黔中地區馬尾松林下杜鵑根部內生真菌群落組成及其生態功能
2020-04-16 05:40:38李林盼楊舒婷何躍軍李朝嬋
生態學報 2020年4期
熊 丹,歐 靜,*,李林盼,楊舒婷,何躍軍,李朝嬋
1 貴州大學林學院, 貴陽 550025 2 貴州師范大學貴州省山地環境重點實驗室, 貴陽 550001
真菌多樣性是生物多樣性的重要組成部分,主要包括形態多樣性、物種多樣性和功能多樣性,有研究表明菌根真菌能提高苗木移栽成活率,研究其多樣性可以為荒山改造、水土保持、增加森林覆蓋率等做貢獻,因而具有重大意義[1- 2]。杜鵑(Rhododendronsimsii)生于海拔500—1200(-2500)m的山地疏灌叢或松林下[3],素有“花中此物最西施”的美譽,具有重要的觀賞價值和園林應用價值[4]。植物與土壤中的真菌互作形成根系共生體(Root-associated fungi, RAF)[5],真菌通過分解有機物質為宿主植物提供必須的營養[6- 7],可促進種子的萌發[8]、營養物質的吸收[9]、幼苗的生長與發育[10],增加對水分脅迫的耐受性和光合效率,進而提高其對惡劣生境的適應能力[11- 12]。杜鵑屬植物具有豐富的真菌多樣性[13- 14],Heinonsalo等[15]對同一生境下的杜鵑屬植物菌根(Ericoid mycorrhizal fungi,ERM)及伴生植物菌根進行比較研究,得出ERM在其他植物根部也能形成,共生植物菌根之間存在關聯性。以往對ERM的研究多從分離鑒定其菌根菌后回接到宿主植物,通過觀察其對宿主生理生態的影響分析其功能性,故而忽略了杜鵑根部不能分離的未知真菌類群,分離真菌回接到宿主具有隨機性和盲目性,工作量大且研究周期長。相關報道發現直接分子檢測法能較為全面地反映杜鵑屬植物RAF多樣性[16],FUNGuild 注釋軟件可不依賴于培養的分子檢測技術,根據不同類型真菌能以相似方式利用相同類型營養物質為劃分依據,借助生態功能群從分類學上解析真菌的生態類型[17],近年來以 Venn 圖中重疊的圓圈部分表示核心微生物組為微生物群落的研究提供了新的思路[18- 19],而鮮有關于馬尾松林下杜鵑RAF群落組成和生態功能結構分析的報道,其核心基因組真菌研究更是罕見。
本文基于高通量測序技術,從真菌分類學和功能多樣性角度對黔中3個地區馬尾松林下杜鵑根內生真菌進行多樣性分析,在系統了解菌群結構組成的基礎上對真菌功能多樣性進行深入探討,分析生活在馬尾松林下的杜鵑根內生菌中是否存在具標記性的核心真菌組(core microbiome) 及關鍵物種(keystone species),并以網絡圖的形式展現不同菌群之間的相關性,初步探討了黔中三個地區杜鵑根部真菌的群落組成及生態功能群二者間的聯系,有助于揭示宿主植物的環境適應性機制,為今后開展杜鵑菌根真菌在杜鵑人工繁育中的應用奠定基礎,同時為園林上將杜鵑與馬尾松配植提供數據參考。
1 研究方法
1.1 研究地概況
