幾種不同菌種發酵豆粕發酵過程指標對比
2020-04-20 15:57:32江國托
飼料博覽 2020年1期
劉 星,趙 榮,李 波,江國托,劉 艷*
(1.江蘇三儀科研質量控制中心,江蘇 邳州 221300;2.盤錦市獸藥飼料監察所,遼寧 盤錦 124010;3.大連三儀生命科學研究院,遼寧 大連 116036)
豆粕是目前飼料工業中應用最廣泛的植物蛋白源。但由于其自身的理化特點如氨基酸不平衡,存在多種抗營養因子,質地結構致密,導致其適口性較差,不易被完全消化,限制了豆粕的應用價值。尤其是在國家倡導節能減排,降低飼料中蛋白含量,提高我國飼料蛋白自主性,減少對大豆進口的依賴性的社會環境下,如何更好的利用大豆蛋白成為迫切的需要[1]。
發酵豆粕的實質就是利用發酵技術對豆粕進行發酵預處理,其目的是提高豆粕的飼用品質。豆粕中的蛋白多為大分子蛋白,肽鏈較長,難以被動物直接消化利用,且消化率不高[2]。利用發酵技術,可以將大豆蛋白較長的肽鏈變成較短的肽鏈,且在發酵過程中產生多種風味物質,使豆粕由致密變得較蓬松,從而提高其適口性,促進動物采食[3-4]。同時,發酵過程中,有益菌大量繁殖。在用發酵豆粕飼喂動物的同時,實際上也飼喂了動物大量有益菌,這對維持動物腸道健康平衡,提高腸道吸收營養效率均有益處[5]。
雖然發酵豆粕的應用已經有較長的歷史,其顯著作用也被行業廣泛認可,但由于發酵豆粕工藝繁多,菌種和酶制劑多種多樣,使得不同工藝的發酵豆粕其指標差異較大,且不同發酵工藝的發酵豆粕指標平行比較的數據較少或者較不完整[6]。本試驗將幾種具有代表性的發酵工藝而得的發酵豆粕進行主要指標的平行對比,為發酵豆粕廠家及用戶選擇何種工藝的發酵豆粕提供一定的參考。
1 材料與方法
1.1 菌種及培養條件
植物乳酸桿菌SAM0038(實驗室保藏),MRS培養基,37℃,靜置培養[7];丁酸梭菌SAM0109(實驗室保藏),參考夏會麗等方法[8];釀酒酵母(實驗室保藏)SAM0252,PDA培養基,30℃,220 r·min-1[9];枯草芽孢桿菌(實驗室保藏)SAM0125,LB培養基,37 ℃,220 r·min-1[10]。
蛋白酶(酶活力10萬U·g-1,購自河南仰韶生化工程有限公司)[11]。
1.2 樣品處理方法
