MAPK信號通路基因在雷州山羊不同生長發育階段表達規律研究
2020-04-28 10:58:58陳智成練志全蔡丹鳳侯云飛楊健華李耀坤
甘肅畜牧獸醫 2020年2期
關鍵詞:信號
黃 強,陳智成,練志全,鄧 銘,蔡丹鳳,侯云飛,楊健華,李耀坤
(華南農業大學 動物科學學院,廣東 廣州 510642)
MAPK信號通路中,Ras/Raf/Mek(絲裂原活化蛋白激酶/Erk激酶)/Erk(細胞外信號調節激酶)途徑是控制細胞增殖、分化和存活的信號傳導網絡的核心[1-3],主要起正調控作用;Erk的激活對細胞增殖至關重要[4];Ras蛋白是Mek/Erk通路的激活因子,不僅能促進脂肪細胞的分化[5],也能促進成骨基因表達和成骨細胞增殖[6]。另有研究發現,山羊背最長肌組織的mRNA組學特性,在不同發育階段,呈現明顯的時序表達特征[7]。山羊至少有三個肌發生波,在胎兒發育的中后期(85 d左右)肌細胞明顯增加[8]。動物骨骼肌形成和發育機制一直是研究的熱點,關于動物肌肉生長和發育的基因調控已有很多報道,但大部分是檢測單個基因的在動物組織的表達情況[9],關于調控山羊肌肉生長發育的通路基因報道較少。
本次研究通過分析MAPK信號通路中Ras/Raf/Mek通路基因在雷州山羊不同生長發育時期背最長肌的表達情況,探究影響雷州山羊背最長肌生長發育的關鍵基因。實驗檢測了雷州山羊胎兒期90 d、100 d、120 d及出生后3月齡、6月齡的雷州山羊背最長肌的Ras/Raf/Mek通路基因表達水平,為進一步研究雷州山羊肌肉生長發育機制提供理論參考依據。
1 材料
1.1 實驗動物
雷州山羊妊娠第90 d、100 d、120 d胎兒各3只,及出生后3月齡、6月齡雷州山羊各2只,由溫氏羊業有限公司提供。
1.2 主要試劑
RNA提取試劑盒(型號為Total RNA Kit II R6934),購自OMEGA公司。
1.3 主要儀器
超凈工作臺(型號為SW-CJ-1B),購自中國蘇凈集團;高速冷凍離心機(型號為MicroCL 17R),微量核酸蛋白質測定儀(型號為Nano-200),實時熒光定量PCR儀(型號為QuantStudio 7 Flex System),均購自賽默飛世爾科技公司。
2 方法
2.1 總RNA提取和反轉錄
采用OMEGA Total RNA Kit II R6934試劑盒對動物組織總RNA進行抽提,RNA用ND2000超微量核酸蛋白質測定儀檢測抽提得到的RNA的OD值和濃度,用1.5%的瓊脂糖凝膠電泳檢測RNA的完整性及是否存在DNA和蛋白質的污染。
去DNA反應體系:5×DNA Eraser Buffer 2μl,gDNA 1 μl,Total RNA 7 μl。加完后混勻、離心、PCR儀按設置好的程序運行。
反轉錄體系為:去DNA反應液 10 μl,Prime ScriPt RT Enzyme MixⅠμl,RTPrimer mix 1 μl,5×Primer ScriPt Buffer 4 μl,RNA-free H2O 4 μl。加完后混勻離心,PCR儀按設置好的程序運行。反轉錄后的cDNA放置-20 ℃冰箱保存。
2.2 引物設計
根據GenBank (httP://www.ncbi.nlm.nih.gov/)上公布的候選基因序列,以及通過查詢資料和引用其他文獻資料,采用Primer Premier 5.0軟件設計獲得下列引物,引物由北京六合華大基因科技有限公司合成,引物信息見表1。
2.3 實時熒光定量PCR
