弱,光系統Ⅱ活力下降,電子傳遞和光合磷酸酸化受到抑制,則會造成Pn迅速下降[12],從而使作物干物質積累明顯減少,作物生長發育受到嚴重阻礙。
光合生理過程是作物產量形成的基礎[13],因而關注與光合生理過程相關的生理生態參數變化是研究作物受旱致害過程的關鍵。若以作物自適應特點和作物光合生理過程受不同程度干旱脅迫影響機制不同為依據,我們可假定,在作物遭受干旱脅迫至致害過程中光合生理參數應存在不同的變化特征,據此可用以確定作物受旱的不同階段。此外,一般認為氣孔導度(gs)和Pn呈線性關系[14],然而在干旱脅迫發展過程中,氣孔因素首先起作用,導致作物Pn下降,而隨著干旱脅迫程度加重,非氣孔導度因素的出現會導致Pn下降幅度增大,據此,我們推測非氣孔因素的疊加作用可能會導致gs與Pn的關系發生改變。
基于以上兩個研究假設,我們采用小區和桶栽干旱脅迫試驗,以小麥為供試作物,研究其受旱的階段性特征,從而提供一種可定量描述作物受旱過程的方法。
1 材料與方法
1.1 試驗觀測
試驗在中國氣象局蘭州干旱氣象研究所定西干旱氣象與生態環境試驗站內進行。該站位于甘肅省定西市安定區西川(104.37°E,35.35°N),海拔1 920 m,年平均氣溫6.3℃,年日照時數2 500 h。試驗供試作物為春小麥(定西新24號)。小區試驗于2017年進行,設置2個處理,分別為充足灌水和干旱脅迫處理,充足灌水處理整個生育期保證供水充足,干旱脅迫處理從小麥拔節期開始不灌水直至萎蔫。每個處理設4個重復,每個處理小區面積為2 m×1.5 m。桶栽試驗分別在2014、2015年以及2017年進行,桶栽試驗所用桶直徑29 cm,深度45 cm。供試土壤為黃綿土,從大田采集0~30 cm層次土壤,風干過篩,裝桶(每桶裝土14 kg)。桶裝土平均容重1.15 g·cm3,田間持水量26.8%,萎蔫系數5.5%,其中田間持水量和萎蔫系數均為重量含水量。試驗設兩個處理,各6組重復,共計12桶。春小麥于3月中旬播種,播種后至對比觀測開始前保持12個桶子處理方式相同,土壤水分供給一致。在春小麥長至拔節初期開始設置不同處理,其中一個為充足灌水處理,保證每日的土壤含水量在田間持水量的70%以上,另一個為干旱脅迫處理,在試驗開始后持續不灌水,直至小麥萎蔫。
試驗期間,主要觀測項目為:
(1)土壤含水量(本文均指土壤相對含水量)。每天稱重測量2個處理12個桶子質量(所用稱重天平最小感應為1g)。用所稱得的重量減去干土重量,再除以干土重量即為桶子中土壤重量含水量(忽略小麥本身生物量的變化)。
(2)光合生理過程。采用美國產Li-6400便攜式光合儀,在春小麥長至拔節期,不同試驗處理實施后,于每日上午9∶00—11∶30測定各處理春小麥最上部第一片完全展開葉Pn/Qp(光響應)和Pn/Ci(CO2響應)曲線(2014年和2015年試驗主要測定Pn/Qp曲線,2017年試驗主要測定Pn/Ci曲線)。Pn/Qp曲線的測量:控制CO2濃度在380 μmol·mol-1,葉室溫度控制在25℃,空氣飽和差控制在1.5~2.5 kPa。