雅礱江冬麻豆(Salweenia bouffordiana)種群結構與動態特征
2020-04-16 05:40:38張全建段晨松張遠彬
生態學報 2020年4期
楊 彪,張全建,龔 旭,段晨松,張遠彬
1 西南野生動植物資源保護教育部重點實驗室,西華師范大學,南充 637002 2 中國科學院、水利部成都山地災害與環境研究所,成都 610041 3 中國科學院大學,北京 100049
種群應系指一個地區內同屬于一個種的個體組成的群體[1],種群結構是種群最基本的特征,是種群生態學研究的核心問題之一[2- 3]。極小種群是指地理分布狹小的種群,長期受到外界因素的干擾和壓力,其數量小于防止滅絕所需的最低限度[4],極小種群具有剩余種群少、棲息地受限、人為干擾嚴重和絕種風險極高的特征[5]。目前,我國僅有120個極小種群被關注[4- 5],眾多處于瀕危狀態的物種并未獲得保護地位,而其作為生物多樣性的重要組成,一旦滅絕將給人類帶來不可估量的損失。當前,物種滅絕已是全球最嚴重的生態問題之一,直接威脅著人類社會的可持續發展[6],而保護瀕危物種已成為一個緊迫的保護生物學課題[7]。中國作為世界上最大的生物多樣性國家之一,保護中國最受威脅的野生植物物種比以往任何時候都更加迫切。
雅礱江冬麻豆(Salweeniabouffordiana)是豆科冬麻豆屬(Salweenia)植物新種[8],為我國特有種,主要分布于四川省新龍縣境內的雅礱江河谷干燥灌叢和礫石生境中[8]。由于本種的分布范圍極其狹窄,種群數量稀少,根據世界自然保護聯盟條例[9],有學者們將其保護等級定為極度瀕危(Critically Endangered, CR)[8],《中國生物多樣性紅色名錄——高等植物卷》將其定為瀕危(Endangered, EN)物種[10]。由于缺乏對該物種的種群數量、種群結構等方面的研究,不利于后續關于該物種的相關保護策略的制定和行動的開展,需要對其現有種群數量、分布范圍等進行調查與研究。
1 材料與方法
1.1 研究區概況
雅礱江冬麻豆為常綠叢生灌木,高約0.5—2 m,莖直立,被絨毛,與冬麻豆(S.wardii)的差別主要在于本種的小葉更大、瓣膜呈波浪狀,主要分布于四川省新龍縣境內的雅礱江河谷。本研究的研究區位于四川省甘孜藏族自治州新龍縣,地理位置介于東經99°37′—100°54′E、30°23′—31°32′N之間,海拔2760—3232 m,地處雅礱江中游高山峽谷地帶。該區屬青藏高原型季風氣候,具有獨特的大陸性高原季風氣候的特點。根據新龍縣氣象站多年統計資料可知,全縣平均氣溫為7.5℃,極端最高溫32.7℃,極端最低溫-19.2℃,1月均溫-2.3℃,7月均溫15.1℃,≥10℃活動積溫1887℃,相對濕度在40%左右,全年無霜期114 d。研究區水量分布不均勻,降雨主要集中于每年6月至9月。植被群落主要有河谷灌叢、亞高山暗針葉林、高山松林、高山柏林、櫟類林、樺木林、亞高山草甸、高山草甸等類型。
1.2 調查方法
由于雅礱江冬麻豆主要生長在雅礱江河谷地帶,故沿河岸1 km范圍隨機布設調查樣線8條,沿每條樣線記錄發現的雅礱江冬麻豆、干擾信息和生境信息。在有雅礱江冬麻豆種群分布的地段設置5 m×5 m樣方做常規的種群學調查,共計調查樣方55個。樣方調查時記錄樣方的經緯度、海拔、坡向、坡度等因子,喬木層、灌木層和草本層的植物種類、蓋度和高度等植被狀況,并對雅礱江冬麻豆的種群及生長狀況進行常規種群學調查,記錄樣方內雅礱江冬麻豆的幼苗數量、蓋度、高度、冠幅和地徑等參數。
1.3 計算方法
1.3.1徑級劃分標準
對于木本植物尤其瀕危物種而言,大多數學者認為在數量較少,較難獲取其年齡的情況下,可用徑級結構代替其年齡結構[11]。本研究選擇生物指標特征顯著的地徑結構來代替年齡結構,為分析其種群結構的精細動態,采用李清河等對荒漠珍稀灌木半日花種群的徑級劃分方法[12]進行地徑劃分,即采用較小極差,每1 mm劃分1個徑級。根據地徑(D)的分布狀況,將地徑劃分為25個等級:Ⅰ級(3 mm≤D<4 mm)、Ⅱ級(4 mm≤D<5 mm)、Ⅲ級(5 mm≤D<6 mm)、……、XXIV級(26 mm≤D<27 mm)和XXV級(D≥27 mm)。以上述劃分為標準,統計每個齡級雅礱江冬麻豆個體數,并以此進行種群動態量化分析和建立種群靜態生命表。