選取黔中貴陽市花溪區孟關鄉(N26.840, E106.989,海拔1270m)、烏當區百宜鎮拐比村(N26.415, E106.749,海拔1196m)和貴州省黔南龍里縣龍架山森林公園(N26.465, E106.937,海拔1182m)為試驗樣地。研究區植物群落從上到下依次為喬木層、灌木層、草本層。其中馬尾松-杜鵑為樣地常見植物群落,馬尾松樹高15—20 m,胸徑25—35 cm;杜鵑高1.75—2.40 m,地徑2.5—3.2 cm。此外,樣地伴生植物為:白櫟(QuercusfabriHance)、茅栗(CastaneaseguiniiDode)等多種灌木。黔中地區屬亞熱帶季風性濕潤氣候,土壤以山地黃壤為主,呈酸性,土質粘重。年均溫約15.3 ℃,年降雨量高于870 mm,平均相對空氣濕度79.9%,日照百分率25.7%,無霜期200 d以上。
1.2 試驗方法
1.2.1樣品采集
于2017年5月15日、17日、20日分別在貴州省黔南龍里龍架山森林公園(LL),貴陽市烏當區百宜鎮拐比村(WD)、花溪區孟關鄉(MG)選取馬尾松-杜鵑生長良好且杜鵑長勢相似的群落,在不破壞生態環境的情況下,去除杜鵑垂直蓋度以下的枯枝落葉層和雜物,由主干追蹤到側根和發根區域,挖出5—20 cm的杜鵑根段主要分布層,剪取粗壯發根(直徑0.5—1.5mm,樣品0.5g)用作杜鵑根部真菌群落組成分析,取樣后覆土。每個地區選擇三個樣方采集3個樣品,每個樣品之間間隔超過5 m,試驗一共9個樣品。試驗工具在采樣之前均經過嚴格滅菌處理,樣本采集后立刻用液氮處理,帶回后保存在-20 ℃冰箱中備用。
1.2.2根樣中真菌基因組DNA提取、擴增及測序
采用Illumina MiSeq二代測序平臺對樣本DNA片段進行雙端(Paired-end)測序(由上海派森諾生物測序公司完成)。利用試劑盒omega EZNA soil extration kit提取杜鵑菌根真菌DNA,操作步驟如下:
(1)將預處理好的根樣放到對應編號且預裝有500mg glassbeads的2mL離心管中;
(2)向上述樣品管中加入0.8mL Buffer SLX Mlus.于組織破碎儀25HZ振蕩7min;
(3)加入80μL Buffer DS,振蕩混勻;
(4) 90℃孵育10min,孵育期間顛倒離心管以混勻其中液體1—2次(因不知樣品中是否含有難裂解的真菌,故將孵育溫度提高至90℃);
(5)13000×g(12000rpm)室溫離心5min,轉移600μL上清至新的96深孔板中,加入200μL Buffer SP2,用手輕輕振蕩混勻,加入100μL HTR Reagent;
(6)冰浴5min,4000×g(3700rpm)離心10min;
(7)轉移上清400μL至96深孔板中,加入40μL磁珠,450μL Binding Buffer,混勻,室溫放置2min;
(8)上核酸純化儀進行純化。