測量葉室采用紅藍光源,光合有效輻射(PAR)設置不同的梯度自動測量,梯度水平分別為0、15、30、60、120、200、300、600、900、1 200、1 500、1 800 μmol·m-2·s-1以及2 100 μmol·m-2·s-1。每次測量用1 500 μmol·m-2·s-1的PAR適應30~40 min,待儀器讀數穩定后,進入自動測量程序。Pn/Ci曲線測量時,初始CO2濃度控制在380 μmol·mol-1,葉室溫度控制在25℃,空氣飽和差控制在1.5~2.5 kPa之間,光強控制在1 500 μmol·m-2·s-1,開始測量之前先在初始條件下適應30~40 min,設置CO2梯度為400、200、100、50、400、600、800、1 000 μmol·mol-1和1 200 μmol·mol-1,依次自動測量。在觀測葉片氣體交換的同時,Li-6400還可記錄觀測時的葉室空氣溫度、相對濕度、葉氣溫差、葉氣飽和氣壓差以及空氣CO2濃度等氣象要素指標。
1.2 光合參數計算
Farquhar等[15]在1980提出了一個計算Pn的生化模型,其以胞間CO2濃度(Ci)分別計算在Rubisco(1, 5-二磷酸核酮糖羧化酶/加氧酶)活性和RuP2(核酮糖-1, 5-二磷酸)再生限制條件下的Pn:
Pn=Vc-0.5V0-Rd=min{Pc,Pj}-Rd
(1)
(2)
(3)
式中,Vc為羧化速率,V0為氧化速率,Rd為暗呼吸速率,Pc和Pj分別為受Rubiso活性和RuP2再生限制的光合速率,Vcmax是最大羧化速率,Г*是CO2補償點,Kc和Ko是CO2和O2的Michaelis-Menten動力參數,O是葉片胞間氧氣分壓,J是量子傳遞效率。(3)式中,J的計算依賴于光強和最大量子傳遞效率,依據下式計算獲得:
θJ2-(I2+Jmax)J+I2Jmax=0
(4)
式中,I2為光強,Jmax為最大量子傳遞效率,θ為曲度。
Vcmax和Jmax反映了作物葉片的光合能力,其值由Pn/Ci曲線觀測數據估算。當Ci值小于150 μmol·mol-1時,以(2)式估算Vcmax,當Ci大于150 μmol·mol-1時,用(3)和(4)式估算Jmax,具體計算過程可參照Xu等[16]的研究工作。
1.3 氣孔導度模型
Pn與gs是描述植物葉片氣體交換過程非常重要的兩個指標,而它們之間的關系又可由內稟水分利用效率(IWUE, 即Pn/gs)或半經驗氣孔導度模型反映[17]。當前有很多半經驗氣孔導度模型,而其中尤以Ball-Berry[18]模型的使用最普遍。Ball等[18]根據前人的研究結果,發現gs的變化與Pn、葉片表面相對濕度(RHs)以及葉片表面CO2濃度(Cs)有關:
gs=a1AnRHs/Cs+b1
(5)
式中,a1為氣孔導度模型斜率,由gs與PnRHs/Cs(BB參數)關系擬合獲得。b1為凈光合速率為零或小于零時的gs,部分文獻認為其為無光照時的gs[17],可由gs與PnRHs/Cs關系擬合截距獲得,或在黑暗條件下由光合儀測量獲得。Leuning[19]與Medlyn等[20]此后陸續分別提出了與Ball-Berry類似的2個模型,即Leuning和Medlyn模型,它們對gs模擬精度的改進有一定的幫助。其中Leuning(1995)模型的形式為:
gs=a2An/(Cs-Γ)(1+VPDa/b2)+c2
(6)
式中,a2為氣孔導度模型斜率,Г是CO2補償點,VPDa是空氣飽和氣壓差,b2和c2都是參數。Medlyn模型的形式為:
(7)
式中,a3為氣孔導度模型斜率,VPDL是葉片與空氣之間的飽和氣壓差,b3為參數。
1.4 反映干旱脅迫過程的指標
本研究采用土壤有效含水量(ASWC)和飽和光強下的氣孔導度(gssat)來反映小麥遭受干旱脅迫的狀況。其中ASWC計算如下:
(8)
式中,θa、θw以及θf分別表示實際土壤含水量、萎蔫含水量以及田間持水量。研究者發現gssat較其他水分指標更能有效地反映作物遭受干旱脅迫的狀況[21],由于本研究小區試驗小麥最大根系深度未知,因此以gssat作為另一個反映小麥水分供給狀況的指標。已有研究表明小麥光飽和點處于1 200~1 700 μmol·m-2·s-1之間[22],本文選擇光合有效輻射在1 500 μmol·m-2·s-1時的gs作為小麥的gssat。
1.5 數據分析
Pn/Qp曲線數據主要用以擬合求取3個不同氣孔導度模型的參數,在Origin 9.0軟件中使用非線性曲線擬合模塊(Nonlinear curve fit),自定義編制與(5)、(6)以及(7)式相同的公式擬合計算獲得所需參數。Pn/Ci曲線觀測數據主要用以估算Vcmax和Jmax,采用R語言以Duursma等[23]給出的“plantecophys”包計算獲得。同時,為證明干旱脅迫階段對氣孔導度模型參數的影響,本研究選擇不同年份(2014、2017年)和不同觀測來源(Pn/Qp和Pn/Ci曲線)數據,先以2014年Pn/Qp曲線數據擬合計算了不同干旱脅迫條件下不同光強的gs,并與實測值進行了對比分析;之后又將小區和桶栽2017年的數據整合,提取了飽和光強下的光合生理參數數據,分析比較了不同干旱脅迫階段氣孔導度模型參數的差異。
2 結果與分析
2.1 干旱脅迫下小麥葉片光合生理參數的變化
2.1.1 光合生理參數對干旱脅迫的響應 分析gssat與ASWC的關系(圖1a),發現gssat隨ASWC的變化存在3個階段,在第1個階段gssat變動不明顯,呈輕微的下降趨勢;而當ASWC減少至0.5左右時,gssat呈減小趨勢,但統計檢驗不顯著(P>0.05);而在第3階段,當ASWC小于0.3左右時,gssat快速線性下降,且統計檢驗極顯著(P<0.01)。同時,比較這3個階段的gssat,發現差異極顯著(圖1b)。
與gssat類似,Ci/Ca值隨著ASWC的變化也存在3個明顯的階段(圖1c),只是變化趨勢不完全相同。第1階段,Ci/Ca相對穩定,之后快速減小,在ASWC降至0.06開始,Ci/Ca呈增大的趨勢,最小值出現在ASWC為0.1時,此時Ci/Ca為0.32。同時,比較3個階段的Ci/Ca(圖1d),第2階段與其他2個階段差異極顯著,而第1和第3階段無顯著差異。
