圓紅冬孢酵母脂肪酸合酶的生物信息學分析
2020-04-17 13:47:02劉玉青胡煥煥豐慧根
中州大學學報 2020年1期
康 靜,劉玉青,胡煥煥,豐慧根
(新鄉醫學院 生命科學技術學院,河南 新鄉453003)
基于生物質轉化生產的生物柴油是當今最為理想的可再生燃料之一[1]。圓紅冬孢酵母(Rhodosporidiumtoruloides)是目前研究較多的高產油脂菌種,作為生物柴油的原料,與藻類或植物生物柴油相比,它可在胞內積累油脂、類胡蘿卜素等[2],它不僅是清潔能源,更具有可持續生產微生物類脂和工業化廣泛應用的潛力。該菌株經酯交換產生生物柴油[3],但反應速率慢,反應體系中過多的甲醇和副產物甘油還會影響脂肪酶的活性及穩定性[4]。對圓紅冬孢酵母油脂代謝中的關鍵酶脂肪酸合酶(Fatty acid synthase,FAS)的結構特征及理化性質進行預測,可以為下一步構建產油工程菌奠定基礎,也為生物柴油及其他相關行業的原料油脂資源和廢棄物的再利用提供新的思路[5]。
1 材料與方法
1.1 FAS基因序列的Genbank檢索
在NCBI的Genbank數據庫中檢索圓紅冬孢酵母脂肪酸合酶,其基因序列編碼為XM_016415817.1,氨基酸序列編碼為XP_016272387.1。
1.2 FAS的生物信息學分析及預測
1.2.1 構建系統進化樹
在NCBI數據庫中檢索20種生物FAS氨基酸序列,利用MEGA7.0軟件對20種生物相應的氨基酸序列進行遺傳距離的分析,構建出FAS的系統進化樹。
1.2.2 使用在線生物信息學預測工具
ProtParam:對FAS進行理化性質的預測,如分子量、等電點、原子構成、氨基酸組成、摩爾消光系數、半衰期、脂溶指數等;ProtScale:對FAS進行親水參數的預測;TMpred:對 FAS 進行跨膜區域的預測;SigalP:對FAS進行信號肽的預測;PredictNLS:對FAS進行核輸出信號的預測;SOPMA和PredictProtein:對FAS進行蛋白質二級結構折疊類型的預測分析;SMART:對FAS進行蛋白質結構域的預測與分析。