本實驗采用SYBRGreen染料法進行熒光定量PCR反應。熒光定量PCR的反應體系為:上、下游引物各0.3μl,2×SYBRGreen熒光染料5μl,cDNA模板1 μl,加ddH2O補至總體積10 μl;熒光定量PCR的擴增程序為:45個循環(95℃預變性10 min;95℃變性15 s,60℃退火10 s,72℃延伸15 s);58℃檢測熒光值;15℃保存。

表1 熒光定量Q-PCR所用引物的相關信息
2.4 統計分析
本實驗采用2-ΔΔCt法計算各個基因的mRNA相對表達量,以β-Actin為內參基因進行標準化校準。所得數據采用SPSS 20.0統計軟件One-way ANOVA進行分析。
3 結果與分析
3.1 Ras、Raf、RafA、Mek1和Mek2基因的組織表達譜
采用qRT-PCR技術,以β-actin作為內參,以腎組織為參照,結果表明(圖1)Ras、Raf、RafA、Mek1和Mek2基因在8個組織中均有表達。Ras、Raf、RafA、Mek1和Mek2基因在背最長肌和心臟中的表達量最高,在下丘腦、肝、脾、肺和垂體中的表達量較低。
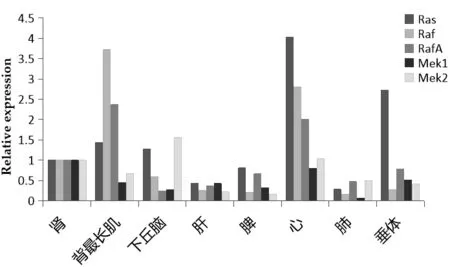
圖1 Ras、Raf、RafA、Mek1和Mek2基因的組織表達譜分析
3.2 不同生長發育時期Ras、Raf、RafA、Mek1和Mek2基因在背最長肌的表達情況
Ras基因在胎兒發育120 d齡時的表達量最高,表達模式為:120 d>100 d>90 d>6月齡>3月齡,120 d齡胎兒背最長肌中的基因表達量顯著高于6月齡羔羊。Raf基因在羔羊3月齡的表達量最高,3月齡羔羊的表達量顯著高于90 d和100 d胎兒。RafA基因在胎兒發育120 d齡時的表達量達到最高,90 d齡的表達量最低,在出生后6月齡的表達量高于3月齡,但各組間基因的表達量差異不顯著。Mek1基因在120 d的表達量顯著高于其它組,其余組間基因的表達量差異不顯著。Mek2基因在胎兒期120 d的時候基因的表達量最高,120 d的表達量顯著高于100 d;90 d、100 d、3月齡和6月齡的基因表達量沒有顯著差異(圖2)。
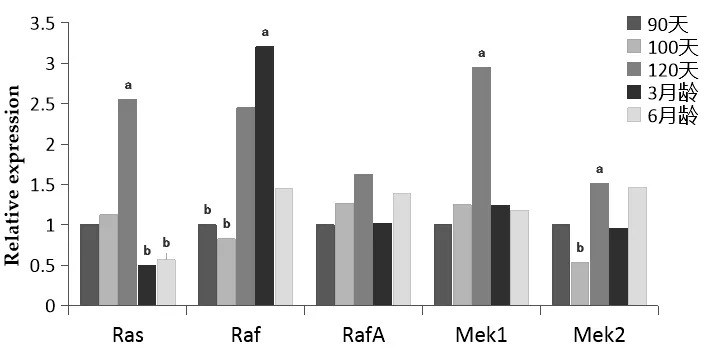
圖2 Ras、Raf、RafA、Mek1和Mek2基因在背最長肌的表達情況
3.3 不同生長發育時期Ras、Raf、RafA、Mek1和Mek2基因在心臟的表達變化規律
Ras基因在羔羊3月齡時的表達量最高,在胎兒期100 d的表達量最低,表達模式為:3月齡>90 d>6月齡>90 d>100 d,3月齡羔羊心臟中的基因表達量顯著高于其它組。Raf基因在羔羊3月齡的表達量最高,100 d齡的表達量最低,3月齡羔羊的表達量顯著高于其它組。表達模式為:3月齡>6月齡>120 d>90 d>100 d。RafA基因在羔羊3月齡時表達量最高,3月齡基因的表達量顯著高于其它組。Mek1基因在胎兒期120 d的表達量顯著高于其它組,3月齡和6月齡羔羊表達量顯著高于胎兒期90 d和100 d。Mek2基因在羔羊3月齡時基因的表達量最高,3月齡的基因表達量顯著高于其它組,90 d、100 d、120 d和6月齡的基因表達量沒有顯著差異(圖3)。
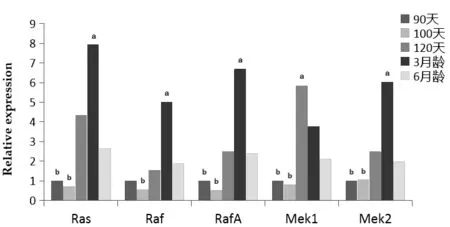
圖3 Ras、Raf、RafA、Mek1和Mek2基因在心臟的表達情況
4 討論