Pn隨ASWC的變化存在2個明顯的階段(圖1e),在第1階段,Pn基本保持穩定,而在ASWC降至0.35左右時,Pn隨ASWC的減少呈線性減小趨勢,且統計檢驗極顯著。而且第1階段與第2階段Pn值的差異也極顯著(圖1f)。
2.1.2 光合生理參數間的關系Ci/Ca和gs是決定作物葉片Pn的重要指標,觀察gssat以及Ci/Ca與Pn間的關系(圖2),這2個參數與Pn并不呈簡單的線性關系,而是隨水分條件變化存在階段性關系特征。
當gssat大于0.15 mol·m-2·s-1時,Pn隨gssat減小緩慢下降(圖2a),速率為1.64 μmol·m-2·s-1Pn/0.1 mol·m-2·s-1gs(P<0.001),而當gssat小于0.15 mol·m-2·s-1時,Pn隨gssat的降低快速減小,速率增至11 μmol·m-2·s-1Pn/0.1 mol·m-2·s-1gs(P<0.001),說明在以gssat=0.15 mol·m-2·s-1為界的2個不同階段,Pn對gssat變化的響應不同,即控制Pn變化的因素存在差異。
Ci/Ca與Pn的關系也存在2個明顯的階段(圖2b),即Pn與Ci/Ca呈線性正相關關系和Pn隨Ci/Ca增大而減小的階段。在第1階段,Pn隨Ci/Ca的減小呈顯著的減小趨勢,但是仔細觀察該階段依然可以分為2個子階段,即在Ci/Ca從0.8降至0.55的階段(gssat>0.06 mol·m-2·s-1),Pn隨Ci/Ca的變化降幅較小,而后下降速率加快。而當Ci/Ca降至0.32后(gssat<0.06 mol·m-2·s-1),Ci/Ca開始增大,但Pn卻隨Ci/Ca的增大依然在降低。
2.2 小麥葉片光合能力參數對干旱脅迫的響應
觀察小麥葉片Vcmax與Jmax在不同水分條件下的變化(圖3a,c),無論是在小區還是桶栽環境,小麥葉片的這2項參數對水分變化均存在明顯的閾值反應。當gssat大于0.15 mol·m-2·s-1時,隨gssat的降低,Vcmax和Jmax均保持相對穩定。而當gssat小于0.15mol·m-2·s-1時,隨gssat的降低,Vcmax和Jmax均呈快速降低趨勢(P<0.01)。而若以gssat=0.15 mol·m-2·s-1為界,再以生長環境為類別(小區和桶中),可將Vcmax和Jmax分為4組,經統計分析發現在gssat小于0.15 mol·m-2·s-1時,2個生長環境下的小麥葉片Vcmax和Jmax均小于gssat大于0.15 mol·m-2·s-1時的值,且統計檢驗極顯著(圖3b,d)。而在相同的gssat范圍內,不同生長環境下除Vcmax大田和桶栽有極顯著區別外,其他統計檢驗均不顯著。
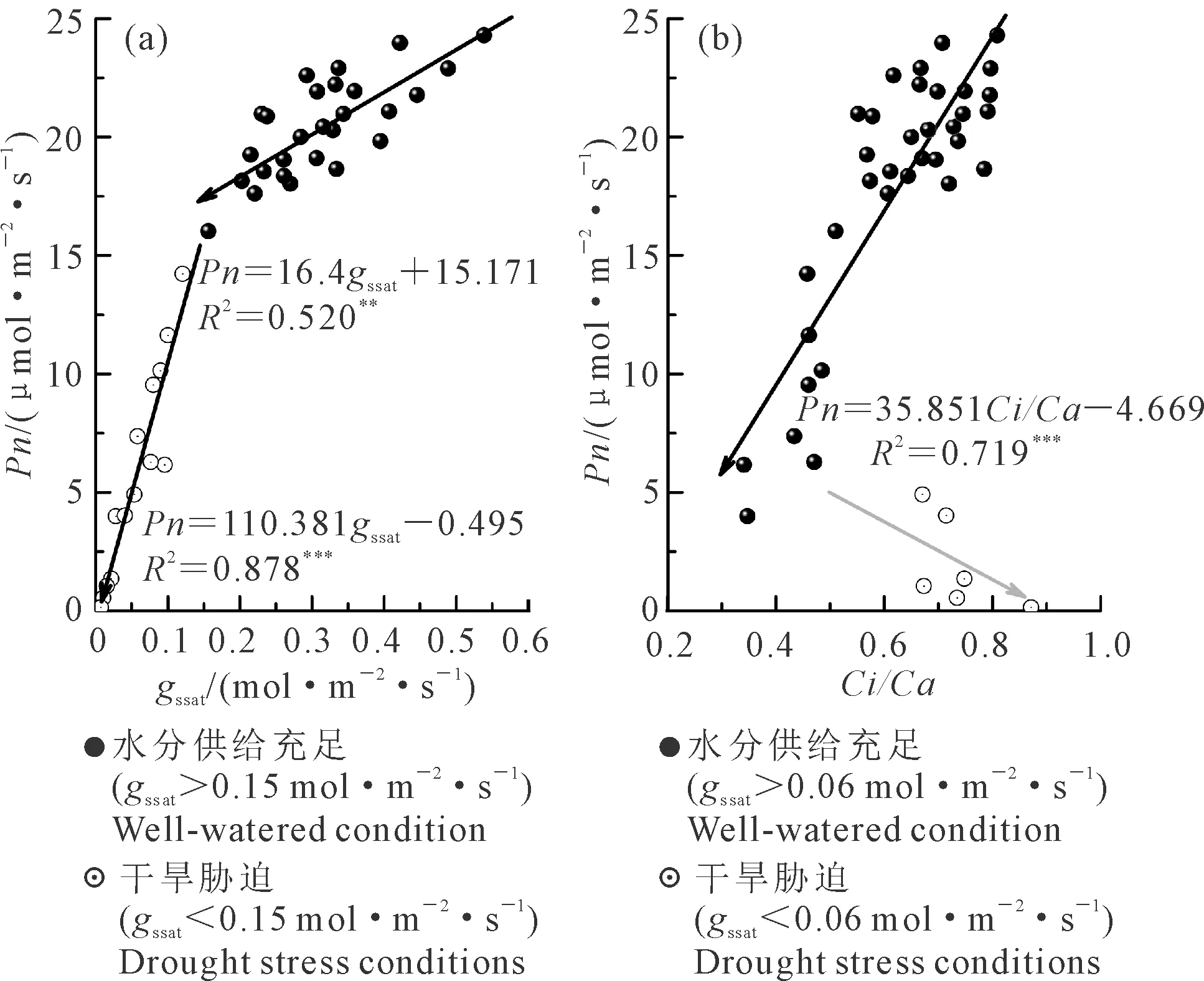
圖2 不同光合生理參數間的關系Fig.2 Relationship between different photosynthetic parameters