利用0.8% 的凝膠電泳和紫外分光光度計檢測DNA的質量和濃度。檢測合格后,利用真菌rDNA-ITS保守區上游引物:5′-GGAAGTAAAAGTCGTAACAAGG- 3′和下游引物:5′-GCTGCGTTCTTCATCGATGC- 3′[20],進行特異性擴增,PCR反應程序為:98 ℃預熱變性2 min;25個循環,每個循環98 ℃,熱變性15 s,55 ℃退火30 s,72 ℃延伸30 s;最后72 ℃ 延伸5 min,10 ℃保存。將擴增產物進行回收、熒光定量,回收采用AXYGEN公司的凝膠回收試劑盒。采用Illumina公司的TruSeq Nano DNA LT Library Prep Kit制備測序文庫,文庫制備合格后上機測序,測序得到的基因序列提交至GenBank數據庫。登錄號PRJNA505090,編號為SAMN10386261-SAMN10386263,SAMN10386267-SAMN10386269,SAMN10386273-SAMN10386275。
獲得原始數據后,對測序質量進行評估和質控,利用QIIME軟件將分類單元OTU (Operational Taxonomic Units)歸并和聚類(相似度閾值為97%)[21],采用UNITE數據庫[22]進行OTU分類注釋;獲得OTU豐度矩陣之后,計算每個樣本真菌群落的Alpha多樣性。
1.2.3杜鵑根內生真菌多樣性檢測和功能分析
生態功能群分析:選取菌群豐度大于10的OTU序列,采用FUNGuild在線數據庫平臺(https://github.-com/UMNFuN/ FUNGuild)[17],將杜鵑根部內生菌劃到不同的營養型(Trophic mode)和共位群(Guild)。營養型包括共生型(Symbiotroph)、腐生型(Saprotroph)、病原型(Pathotroph);共位群包括杜鵑菌根真菌、外生菌根真菌、叢枝菌根真菌、蘭科植物菌根真菌、內生真菌、病原真菌、腐生真菌等[23- 24]。
關聯網絡圖分析:根據真菌在各樣本中的分布,選取豐度大于20的物種,調用R語言vegan、Igraph、psych、pheatmap、magrittr、ggplot2、RColorBrewer、reshape2、cluster包,對菌群進行關聯網絡圖分析。
2 結果
2.1 杜鵑根內生真菌的群落結構
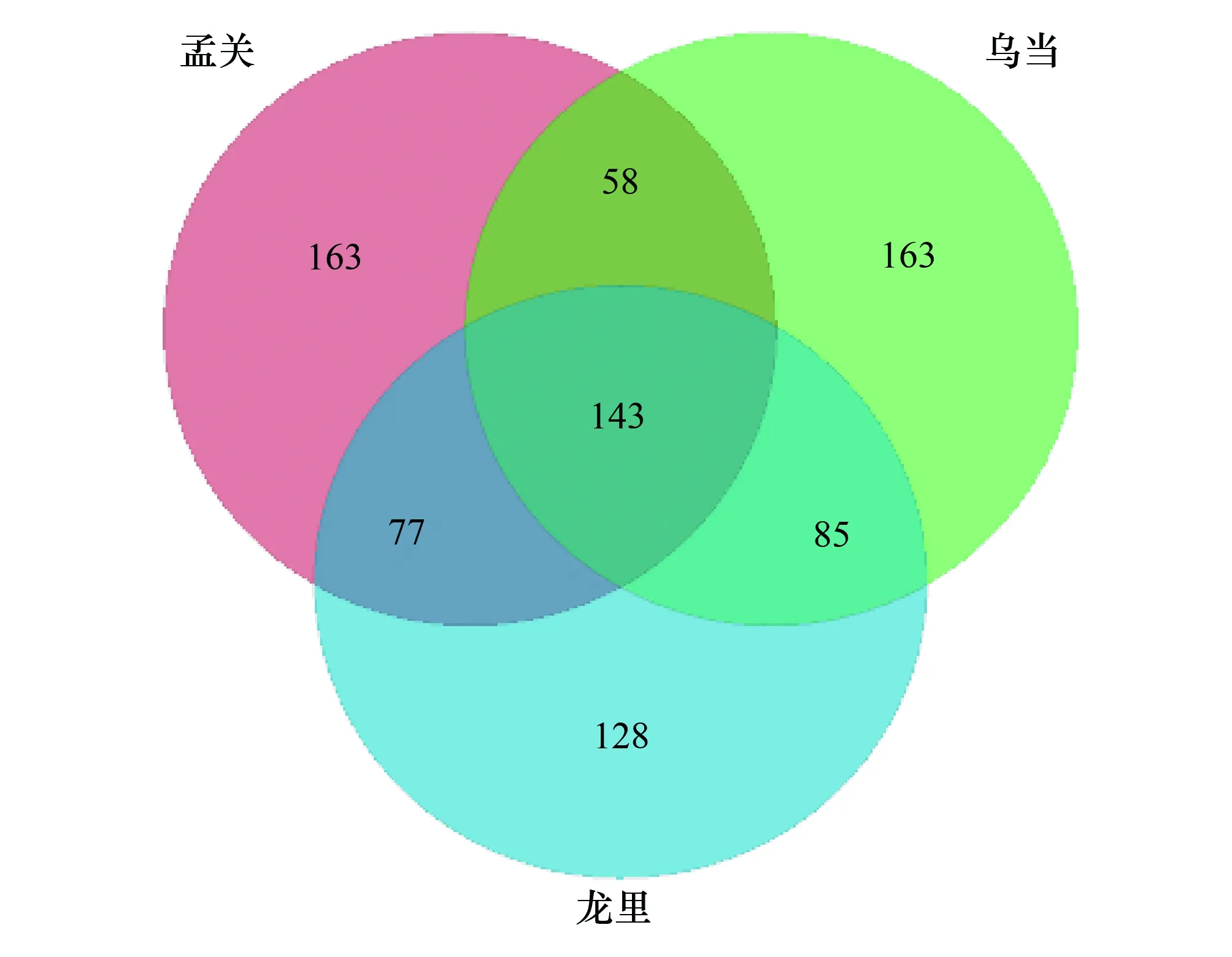
圖1 共有分類單元(OTU)的Venn圖Fig.1 Venn diagrams of OTU
通過高通量測序方法對黔中烏當(WD)、孟關(MG)、龍里(LL)三個地區的杜鵑根內生真菌群落組成分析,共獲得有效序列425799條,按照97%的相似閾值對有效序列進行聚類,共獲得817個OTU,韋恩圖(圖1)顯示,MG真菌OTU總數441,WD為449,LL為433,共有真菌OTU數143,表明三個地區杜鵑根部內生菌OTU值數量差異較小。
經聚類,817個OTU分屬于8門、18綱、53目、105科、152屬,優勢菌群系為子囊菌門(81.2%)、接合菌門(5.8%)、擔子菌門(4.7%);優勢菌綱主要為散囊菌綱(43.0%)、糞殼菌綱(18.9%)、錘舌菌綱(7.0%)等;優勢菌目為散囊菌目(39.2%)、肉座菌目(9.1%)、Xylariales(8.4%)、柔膜菌目(6.1%)等;優勢科有發菌科(39.2%) 、圓孔殼科(8.4%)、肉座菌科(7.2%)等;152屬中相對豐度大于0.01%的真菌共有30個屬,主要是青霉屬(38.60%) 、木霉屬(7.20%)、擬盤多毛孢屬(6.10%)、Umbelopsis4.40%等(見圖2)。