2 結果
2.1 FAS進化樹的構建
使用鄰位相連法(Neighbor-joining,NJ 法)構建FAS的系統進化樹,如圖1所示。將20種FAS的氨基酸序列對齊后,使用軟件MEGA 7.0構建系統進化樹。由圖1可知,該FAS與紅酵母屬的菌種都具有較高同源性,且進化位置上也比其他菌種要高。
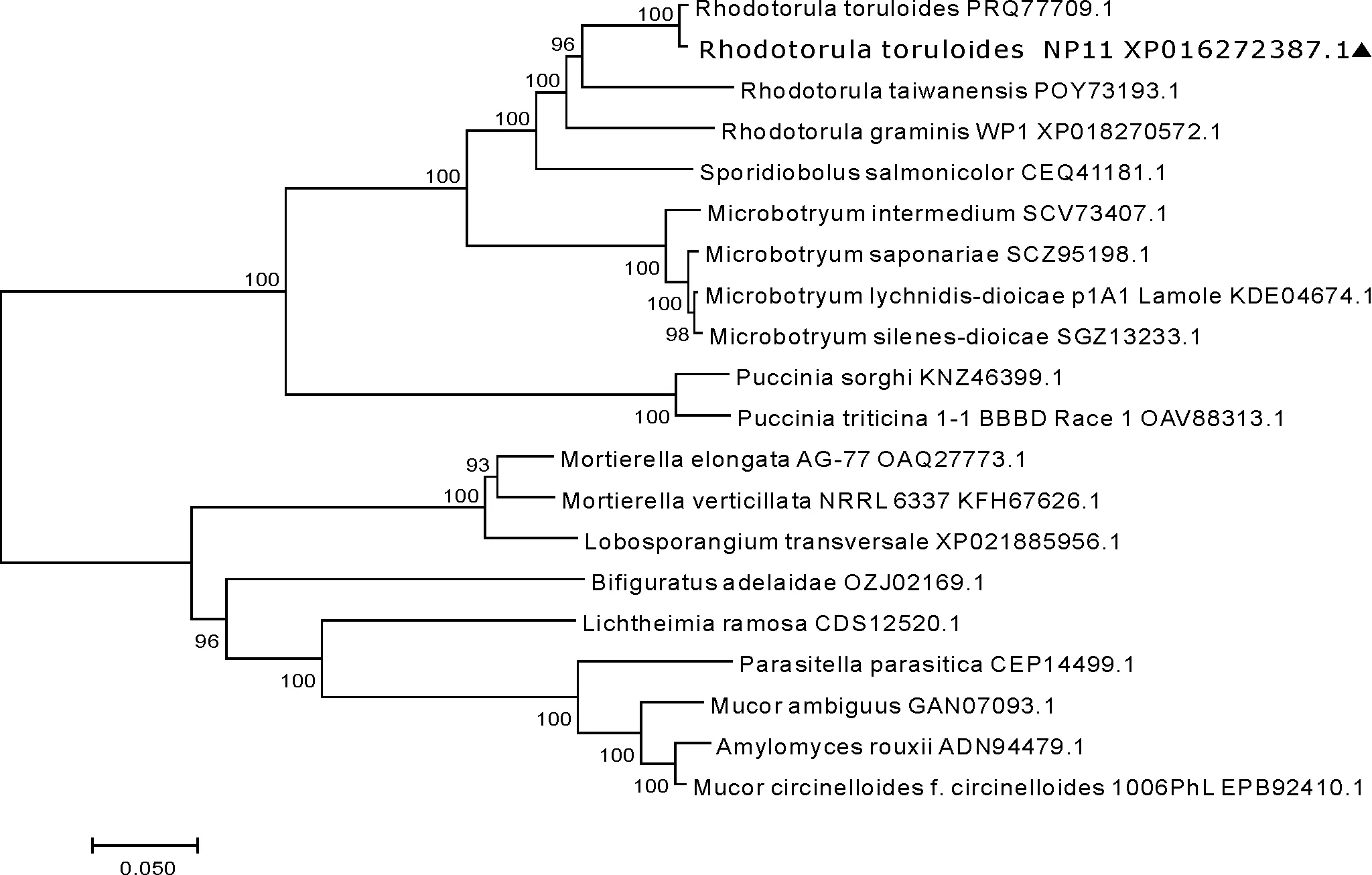
圖1 圓紅冬孢酵母FAS系統進化樹
2.2 氨基酸的組成
利用在線軟件Protparam對該FAS編碼的氨基酸組成進行初步預測分析。如圖2所示,預測結果:該FAS編碼區的氨基酸數目為2928個,其中含量較多的氨基酸為Ala(311個,10.6%),Leu(260個,8.9%)和Gly(250個,8.5%)。該FAS氨基酸中,帶正電的總數殘基/酸性氨基酸(Arg+Lys=320)少于帶負電的總數殘基/堿性氨基酸(Asp+Glu=338)[6]。它還含有17個半胱氨酸,說明該蛋白可能具有二硫鍵。它沒有Pyl(O)Sec(U)(B)(Z)(X)氨基酸。該FAS的氨基酸組成分別為:Ala(10.6%),Arg(4.5%),Asn(3.0%),Asp(5.0%),Cys(0.6%),Gln(4.0%),Glu(6.5%),Gly(8.5%),His(1.9%),Ile(5.4%),Leu(8.9%),Lys(6.4%),Met(2.3%),Phe(3.5%),Pro(4.9%),Ser(6.7%),Thr(6.1%),Trp(1.1%),Tyr(3.1%)以及Val(7.0%)。
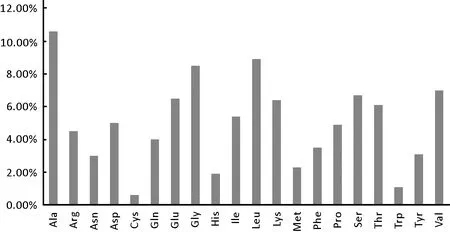
圖2 圓紅冬孢酵母FAS的氨基酸組成分布圖
2.3 蛋白質的理化性質