以gssat0.15 mol·m-2·s-1為界混合分析小區和桶栽小麥Vcmax和Jmax的關系(圖4a),發現當gssat大于0.15 mol·m-2·s-1時,Vcmax和Jmax關系并不明確,在相同的Vcmax值下,Jmax變幅較大。而當gssat小于0.15 mol·m-2·s-1時,Vcmax隨Jmax的減小呈極顯著的降低趨勢,同時比較以氣孔導度0.15 mol·m-2·s-1為界2個階段的Vcmax/Jmax(圖4b),發現統計檢驗極顯著。說明不同的水分條件影響Vcmax和Jmax的相互關系。
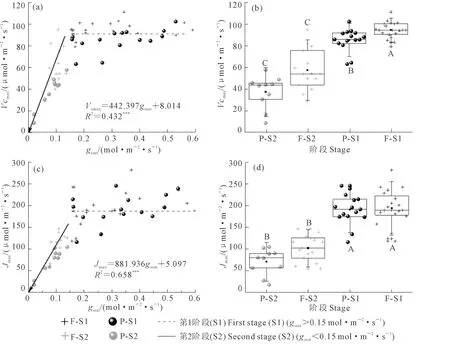
注:F-S1表示小區試驗光合生理參數變化的第1階段(gssat>0.15 mol·m-2·s-1),F-S2表示小區試驗光合生理參數變化的第2階段(gssat<0.15 mol·m-2·s-1);P-S1表示桶栽試驗光合生理參數變化的第1階段,P-S2表示桶栽試驗光合生理參數變化的第2階段。圖6相同。Note:F-S1, F-S2, P-S1, and P-S2 indicate the first (gssat> 0.15 mol·m-2·s-1) and second (gssat< 0.15 mol·m-2·s-1) stages of photosynthetic parameters variation for spring wheat growing in plots and pots, respectively. The same for Fig. 6.圖3 光合能力參數對干旱脅迫的響應Fig.3 Response of photosynthetic capacity to water stress
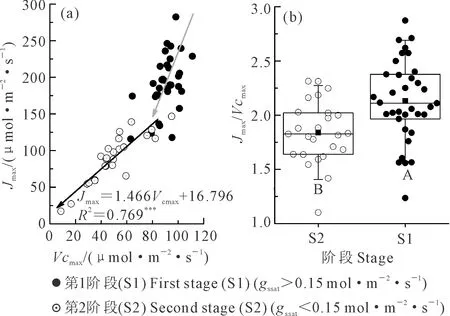
圖4 不同干旱脅迫階段Vcmax與Jmax的關系Fig.4 Relationship between Vcmax and Jmax under ifferent water stress stages
2.3 干旱脅迫下小麥葉片氣孔導度與凈光合速率之間的關系
若不區分干旱脅迫階段,使用擬合獲得的氣孔導度模型斜率計算2014年所觀測不同光強下的gs(圖5a、b、c),發現3個典型的氣孔導度模型實測值與模擬值均由1∶1線將數據明顯分為兩部分,其中一部分數據觀測值明顯偏大,而另一部分模擬值明顯偏大。對數據進行分類發現模擬值偏大的數據來源于gssat小于0.15 mol·m-2·s-1的部分,而實測值偏大的數據來自gssat大于0.15 mol·m-2·s-1的部分。而如果以gssat在0.15 mol·m-2·s-1處為界,將數據以2個不同的供水階段進行劃分,再次分析模擬值與觀測值的關系,發現模擬值與實測值的決定系數較不區分干旱脅迫階段時有較大的提高(圖5d、e、f),說明模型參數的選擇對氣孔導度模擬存在影響。
以Ball-Berry(1987)氣孔導度模型為例(其他2個模型結果類似,圖省略),整合大田和桶栽的數據,選擇飽和光強下的數據擬合氣孔導度模型,發現無論是生長在小區還是桶栽環境中的小麥,以gssat在0.15 mol·m-2·s-1為分界,不同水分供給階段gssat與BB指數的關系不完全一致(圖6)。在水分供給相對充足時(gssat大于0.15 mol·m-2·s-1),無論是小區還是桶栽,擬合所得的小麥氣孔導度模型斜率a1均較受水分脅迫程度嚴重時(gssat小于0.15 mol·m-2·s-1)的值大,其中在水分相對充足時小區和桶栽小麥a1分別為10.8和10.6,而水分相對不足時a1分別為7.5和6.9,水分供給條件不同,a1統計檢驗差異極顯著,而在相同水分條件下,不同生長環境對a1無顯著影響。