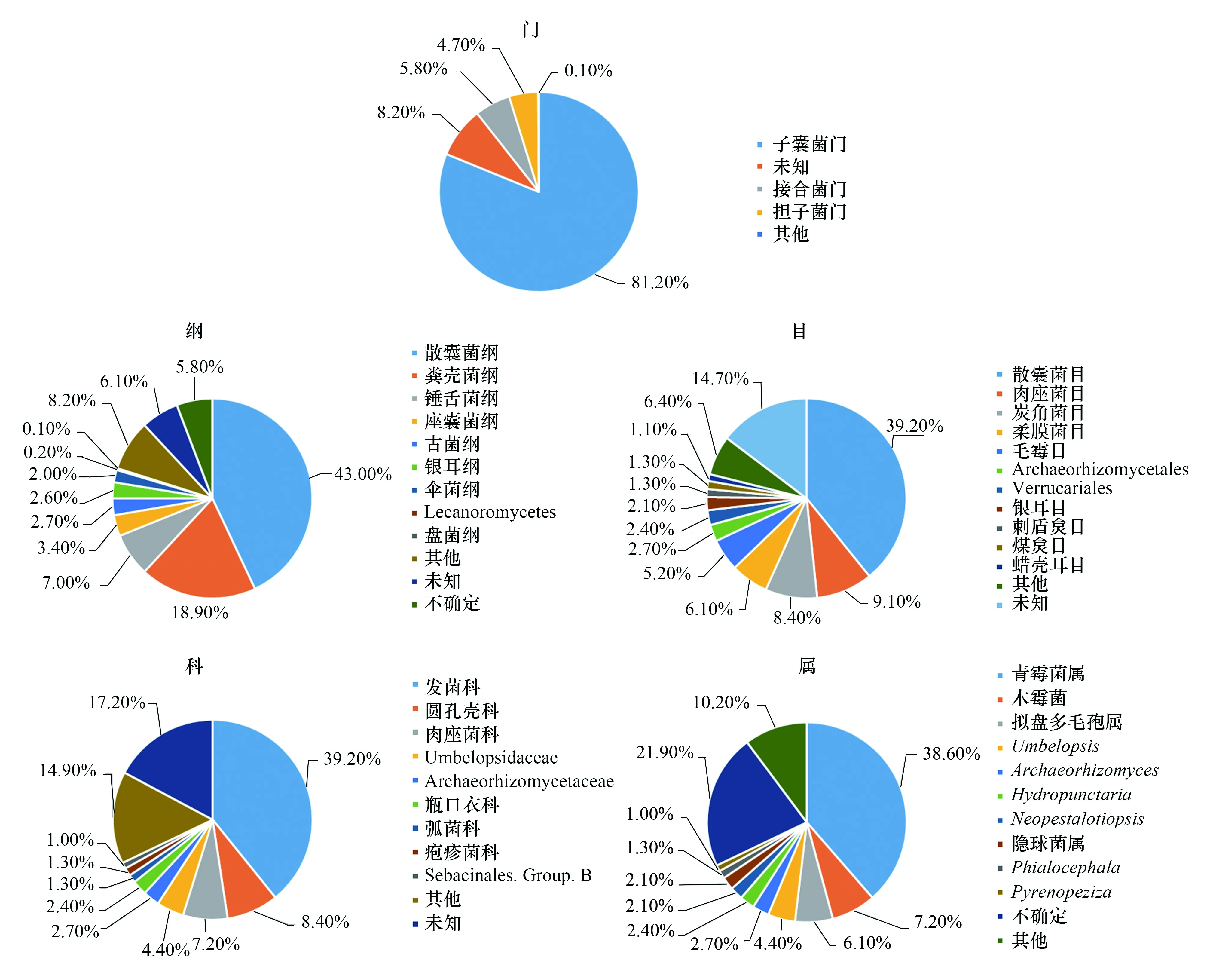
圖2 杜鵑根內生真菌群系組成Fig.2 Composition of endophytic fungi in roots of Rhododendron simsii
2.2 不同區系優勢菌群
孟關(MG)地區杜鵑根內生優勢真菌為子囊菌門Ascomycota的青霉菌屬Penicillium、擬盤多毛孢屬Pestalotiopsis、木霉屬Trichoderma、新擬水蚤屬Neopestalotiopsis和一未知屬,接合菌門Zygomycota傘狀霉屬Umbelopsis,擔子菌門Basidiomycota隱球菌屬Cryptococcus;烏當(WD)地區優勢真菌組成為子囊菌門的青霉菌屬、Hydropunctaria、木霉屬、Chloridium、Neopestalotiopsis、隱球菌屬Cryptococcus,接合菌門的Umbelopsis;龍里(LL)地區的優勢真菌子囊菌門的青霉菌屬、擬盤多毛孢屬、木霉屬、Archaeorhizomyces、Phialocephala、Chloridium、Neopestalotiopsis、Cryptococcus,接合菌門的Umbelopsis,擔子菌門傘菌綱Agaricomycetes的未定屬。三個地區杜鵑根部真菌具有較高的多樣性,真菌之間既有相似又有差異,其中包括了典型的ERM樹粉孢屬Oidiodendron(0.4%),ECM紅菇屬Russula(0.2%)、乳菇屬Lactarius(0.2%)及其他類型的菌根菌。而豐度最高的是青霉菌屬、木霉屬和擬盤多毛孢屬,在杜鵑根部均有分布,表現出一定的寄主專一性。杜鵑根部內生真菌類型豐富,但杜鵑類菌根菌(ERM)占比相對較低。
2.3 真菌的Alpha多樣性
通過R軟件,對屬水平的真菌群落組成結構進行PCA分析,第一和第二主軸的累積方差貢獻率為83.46%,其中第一主軸解釋真菌屬水平組成方差變異的72.58%,第二主軸解釋10.88%,不同地區之間菌群組成差異較大(圖3)。3 個樣地的 α 多樣性指數見表1,Simpson指數、Shannon指數LL>WD>MG,LL Simpson指數顯著高于MG(P<0.05),LL與WD、WD與MG之間不存在顯著差異,而Shannon指數WD、LL顯著高于MG(P<0.05),WD、LL之間不存在顯著差異。ACE指數、Chao1指數WD>MG>LL,Chao1指數WD顯著高于MG和LL(P<0.05),MG和LL不存在顯著差異。可見,WD地區真菌多樣性最大,豐富度也最高,LL地區真菌多樣性次之,MG豐富度略高于LL,但不存在顯著差異。馬尾松林下杜鵑根部內生真菌OTU數量差異值較小,但組成上存在較大差異。
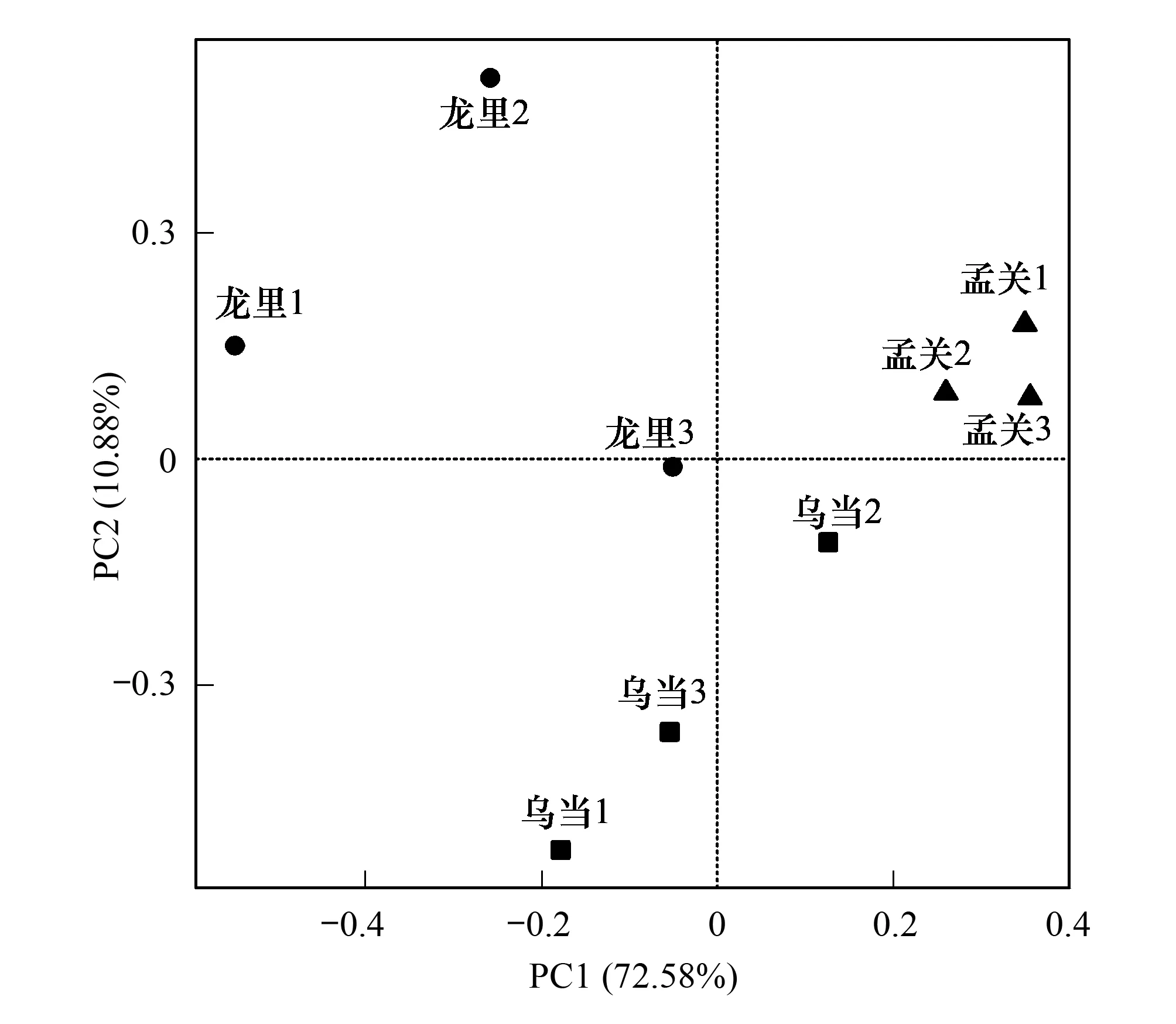
圖3 屬水平的群落組成結構PCA分析Fig.3 PCA analysis of community composition at genus level

表1 杜鵑根內生真菌Alpha多樣性指數表