利用在線軟件ProtParam對FAS理化性質的各個參數進行初步預測分析,得到結果如下:該蛋白的原子構成為C14186H22484N3858O4275S84,原子總數44887,相對理論分子量為318178.90道爾頓,理論等電點pI值為6.39。對于該蛋白的摩爾消光系數(Molar Extinction Coefficient),以M-1cm-1為單位,在水中280nm處測定。假設該FAS所有的胱氨酸殘基以半胱氨酸的形式出現,即形成二硫鍵,此時該蛋白的消光系數值為312590L/(mol·cm),0.1%濃度(1g/L)的吸光度(A280)為0.982;假設二硫鍵全部打開,該蛋白的消光系數值為311590L/(mol·cm),0.1%濃度(1g/L)的吸光度(A280)為0.979[6]。當其成熟肽N端為Met甲硫氨酸時,FAS的估計半衰期在體外哺乳動物網織紅細胞為30h,在酵母體內半衰期大于20h,在大腸桿菌體內半衰期大于10h。該蛋白的不穩定系數為38.47,可知該FAS為穩定蛋白(定義當蛋白質的穩定系數<閾值40時,其在溶液中性質穩定)。該蛋白脂肪指數為86.64,親水性為-0.199。對于蛋白質的疏水性/親水性,定義為氨基酸序列中全部氨基酸的親水值總和與氨基酸數量的比值。負值越大親水性越好,正值越大疏水性越強。該FAS親水性負值,表明此蛋白為親水性蛋白。
2.4 疏水性分析
蛋白質疏水性/親水性的預測和分析是進行蛋白質二級結構預測及功能域劃分的一個必要過程[6]。運用在線軟件ProtScale來預測該FAS氨基酸序列的疏水性/親水性。由圖3得預測結果:該FAS多肽鏈中,第2316位的氨基酸Arg(R)具有最低分值-3.178,第1919位的氨基酸Gly(G)具有最高分值2.400。依據氨基酸分值越低親水性越強,分值越高疏水性越強的規律,可以看出第2316位的Arg親水性較強,第1919位的Gly疏水性較強。整個肽鏈中親水性氨基酸多于疏水性氨基酸,因此整個多肽鏈表現為親水性,可判斷該FAS是親水性蛋白。
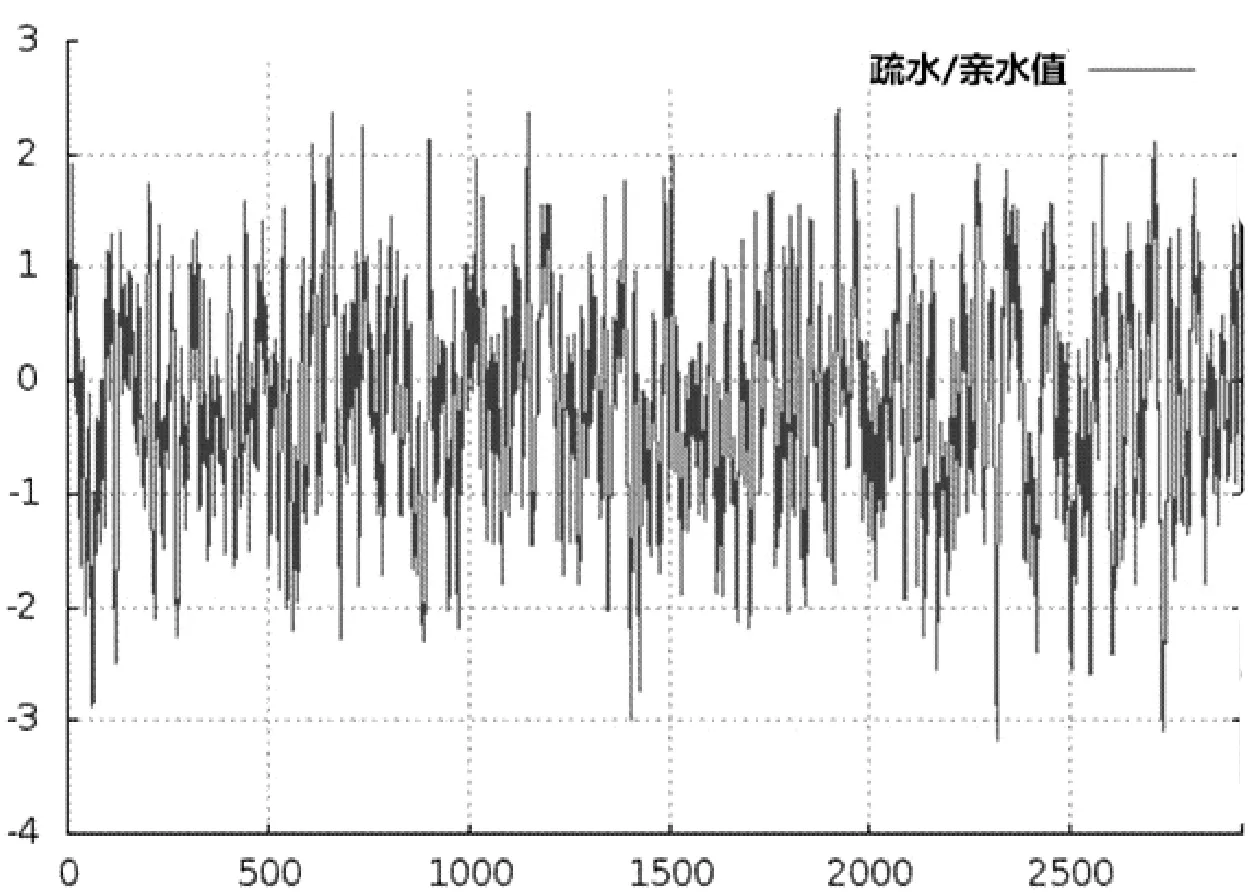
圖3 FAS疏水性預測分析結果
2.5 FAS跨膜預測
用TMpred工具對該FAS氨基酸序列的跨膜結構域進行預測。橫軸為氨基酸位置,縱軸為跨膜預測得分值,定義預測分值低于500,顯示不具有跨膜結構。從圖4中可得,對該FAS氨基酸序列跨膜的預測分值有多個超出500,因此推測該蛋白具有跨膜結構。