3 討 論
3.1 春小麥受旱過程的階段性劃分
gssat和Ci/Ca隨水分條件變化存在明顯的3個階段,而Pn的變化則存在2個階段,Pn與gssat以及Ci/Ca的關系也因不同水分條件存在2~3個階段。綜合來看,小麥葉片光合生理過程對干旱脅迫的響應存在4個明顯的階段,這種階段性變化是由于作物自身在隨干旱脅迫程度不斷加重時相應的調節機制不同決定的。
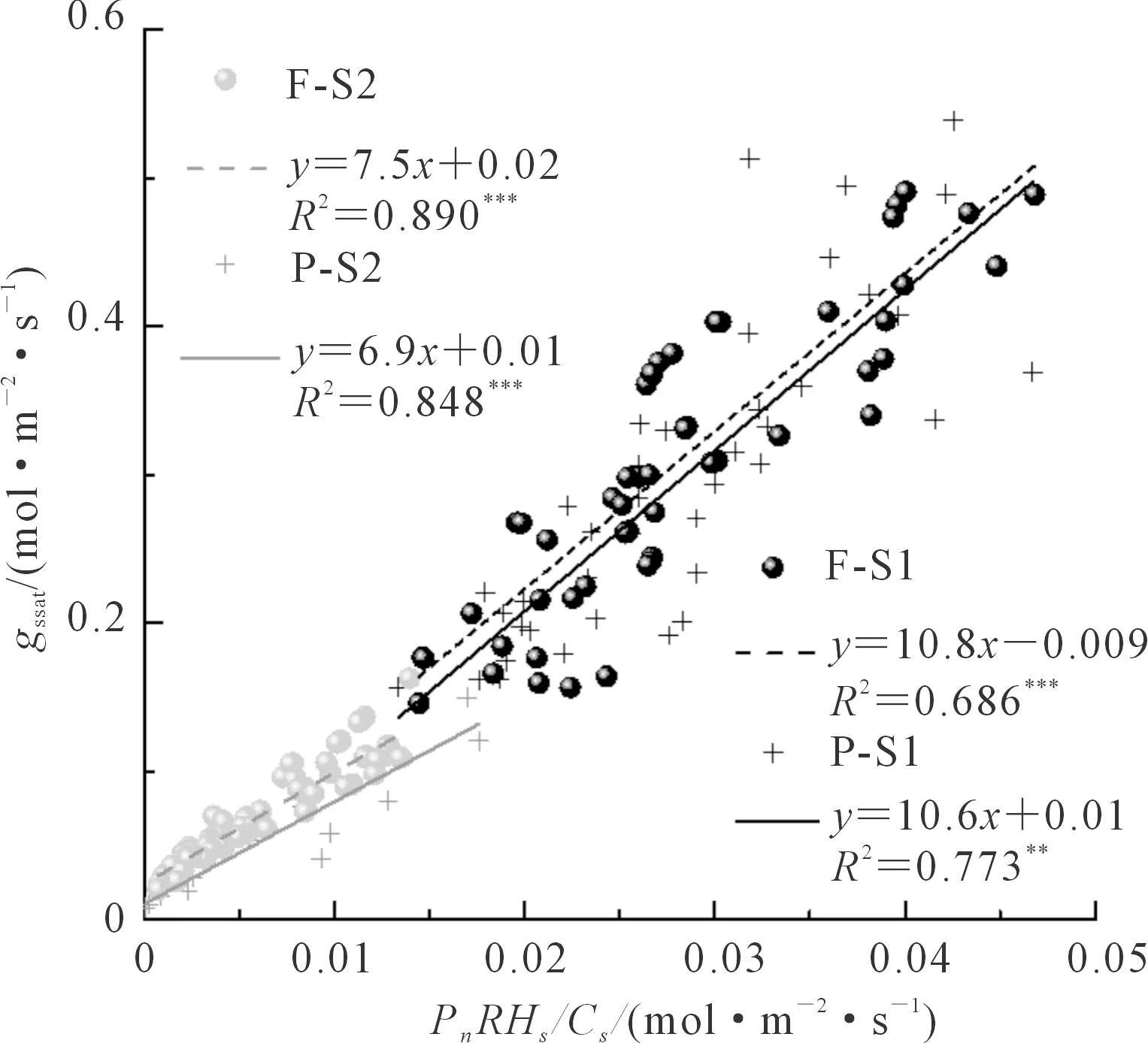
圖6 不同水分條件下的氣孔導度模型Fig.6 Stomatal conductance models under different water conditions
在干旱脅迫之初,隨著水分供給的減少,作物能夠通過自身的調節,適應水分的減少。當ASWC從1變化至0.5時(gssat>0.35 mol·m-2·s-1的階段,第1階段)(圖1a),gssat略有降低,而Ci/Ca值卻相對比較穩定,因此Pn無明顯的減少趨勢。而相較CO2,水分對gs的變化更敏感(CO2通過氣孔的阻力是水分的1.6倍),過大的蒸騰速率在該階段對作物的光合產出而言是無效的,該階段的光合生理參數變化特征間接說明了為什么有的研究發現輕到中度干旱脅迫下,植物葉片尺度水分利用效率(Pn與蒸騰速率比)或IWUE存在隨供水減少而增大的現象[10]。
當ASWC從0.5降至0.3時,gssat隨供水的減少呈波動減少趨勢(從0.35降至0.15 mol·m-2·s-1時,第2階段),但Ci/Ca在該階段呈極顯著的降低趨勢(直至ASWC降至0.1),而Pn也在該階段隨水分減少由穩定少動轉變為逐漸減小。同時觀察Vcmax和Jmax在該階段隨水分供給的變化保持相對穩定(圖3),Pn與gssat的關系以及Ci/Ca與Pn的關系也無明顯變化(圖2)。據此,我們推測,在該階段氣孔因素在起主導作用而影響小麥光合生理過程,gs的變化導致Ci/Ca值出現波動,從而使得小麥供給光合作用的場所,即葉綠體可用的CO2量減少。由于光合原材料不足,達到一定程度時,小麥葉片Pn出現降低的趨勢。
而在ASWC<0.3后,gssat進入一個新的變化階段(gssat<0.15 mol·m-2·s-1時,第3階段),即隨ASWC減小呈極顯著的降低趨勢。此外,從Vcmax和Jmax與gssat的關系來看(圖3),在該階段光合能力從之前的相對穩定到該階段開始隨干旱脅迫程度的增加而逐漸減小。而且Vcmax和Jmax之間的關系也以gssat在0.15 mol·m-2·s-1處為界有明顯的不同,gssat>0.15 mol·m-2·s-1時,Vcmax/Jmax值與許多作物水分供給充足條件下的研究結果一致[24],而當gssat<0.15 mol·m-2·s-1時,Vcmax/Jmax值明顯小于水分較為充足下的值,這與部分林木的觀測結果相近[25]。同時,gssat與Pn的關系也在氣孔導度<0.15 mol·m-2·s-1后發生變化,Pn隨gssat降低的幅度增加。這些現象充分說明,除氣孔因素外,與小麥光合生理過程相關的其他代謝活動(即非氣孔因素)由于干旱脅迫程度的增加開始限制Pn。