同列數據后不同大寫字母表示差異顯著(P<0.05)
2.4 杜鵑根內生菌的生態功能分析
基于FUNGuild軟件解析結果顯示(表2),除未知菌群外杜鵑根內生真菌主要分為10個生態功能群,分別是未定義腐生菌Undefined Saprotroph (194 OTU)、植物病原菌Plant Pathogen (20 OTU)、土壤腐生菌Soil Saprotroph (18 OTU)、外生菌Ectomycorrhizal (14 OTU)、地衣共生真菌Lichenized fungi (10 OTU)、杜鵑類菌根真菌Ericoid Mycorrhizal (5 OTU)、木腐生菌Wood Saprotroph (5 OTU)、叢枝菌根Arbuscular Mycorrhizal (4 OTU)、內生菌Endophyte (2 OTU)和動物病原菌Animal Pathogen (8 OTU) 10類。此外還有多種混合營養型的真菌如:寄生性真菌-不定腐生菌Fungal Parasite-Undefined Saprotroph (11 OTU);植物病原-土壤腐生-木材腐生菌Plant Pathogen-Soil Saprotroph-Wood Saprotroph (8 OTU);植物內生-病原菌Endophyte-Plant Pathogen (2 OTU)等21類,102個Undefined種類在FUNGuild數據庫中沒有參考信息。
2.5 根內生真菌核心真菌組及關鍵物種分析
不同類型的真菌行使不同的功能,推測存在著重要的功能菌群,為此,采用R語言對杜鵑根部內生菌功能群做關聯網絡分析,獲得的有效功能類別關聯圖結構為8大部分,包含265個真菌OTU,3357條關系,不同顏色代表不同關聯性的菌群,OTU所在網絡圖中的圓球越大,則競爭能力越大,之間的連線越粗代表影響力和關聯性也越高(圖3)。結合三個地區杜鵑根內生共有真菌Venn圖發現,共有屬中有19個分類單元 otu在共有功能群中分布頻率較高,分別是otu3717青霉屬(Penicillium)、otu1062木霉屬(Trichoderma)、otu0青霉屬(Penicillium)、otu1580擬盤多毛孢屬(Pestalotiopsis)、otu2468(Chloridium)、隱球菌屬(Cryptococcus)、otu108銀耳目(Tremellales)、otu807(Hydropunctaria)、otu2049毛孢子菌屬(Trichosporon)、otu2592樹粉孢屬(Oidiodendron)、otu2115短梗霉屬(Aureobasidium)、otu376短梗霉屬(Aureobasidium)、otu1537(Phialocephala)、otu3296支頂孢屬(Acremonium)、otu3433假裸囊菌屬(Pseudogymnoascus)、otu1800(Purpureocillium)、otu2492腐質霉屬(Humicola)、otu1787(unidentified)、otu313(unidentified)。這些核心真菌在不同地區的杜鵑根部均有出現,其生態類型見表2,從功能網絡圖分析看出,不同生態型真菌之間存在關聯性,根部真菌可以形成生態位共享模式,而且不同功能群之間存在耦合性,功能大小不一。功能團之間包含有多種類型的菌群結構,在時間和空間上相互聯系,推測這是多個OTU在微生物組內功能冗余的表現形式,核心基因組與關鍵物種以真菌組形成的生態功能團表現。