雷州山羊是我國著名的地方優良品種,也是廣東省唯一的肉用羊地方良種。雷州山羊具有瘦肉多、脂肪少、膻味輕、營養豐富、味美多汁、易消化吸收等特點,深受消費者青睞[10],其市場需求量巨大,市場前景十分廣闊,具有較大開發潛力。但近年來,雷州山羊品種退化十分嚴重,產肉性能在逐年下降[11],針對優質山羊品種存在的生長不穩定、生長速度慢等問題,本研究通過分析MAPK信號通路中相關基因在雷州山羊各個生長發育時期的表達規律,為雷州山羊背最長肌的生長發育的分子機理的闡明提供證據,同時也為肉羊生長發育機制提供科學依據。MAPK信號通路中的Ras/Raf/Mek/Erk細胞信號傳遞通路是由一個GTP結合蛋白連接活化的受體酪氨酸激酶和胞漿蛋白激酶級聯反應介導,其活化的中心是使Ras進行鳥苷酸交換變成其活化形式Ras-GTP[12]。該通路可由活性氧、Ca2+、蛋白激酶C等激活,參與體內的多種生理生化功能,對細胞生長、分化、增殖、發育、凋亡均有影響[13]。Ras的表達對Raf有促進作用,其機理為Ras-GTP直接與Raf結合,形成一個臨時的膜錨定信號。而活化的Raf再通過磷酸化促進分裂原激活蛋白激酶的激酶(Mek)環上的絲氨酸殘基而將其激活[14]。Mek/Erk通路參與了胰島素調控骨骼肌成肌細胞增殖,具有促增殖或抗凋亡作用,是多途徑多通路的信號轉導過程[15],抑制此通路,可以抑制平滑肌的增殖[16],也可以抑制骨骼的細胞增殖、成熟和鈣沉積[17]。小鼠敲除Raf基因實驗表明,Raf基因在各組織生成中均具有一定功能[18],尤其對胎兒生長發育發揮關鍵作用。基于上述原因,如果能確定Ras/Raf/Mek信號通路基因在雷州山羊背最長肌表達量的差異和表達規律,就可采用基因敲除、轉基因、注射抗體、反義核酸等技術,控制其表達量,從而使其背最長肌充分發育,提高動物的生長性能。另一方面,通過有關基因表達量與生長發育規律相聯系,有助于掌握雷州山羊生長發育特點,以便在妊娠母羊胎兒生長發育關鍵時期和正常的生長發育時期合理調控外界環境和營養條件,充分挖掘其生長潛能。
雷州山羊的組織表達譜分析結果顯示,Ras、Raf、RafA、Mek1和Mek2基因均在雷州山羊的背最長肌有較高的表達量,說明其對調控肌肉生長發育有重要作用。在心臟組織中,3月齡羔羊的Ras、Raf、RafA和Mek2基因表達量顯著高于其他組,Mek1在胎兒期120 d表達量顯著高于其他組。小鼠心臟發育開始于胚胎發育期的第7.5天,在胚胎第11.5天之后心肌細胞大量增殖,促心生長因子有Hedgehog、骨形成蛋白(bone morphogenetic proteins, BMPSs)、成纖維生長因子(firbroblast groth factors, FGFs)和非經典Wnt/JNK通路[19],但關于山羊心臟發育過程中的分子調控研究報道較少,本文可為雷州山羊心臟生長發育機制提供一定的理論參考。在所研究的雷州山羊胎兒期的三個生長階段,Ras、Raf、RafA、Mek1和Mek2基因均在120 d表達量最高;表達模式為:120 d>100 d>90 d。120 d Mek1基因mRNA的表達量顯著高于100 d、90 d的基因表達量山羊妊娠周期為150 d,其胎兒生長發育后期(60 d以后)增長的體重占初生重90%,在妊娠后期胎兒的生長發育速度越來越快。本研究結果表明,Ras/Raf/Mek信號通路基因在雷州山羊的背最長肌發育中呈相同的表達模式,此通路對細胞的生長、增殖發揮著關鍵的作用[20]。根據實驗結果推測,雷州山羊在胎兒生長發育120 d的時候有一個肌肉發生波。綜合判斷,Ras/Raf/Mek信號通路在雷州山羊胎兒生長發育后期起關鍵作用,并且起正調控作用,但具體機制尚不明確。通過分析3月齡、6月齡相關基因表達情況,并未發現較明顯的基因表達模式,可能該通路基因在出生后的調控機制發生了變化,具體的調控機制仍有待深入分析。
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