分別將各菌種發酵液以5%(w/w)的比例加入至發酵豆粕中,調整水分為48%,混勻。植物乳酸桿菌處理的豆粕直接裝入排氣袋,密封,置于37℃48 h;釀酒酵母處理的豆粕直接裝入排氣袋,敞口置于37℃48 h;丁酸梭菌處理的豆粕放入排氣袋中,抽真空后,密封,置于37℃48 h;枯草芽孢桿菌處理豆粕,鋪平放于不銹鋼托盤中,置于37℃48 h;蛋白酶按照0.1%(w/w)的比例添加入豆粕中,調整水分48%,混勻鋪平于不銹鋼托盤中,置于37℃48 h。所有實驗樣品60℃(試驗過程發現,烘干溫度>75℃,影響蛋白溶解度;<45℃較難烘干)烘干后,測量水分,并換算成與原料水分11.56%相同水分條件下進行其他指標平行對比。
1.3 指標測量方法
水分測定方法:GB/T 6435-2014;pH測定方法:DB15/T 1458-2018;酸溶蛋白測定方法:GB/T 22492-2008;KOH蛋白溶解度測定方法:GB/T 19541-2017附錄A;揮發性鹽基氮測定方法:GB/T 32141-2015;總酸測定方法(以乳酸計):GB/T 12456-2008;蛋白電泳測定方法(SDS-PAGE方法):稱取0.1 g樣品與20 mL樣品緩沖液混合均勻,并于沸水中煮沸5 min,冷卻至室溫。濃縮膠和分離膠濃度分別為12%和5%。上樣量為15 μL,電泳緩沖液:0.19 mol·L-1甘氨酸,0.1%十二烷基磺酸鈉和0.025 mol·L-1三羥甲基氨基甲烷。于單向垂直電泳儀中,25 V 30 min,50 V 90 min。染色液染色30 min,脫色液脫色4 h。各溶液配制方法參考詹玉春等方法[12]。
2 試驗結果
2.1 各工藝發酵豆粕粗蛋白質變化曲線
發酵豆粕粗蛋白質變化曲線見圖1。

圖1 發酵豆粕粗蛋白質變化曲線
由圖1可知,各發酵工藝發酵豆粕粗蛋白質含量均有所上升,其中枯草芽孢桿菌發酵對豆粕蛋白質的濃縮作用最明顯,發酵48 h后,粗蛋白含質量從45.28%提高至48.37%(換算成同一水分條件下,下同)。
2.2 各工藝發酵豆粕pH變化曲線
發酵豆粕pH變化曲線見圖2。
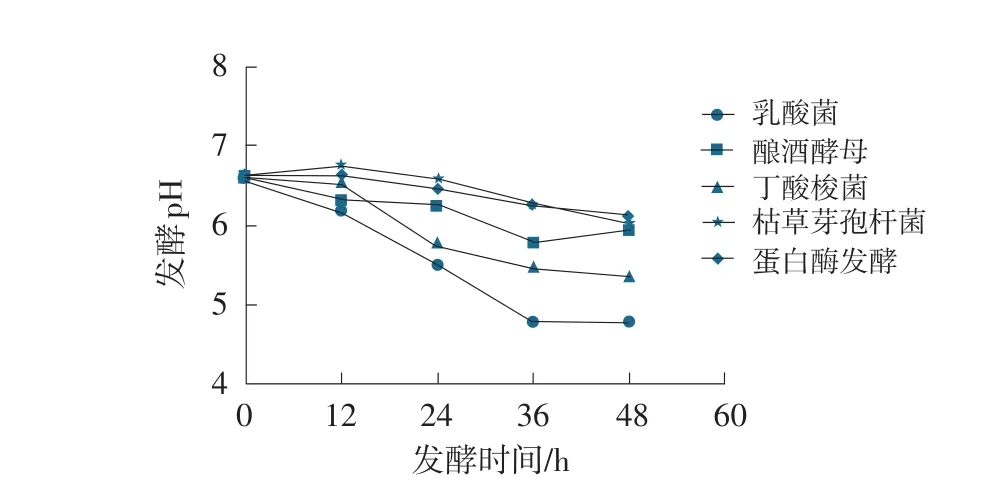
圖2 發酵豆粕pH變化曲線
豆粕經過幾種不同發酵工藝發酵后,pH均有所降低。其中,植物乳酸桿菌在排氣袋密閉條件下發酵豆粕pH降低最為明顯,發酵36 h后,pH可降至4.76。在48 h內,除釀酒酵母處理外,乳酸菌、丁酸梭菌、枯草芽孢桿菌,蛋白酶制劑發酵豆粕pH均呈逐漸降低變化趨勢,釀酒酵母發酵豆粕發酵pH在36 h之后,稍有上升。