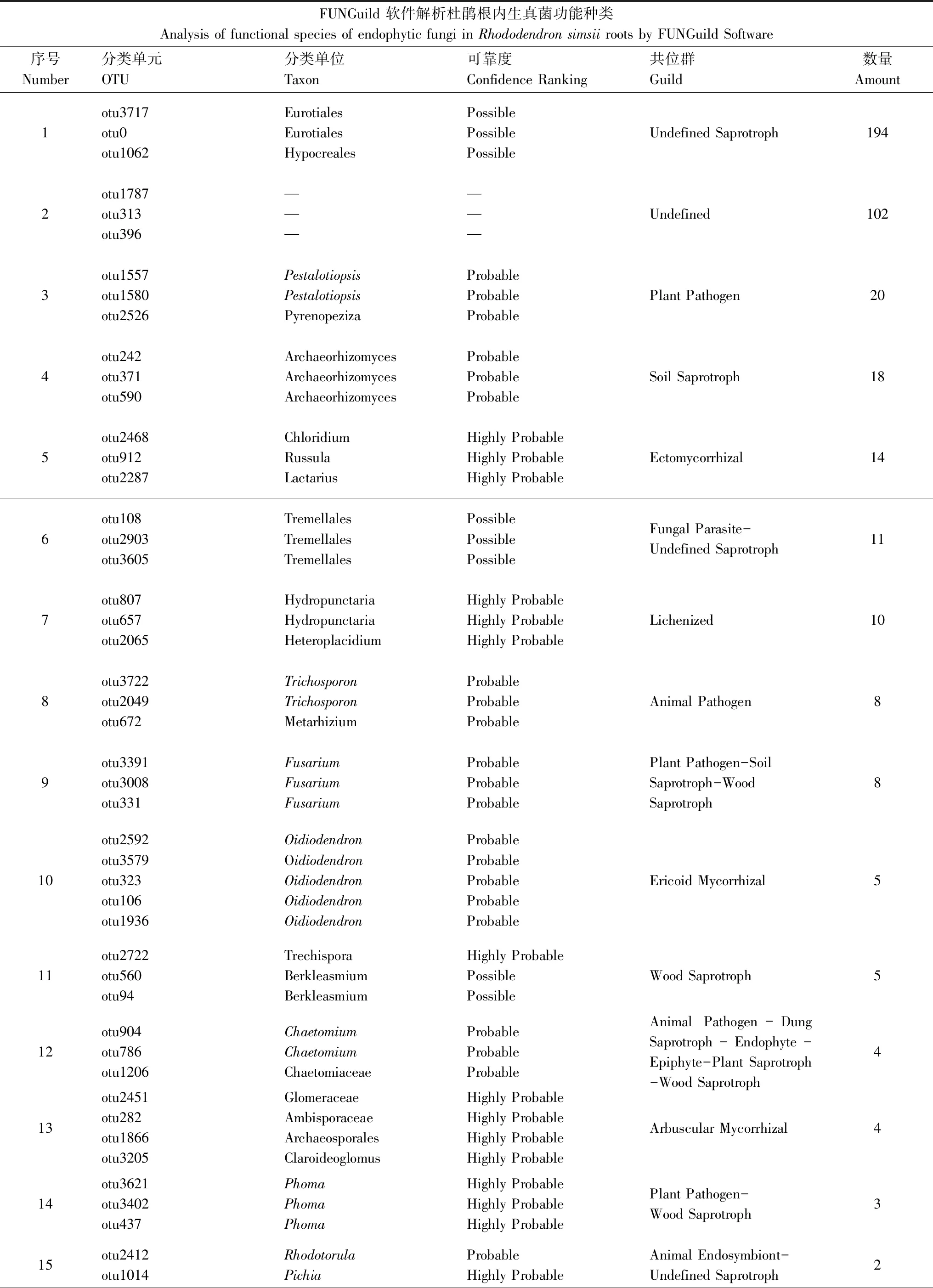
表2 杜鵑根內生真菌功能解析

圖4 杜鵑根內生真菌OTU生態功能群的共存網絡圖Fig.4 Coexistence Network of Endophytic Fungi OTU in Rhododendron simsii RootsAnimal pat和Plant pat 表示動物病原菌和植物病原菌,Und.sap 表示未知腐生菌 (圖中只顯示部分OTU名稱)
3 討論
3.1 杜鵑根內生真菌的群落多樣性
根部真菌(RAF)多樣性決定了植物的多樣性、生態系統的變異性和生產量[25]。高通量測序獲得杜鵑根內生真菌DNA序列425799條,聚類后得到的817個OTU分屬于8門、18綱、53目、105科、152屬。主要隸屬于子囊菌門(81.2%)、接合菌門(5.8%)和擔子菌門(4.7%);優勢綱、目、科分別為:散囊菌綱(43.0%)、散囊菌目(39.2%)、發菌科(39.2%);在屬的水平上,青霉屬(38.60%)占比最高,其次是木霉屬(7.20%)、擬盤多毛孢屬(6.10%)。
杜鵑RAF中有典型的樹粉孢屬Oidiodendron(ERM),紅菇屬Russula(ECM)、乳菇屬Lactarius及其他類型的菌根菌。研究發現的杜鵑RAF已有大量報道,如青霉屬、紅菇屬、蠟殼耳屬、隱球菌屬等在不同林型馬銀花(R.ovatum)根部均有分布[26],臺灣杜鵑(R.formosan)[7]、樹楓杜鵑(R.changii)和云南大理灰背杜鵑(R.hippophaeoides)[27]菌根中均有隱球菌屬。不同生態類型的菌根真菌在杜鵑根部共生,其形成原因可能是共生植物之間菌根存在著關聯性,馬尾松林中不同植物地下部分菌根能形成關聯網絡。子囊菌門柔膜菌目和擔子菌門蠟殼耳目是典型的杜鵑花類植物菌根菌,張春英等[14]、劉仁陽等[28]也分別從云錦杜鵑(R.fortunei)和雷山杜鵑(R.leishanicum)根中分離到此類真菌。相比于黃彩微等[16]從薄葉馬銀花(R.leptothrium)根系檢測到的8個OTU 和銹紅杜鵑(R.bureavii)根系的7個OTU,黔中馬尾松林下生長的杜鵑根內生真菌豐富度和多樣性極高,OTU數量大于來自于我國長白山(355 OTU)、大興安嶺(229 OTU)和小興安嶺(226 OTU)地區的越桔(Vacciniumuliginosum)[29]根部真菌。Alpha多樣性分析顯示, WD地區真菌多樣性最大,豐富度也最高,LL地區真菌多樣性次之,MG豐富度略高于LL,但不存在差異顯著。導致這一現象的原因可能是受杜鵑自身生長周期與生境因素的影響,三個區系杜鵑處于不同小生境下,內生真菌菌群的分布及組成也會有所不同。