當ASWC降至0.1之后(氣孔導度<0.06 mol·m-2·s-1時,第4階段),隨作物供水的進一步減少,Ci/Ca呈增加趨勢,這是光合生理過程遭受干旱脅迫影響不同于上述3個階段的新時期。從Ci/Ca與Pn的關系(圖2b)也可以發現,該階段Ci/Ca值的變化已不能反映Pn的變化。然而,過去較早的一些研究結果往往以Ci/Ca隨干旱脅迫的加劇而出現增大的時期定義為作物光合生理過程遭受非氣孔因素影響開始的階段[26]。但本研究結果顯示在gssat小于0.15 mol·m-2·s-1時,影響作物生理過程的非氣孔因素已經出現,這與近年來的一些研究結果相近[27],即非氣孔因素可能在中等干旱條件下就已開始限制光合生理過程。本研究的第4階段,Ci/Ca的快速增大可能是由于嚴重的干旱脅迫導致小麥光合器官受損,使得光合生理過程出現了不可逆的破壞,這也是部分研究發現干旱脅迫到一定程度,即使恢復供水,而植物光合生理過程也無法恢復的原因[28]。
本研究對小麥受旱的階段性劃分與國外一些研究用gssat界定C3植物葡萄的受旱程度結果類似,即在葡萄生長遭受輕旱時,氣孔導度介于0.5或0.7 mol·m-2·s-1到0.15 mol·m-2·s-1,中旱0.05~0.15 mol·m-2·s-1,重旱<0.05 mol·m-2·s-1[29]。然而考慮到不同試驗條件,不同作物類型、土壤質地以及施肥條件等因素的影響,這種結論的相似性是偶然還是必然,還需進一步通過大量的試驗結果整合分析后確定。
3.2 不同干旱脅迫程度下氣孔導度模型的差異及其意義
氣孔導度模型斜率在干旱脅迫條件下是否會發生變化依然存在許多爭議,很多試驗研究結果相互矛盾[30-31]。本研究發現以gssat=0.15 mol·m-2·s-1為界,gs與Pn的關系有明顯的區別,而若不區分不同的干旱脅迫程度對氣孔導度模型參數的影響,3個典型的氣孔導度模型均會錯估gs。出現這種現象的原因是以gssat=0.15 mol·m-2·s-1為界,當gssat大于0.15 mol·m-2·s-1時,只有氣孔因素在限制光合生理過程,氣孔的調節作用導致CO2進入光合場所葉綠體的數量出現波動,引起Pn變動;而在gssat小于0.15 mol·m-2·s-1時,干旱脅迫加劇,非氣孔因素開始影響光合能力,而此時氣孔因素并非引起Pn變動的唯一因素,因此Pn與gs的關系必然發生變化,即在相同的gs變幅下,Pn變動更劇烈,這在圖3agssat與Pn的相互關系中也有反映,即隨干旱脅迫的加重,小麥IWUE在增大。因此,我們在使用氣孔導度模型時,需要根據干旱脅迫程度,選擇合適的模型參數。否則可能出現在水分供給充足時低估gs,干旱脅迫發生時高估gs的情形(圖5)。而其他一些研究[32]也表明,不準確的氣孔導度模型斜率不僅影響gs的準確計算,還導致對Pn、總初級生產力、蒸騰速率以及水分利用效率的錯誤估計。