2.3 各工藝發酵豆粕總酸變化曲線
發酵豆粕總酸變化曲線見圖3。
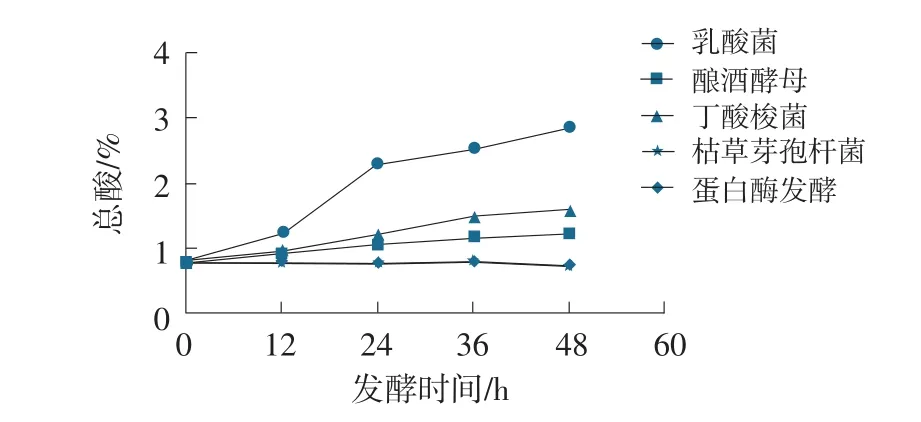
圖3 發酵豆粕總酸變化曲線
在發酵48 h周期內,乳酸菌發酵產酸量(以氫氧化鈉滴定總酸計)最為明顯,產酸量最高可達2.86%。釀酒酵母、丁酸梭菌產酸量較低,最高分別可達1.21%、1.62%。枯草芽孢桿菌與蛋白酶發酵豆粕含酸量基本持平,變化較小。
2.4 各工藝發酵豆粕酸溶蛋白變化曲線
發酵豆粕酸溶蛋白變化曲線見圖4。
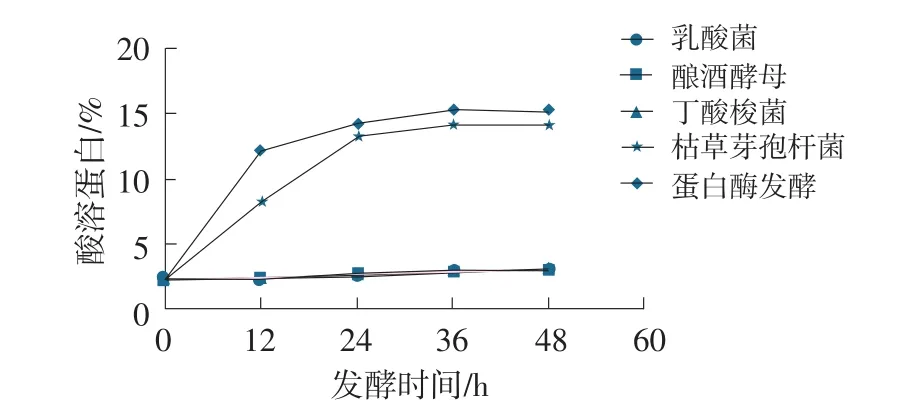
圖4 發酵豆粕酸溶蛋白變化曲線
在發酵周期48 h內,乳酸菌、釀酒酵母、丁酸梭菌發酵豆粕其酸溶蛋白含量較原料豆粕無明顯提升;枯草芽孢桿菌、蛋白酶制劑發酵豆粕酸溶蛋白含量提升明顯,最高分別可達14.27%、15.28%,說明枯草芽孢桿菌和蛋白酶制劑對豆粕中蛋白有顯著的降解作用;乳酸菌、釀酒酵母、丁酸梭菌對豆粕中蛋白降解作用不明顯。
2.5 各工藝發酵豆粕KOH蛋白溶解度變化曲線
發酵豆粕KOH蛋白溶解度變化曲線見圖5。
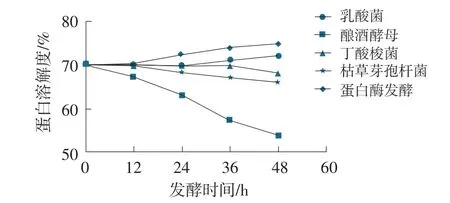
圖5 發酵豆粕KOH蛋白溶解度變化曲線
在發酵周期48 h內,乳酸菌、丁酸梭菌發酵豆粕KOH蛋白溶解度與原料豆粕比較接近,從初始值70.16%分別變為72.18%、68.25%;蛋白酶制劑發酵豆粕發酵48 h后,KOH蛋白溶解度有所提升,從初始值70.16%提升至75.14%;釀酒酵母、枯草芽孢桿菌發酵豆粕發酵48 h后,KOH蛋白溶解度有所降低,從初始值70.16%分別降低至54.12%、66.37%。
2.6 各工藝發酵豆粕揮發性鹽基氮變化曲線
發酵豆粕揮發性鹽基氮變化曲線見圖6。
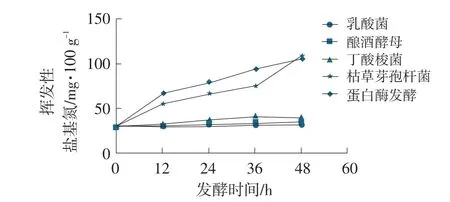
圖6 發酵豆粕揮發性鹽基氮變化曲線
在發酵周期48 h內,乳酸菌、釀酒酵母和丁酸梭菌發酵豆粕中,揮發性鹽基氮(VPN)與原料豆粕相比變化不明顯;但枯草芽孢桿菌與蛋白酶發酵豆粕中揮發性鹽基氮(VPN)與原料豆粕相比顯著增高,枯草芽孢桿菌發酵豆粕中揮發性鹽基氮含量最高可達109.27 mg·100 g-1,蛋白酶發酵豆粕中揮發性鹽基氮含量最高可達106.82mg·100 g-1。