植物內生真菌菌群在宿主植物中的組成及生態分布特征往往會隨著宿主樹齡、組織類型、營養成分、生長季節、溫度升降、降水量多寡等眾多因素的變化而變化[30- 31]。受植物種、植物群落和環境因子的相互作用,植物組成接近的生態群落,其環境因子組成上也比較接近,但生境差異促進了菌群數量和結構的多樣化[32]。筆者所在課題組已對黔中不同片區馬尾松林植物多樣性和杜鵑根部土壤的理化性質進行分析,探討植物多樣性和土壤理化性質對馬尾松-杜鵑混交林真菌群落結構的影響(論文待發表)。
3.2 根內生真菌的生態功能多樣性
菌群中真菌的群落組成對宿主和環境的影響一直是該領域的熱點,微生物群落組成和豐度對生態系統有重要的調節功能,核心真菌組與關鍵物種的貢獻也成為研究的重要部分。FUNGuild 軟件解析菌群結果顯示,otu2592、otu3579、otu323、otu106、otu1936同屬于ERM,生長形態學(Growth Morphology)屬于黑色有隔內生菌(Dark Separate Endophytes, DSE),DSE廣泛存在于高山和極地的植物根部,能增強植物對P的吸收和獲取[16,33]。外生菌根(Ectomycorrhizal,ECM)功能群包含有otu2468、otu912、otu2287等14個OTU,它 們 分 別 是Chloridium、Russula、Lactarius等[34],ECM多為子囊菌門和擔子菌門真菌,能夠在溫帶和熱帶生態系統中與廣泛的宿主建立共生關系,常與松科(Pinaceae)、樺木科(Betulaceae)、豆科(Fabaceae)、龍腦香科(Dipterocarpaceae)、殼斗科(Fagaceae)和桃金娘科(Myrtaceae)等高等植物形成菌根[35- 36]。以子囊菌門為主的腐生型真菌在杜鵑RAF群落中也較常見,如未定義腐生菌群(Undefined Saprotroph 194 OTU),其中otu3717、otu0、otu1062分別是Penicilliumabidjanum,Penicilliumspinulosum,Trichodermaatroviride,這三類真菌出現在有云杉屬(Picea)、歐洲赤松(PinussylvestrisL.)和杜鵑屬(RhododendronL.)共生的瑞典中部混交林中,反映了真菌多樣性與宿主的相關性[37]。土壤腐生菌(Soil Saprotroph)功能群中包含有otu242、otu371、otu590等18個菌群,它們都是古生菌Archaeorhizomycetaceae,普遍存在于松屬、杜鵑屬和其他闊葉樹的根和根際土壤中,對宿主選擇具有專一性并具腐生潛力,其形成的菌根包括數百種復雜且不可識別的絲狀結構,在熱帶潮濕森林中具有豐富的多樣性[38]。