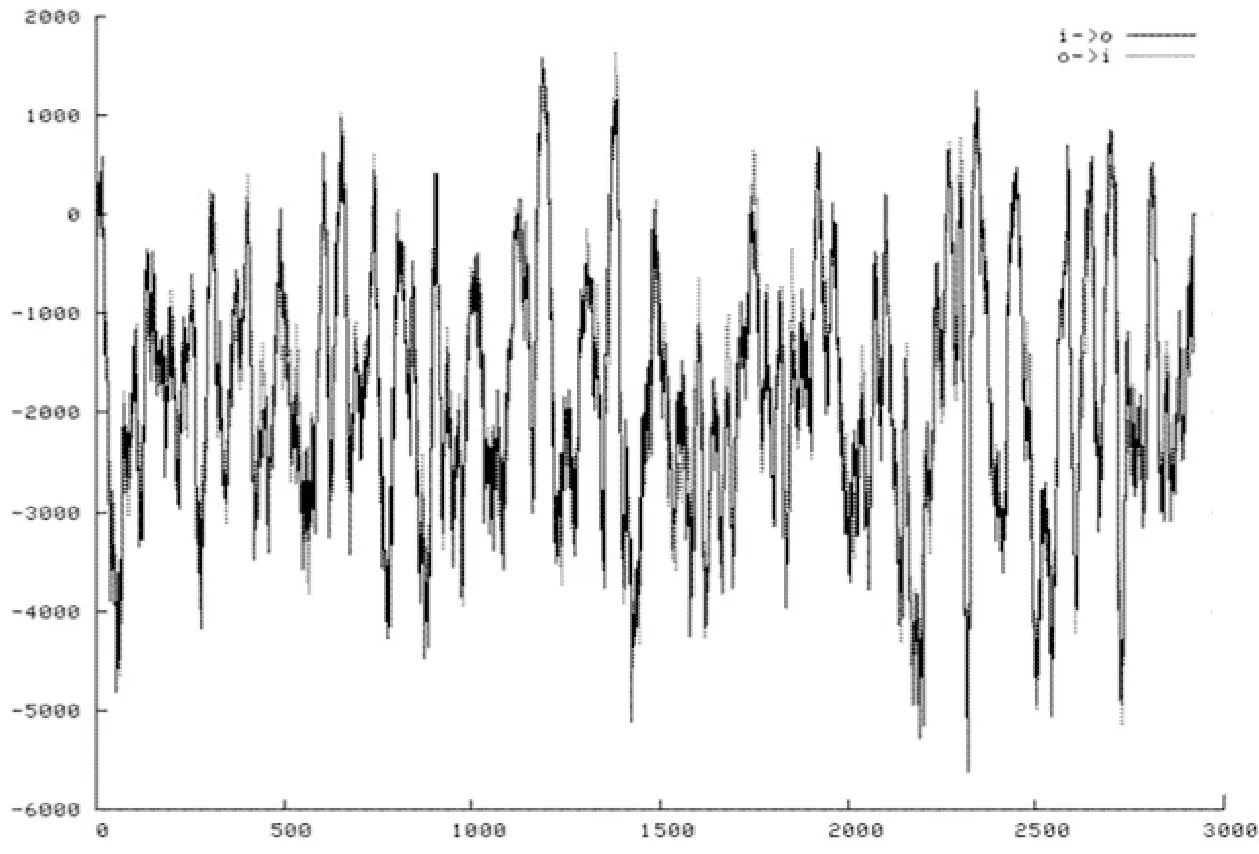
圖4 FAS跨膜預測結果
2.6 信號肽分析
使用SignalP對該FAS進行信號肽預測,得出該FAS的Signal peptide probability預測值為0.167,小于臨界值,未發現信號肽序列(圖5),因此推算該FAS為非分泌性蛋白(Non-secretory protein)。
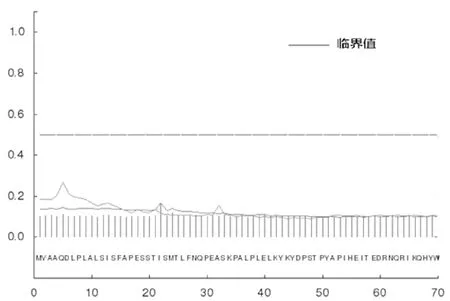
圖5 FAS信號肽預測圖
2.7 FAS核輸出信號分析
使用PredictNLS進行核輸出信號預測。從圖6中可得該蛋白的核輸出信號(nuclear expotr-signal,NES)分值中有超過閾值的,因此可以認為該蛋白質具有核輸出信號。
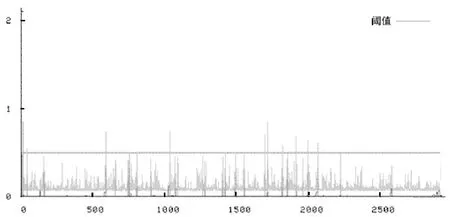
圖6 FAS的核輸出信號預測結果
2.8 蛋白質的二級結構分析
對該FAS的二級結構進行預測,結果顯示該FAS可能包含的二級結構及各個數據分別為α-螺旋1302個,占44.47%;β-折疊418個,占14.28%;β-轉角184個,占6.28%;無規則卷曲1024個,占34.97%。α-螺旋及無規則卷曲是FAS二級結構中最大量的結構元件,而β-折疊和β-轉角則散布于整個蛋白質中。
2.9 蛋白質的結構域分析
利用SMART軟件在線預測該FAS的結構域,分別是位于第87到第210位氨基酸以及第364到第480位氨基酸的脫水酶(Dehydratase)結構域,位于第495到第872位氨基酸的乙酰轉移酶(Acetyl transferase)結構域,位于第2218到第2444位氨基酸以及第2541到第2662位氨基酸的酮酰合酶(Ketoacyl synthase)結構域,位于第2812到第2924位氨基酸的4’-磷酸泛酰巰基乙胺基轉移酶(Phosphopantetheinyl transferase,PPTase)結構域。