2.7 各工藝發酵豆粕SDS-PAGE圖
發酵豆粕SDS-PAGE條帶見圖7。

圖7 發酵豆粕SDS-PAGE條帶
發酵48 h后,從蛋白電泳圖結果來看,乳酸菌發酵豆粕,釀酒酵母發酵豆粕,丁酸梭菌發酵豆粕中蛋白與原料中蛋白比較相似。枯草芽孢桿菌發酵豆粕和蛋白酶發酵豆粕中,大分子蛋白含量較原料豆粕明顯減少,相應的,中小分子蛋白含量明顯增多。說明,枯草芽孢桿菌和蛋白酶降解大豆蛋白的作用比較明顯,而乳酸菌、釀酒酵母和丁酸梭菌在降解豆粕中蛋白作用,效果不明顯。
3討論
發酵豆粕中,酸溶蛋白含量、揮發性鹽基氮和蛋白電泳條帶中蛋白分布可能有一定關系。酸溶蛋白含量高,說明大豆中蛋白的降解程度可能較高,大豆蛋白中有一部分大分子蛋白轉化為小分子蛋白,從蛋白電泳條帶上來看就呈現出上面代表大分子蛋白的條帶減少或變淺,下面代表小分子蛋白的條帶增多或顏色加深,與理論上的蛋白條帶變化相一致。豆粕中蛋白的進一步分解,可能會導致氨氣的產生,從而導致揮發性鹽基氮的升高。發酵飼料通常以酸溶蛋白含量高,蛋白溶解度高,揮發性鹽基氮含量低為質量較高。如果是幾種合適的菌種組合或菌酶協同發酵,可能會提高酸溶蛋白和蛋白溶解度的同時,不提高或降低揮發性鹽基氮的含量。
工藝不同可能會對發酵豆粕的KOH溶解度造成明顯的差異。乳酸菌、枯草芽孢桿菌、丁酸梭菌、蛋白酶制劑對豆粕的KOH溶解度影響不顯著,但釀酒酵母發酵豆粕KOH蛋白溶解度顯著降低,由70.16%降低至54.12%。
各發酵工藝存在一定缺陷,如排氣袋即使抽真空密封也無法保障嚴格厭氧環境,不利于丁酸梭菌生長發酵,都用37℃溫室發酵,不一定是其最適溫度。同理,48%的發酵水分也不一定是其最佳水分,其他因素如pH等因素也應考慮。
4結論
枯草芽孢桿菌提高粗蛋白質含量最優,將原料蛋白45.28%提高至48.37%(同一水分下)。乳酸菌密閉發酵豆粕產酸量最高,產酸量可達2.86%。枯草芽孢桿菌和蛋白酶制劑提高酸溶蛋白含量和揮發性鹽基氮作用明顯,其酸溶蛋白含量分別可達14.27%和15.28%;其揮發性鹽基氮含量分別可達109.27和106.82 mg·100 g-1。釀酒酵母發酵豆粕KOH溶解度降低明顯,從70.16%降低至54.12%,枯草芽孢桿菌和蛋白酶制劑發酵豆粕揮發性鹽基氮含量升高明顯。