與Nguyen等[17]結論相似,FUNGuild數據庫解析不全面,還需補充完善,Undefined包含otu1787、otu313、otu396等102個OTU,此類真菌在屬水平上分類地位未定,需要進一步探索其真菌資源。本研究中杜鵑為馬尾松林下伴生植物,數據分析顯示其根部內生真菌功能群分類達到32個,傳統的ERM真菌在杜鵑中所占比例不大,而未定義腐生菌群(Undefined Saprotroph)占比最高。研究發現,一些 OTU無法在屬水平上被解析到定義的功能群中;而另一方面被定義的 OTU 則又屬于各種功能群中,FUNGuild主要是基于營養型來對真菌進行功能劃分的,但有的真菌不僅只有一種營養方式,他們在生活史的不同階段變換著營養方式適應環境的改變,所以才有多樣的功能營養型。如本研究中,植物病原-土壤腐生-木材腐生菌功能群(Plant Pathogen-Soil Saprotroph-Wood Saprotroph)的8個OTU均為Fusarium,既是腐生菌也是內生菌或植物病原菌。植物內生真菌在植物的不同生活周期中扮演不同的角色,在發育學上處于不穩定的階段,在植物組織健康生長的時候是內生真菌,但當組織衰老或死亡時它們是腐生真菌,這些具復雜生活史的真菌為應對不同的生存條件而可采取了不同的生存策略[39- 40]。此外,杜鵑根部內生菌中生活著動物病原菌(Animal Pathogen)功能群,Baarlen等[41]把這一現象解釋為一些病原微生物能夠侵染不同生物寄主,在自然界跨界侵染人與動物。RAF可以形成促進、拮抗或競爭的地下生態模式,以及真菌之間的生態位共享[24]。植物內生真菌與宿主植物之間往往是一種動態平衡的協同關系,菌群的群落組成對宿主植物多樣性及生態環境穩定性起著巨大的影響,菌群組成豐度保證了生態功能系統正常運行,其多樣性增加生態系統對外界環境抵抗性,具有非常重要的生態功能[42- 43]。有研究表明,生物多樣性越高,在新的環境條件下,部分多余的生物體可以承擔這種功能的可能性就越高,即生物多樣性的保險效應[44]。因此,與地上生物多樣性類似,真菌多樣性可能會對環境變化下的生態系統穩定性產生直接影響[45],共生植物之間菌根存在著關聯性,增加了森林生態系統的生物多樣性和生態穩定性,核心基因組與關鍵物種的存在說明了真菌與宿主之間的相互選擇關系。菌群功能團包含有多種類型的真菌,在時間和空間上相互聯系,推測這是多個OTU在微生物組內功能冗余的表現形式,核心基因組與關鍵物種以真菌組形成的生態功能團而表現。
4 結論
黔中馬尾松林下杜鵑根部擁有豐富的菌根類群,除了傳統的ERM菌根類型外,還有ECM、AM、ENM等多種類型菌根,生態功能群豐富,其中烏當(WD)地區杜鵑菌根真菌多樣性最大,豐富度也最高。這些功能復雜的菌群耦合,增加了森林生態系統的生物多樣性和生態穩定性,而核心基因組與關鍵物種只能以真菌組形成的生態功能團表現,這些核心真菌類群值得進一步研究其生態意義。
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
保健醫苑(2021年7期)2021-08-13 08:48:02
學生天地(2020年36期)2020-06-09 03:12:30
小學科學(學生版)(2020年5期)2020-05-25 07:11:32
中國科技論壇(2017年7期)2017-07-25 08:49:53
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
領導文萃(2015年4期)2015-02-28 09:19:05