氣孔導度模型在不同干旱脅迫階段的斜率差異,一方面表明氣孔導度模型需要根據作物所受脅迫程度調整參數從而以模擬獲得準確的gs,另一方面也再次間接證明了干旱脅迫的階段性特征,即在其他環境要素一定時,干旱脅迫條件下,以gssat=0.15 mol·m-2·s-1為界,大于該值,光合生理過程處于氣孔為主要限制因子影響Pn變化的階段,而小于該值后,轉變為非氣孔因素開始限制作物葉片光合生理過程的階段。
3.3 本研究存在的不足與展望
近年來學者認為干旱脅迫條件下葉肉導度的下降是導致光合生理過程下調的重要因子[33],然而在本研究中,并沒有單獨考慮葉肉導度在不同干旱脅迫階段對光合生理過程的影響,而是將其歸為非氣孔因素。在干旱脅迫條件下,葉肉導度的快速降低可能會影響Ci的準確測定,從而影響對Vcmax和Jmax的計算。但是由于葉肉導度的定量估計依然存在許多不確定性,因此分析葉肉導度在非氣孔因素中所占的比例是干旱脅迫對光合生理過程影響研究的重要方面,同時這也是農業干旱致害過程試驗研究與數值模擬亟需解決的問題。
過去的研究在確定作物受旱等級時,往往以產量作為劃分標準。而本研究所涉及的光合生理過程只是決定作物產量高低的眾多因素之一。而且作物不同生育期遭受干旱脅迫,對產量的影響不完全相同。例如,許多研究得出與光合生理過程相比,葉片的生長對干旱脅迫更敏感[34],在生育期前期,作物葉面積受旱而停止生長可能對產量影響更大。在自然條件下,一次降水或恢復供水后,光合等生理參數可能會迅速恢復至正常水平,但是葉面積的生長卻因水分虧缺而停滯,即使恢復供水也無法達到水分供給充足條件下的水平。作物光照截獲面積的減少,必然導致作物生物量積累減少。因此在研究農業干旱時,特別需要關注干旱脅迫對一年生作物生長盛期葉片生長的影響。本文只針對選定的生育期進行研究,沒有觀測葉面積變化對干旱脅迫的響應,在實際干旱監測中可能依然無法定量估計作物不同生育期所遭受干旱對其最終產量的影響。干旱脅迫對葉面積和光合生理過程的影響及二者對作物最終產量減損的作用還需進行更深入地分析和探討。
4 結 論
本研究依據試驗分析探討了光合生理過程對干旱脅迫的階段性響應特征,從而定量確定了干旱脅迫發展的不同階段,并討論了不同干旱脅迫階段Pn與gs的關系及其對氣孔導度模型使用的影響。小麥光合過程受干旱脅迫的影響,根據脅迫影響程度可劃分為4個階段,其臨界值以ASWC表示分別為0.48、0.3和0.1,而相對應的gssat為0.35、0.15以及0.06 mol·m-2·s-1。非氣孔因素開始影響光合生理過程的臨界點在ASWC為0.3時,非氣孔因素的出現導致Pn與gs的關系發生變化,從而在使用氣孔導度模型時,需要考慮不同的干旱脅迫程度對氣孔導度模型參數的影響,否則會出現水分供給充足時低估gs,干旱脅迫時高估gs的情形。本研究結果可為農業干旱等級定量劃分、作物模型改進以及農田灌溉制度的確定提供幫助。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