3 討論
脂類作為生化大分子的一員,可以說它已經不僅僅是組成微生物細胞膜的重要部分,還是生物體細胞內許多重要生化反應的參與者和調節者。它構成了細胞的完整性并參與了免疫防御機制[7,8]。
生物油脂作為一種生活中的保健食品,其反應作用已經有較多學者進行研究。隨著不斷出現的各種多不飽和脂肪酸(GLA,EPA,DHA,AA)在抗腫瘤、抗衰老、降血脂、降糖等方面作用的逐步研究,微生物油脂的應用有十分廣闊的市場和值得期待的前景[9]。
在對新能源的開發中,生物柴油作為主要能源備受矚目。它作為與現有石化柴油非常相似的能源,是將甘油三酯轉脂化得到的脂肪酸甲脂(或乙脂),其原料可再生產,且屬于清潔能源,成本上也不用經常更換和清洗發動機,其分析方法也多種多樣,因此受到廣泛關注[10]。目前生產生物柴油的原料主要為植物油或廢棄食物油,其生產依賴植物油脂資源。
在適宜條件下,某些微生物可在體內積累單細胞油脂,例如細菌、酵母和藻類等,尤其是產油脂酵母生長速率快,油脂含量高[11]。圓紅冬孢酵母產生的油脂具有與植物油類似的脂肪酸組成,是生物柴油生產的潛在替代品。它作為脂滴細胞生物學和可持續化學生產的模型具有巨大潛力。利用生物發酵的方法生產脂肪酸及其衍生物被認為是一種可再生且環境友好的生產模式[12]。
本實驗對圓紅冬孢酵母油脂代謝生化反應中重要的脂肪酸合酶進行生物信息學分析,通過研究該蛋白的理化性質及結構特征,以便預測其在油脂代謝中所起到的作用,并為下一步構建產油工程菌奠定基礎。
猜你喜歡
食品工業(2023年12期)2023-12-12 12:56:16
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
瘋狂英語·新讀寫(2020年3期)2020-06-06 09:06:14
航空世界(2020年10期)2020-01-19 14:36:20
中國糧油學報(2019年4期)2019-07-12 09:06:38
食品與機械(2018年11期)2019-01-03 06:15:34