1.3.2種群動態量化分析方法
種群動態量化采用陳曉德[13]的量化分析方法定量描述雅礱江冬麻豆種群動態,以Leak[14]的劃分理論作為確定種群結構類型的依據,公式如下:
(1)
(2)
式中,Vn為種群從n到n+1級的個體數量變化;式(2)中Vpi為整個種群結構的數量變化動態指數(忽略外部干擾);Sn、Sn+1分別為第n和n+1級種群個體數;k為種群大小級數量。而當考慮未來的外部干擾時,Vpi還與年齡級數量(k)以及各齡級內的個體數(Sn)相關,可進一步將式(2)修正為:
(3)
(4)
式中,Vn、Vpi、Vpi′取正、負、零值時分別反映種群個體數量的增長、衰退和穩定的結構動態關系。其中,Vpi值越大,說明種群的增長趨勢越大,而Vpi′值越大,說明種群抗干擾能力越大,種群穩定性越好。僅當P(種群對外界干擾所承擔的風險概率)取值為最大時才會對種群動態Vpi′構成最大的影響。
1.3.3種群靜態生命表和生存分析方法
靜態生命表又稱特定時間生命表,多用于對長壽命的木本植物種群的統計研究,主要包括以下參數:Ax是在x齡級內現有個體數;lx是在x齡級開始時標準化存活個體數;dx是從x到x+1齡級間隔期內標準化死亡數;qx是從x到x+1齡級間隔期間死亡率;Lx是從x到x+1齡級間隔期間還存活的個體數;Tx是從x齡級到超過x齡級的個體總數;ex是進入x齡級個體的生命期望或平均期望壽命;Kx為消失率(損失度)。它們之間的計算關系如下:
lx=(ax/a0)×1000
(5)
dx=lx-lx+1
(6)
qx=(dx/lx)×100%
(7)
Lx=(lx+lx+1)/2
(8)
(9)
ex=Tx/lx
(10)
Kx=lnlx-lnlx+1
(11)
由于靜態生命表是用同一時期收集的種群所有個體編制而成,雅礱江冬麻豆種群為天然種群,分析中利用不同齡級結構在空間上的差異來研究特定時間段的種群動態,難免存在系統抽樣誤差產生與數學假設技術不符的現象,但仍能提供有用的生態學記錄[15]。對于這種情況,在編制靜態生命表時常采用勻滑技術對統計數據進行處理,本研究采用方程擬合方法進行勻滑處理[15- 17]。由于第Ⅵ齡級前的植株數存在明顯異常,所以擬合采用第Ⅵ齡級及其之后的數據,以年齡級為自變量,以齡級對應的現存個體數Ax為因變量,經R3.3.3軟件擬合得到方程:y=1473.506x-2.1263(R2=0.7517,P<0.001)。經此擬合方程勻滑修正后得到修正后的ax,然后據此編制出雅礱江冬麻豆種群的靜態生命表。
為了檢驗種群存活狀況是符合Deevey-Ⅱ型還是Deevey-Ⅲ型曲線,本文采用Hett和Loucks提出的數學模型對雅礱江冬麻豆種群存活曲線進行檢驗,即用指數方程Nx=N0e-bx和冪函數方程Nx=N0x-b分別描述Deevey-Ⅱ和Deevey-Ⅲ型存活曲線[18]。式中Nx、N0分別代表x齡級內存活數和種群形成初期的個體數,即Nx、N0為經勻滑后的個體存活數,b為死亡率。
為了更好地分析雅礱江冬麻豆種群動態,闡明其生存規律,采用種群生存率函數(S(i))、累計死亡率函數(F(i))、死亡率密度函數(f(ti))和危險率函數(λ(ti))進行分析[19-20]。計算公式如下:
S(i)=S1×S2×…Si
(12)
F(i)=1-S(i)
(13)
(14)
(15)
式中,Si為存活率,其計算公式為:Si=lx+1/lx,hi為齡級寬度。依據這4 個生存函數的估算值,繪制生存率曲線、累計死亡率曲線、死亡密度曲線與危險率曲線。
1.3.4種群數量動態的時間序列預測方法
本研究采用時間序列分析中的一次移動平均法[21]對雅礱江冬麻豆種群的年齡結構進行預測。
(16)
式中,n表示需要預測的時間(本研究為齡級時間),t為齡級,Xk為k齡級內的個體數量,Mt(1)表示經過未來n個齡級時間后t齡級的種群大小。本文對未來經過2、4、9、6、8、12、20和24齡級時間后的種群各齡級的個體數量進行預測。
2 結果與分析
2.1 雅礱江冬麻豆種群分布現狀和結構特征

圖1 雅礱江冬麻豆種群地徑結構Fig.1 Size structure of S. bouffordiana population
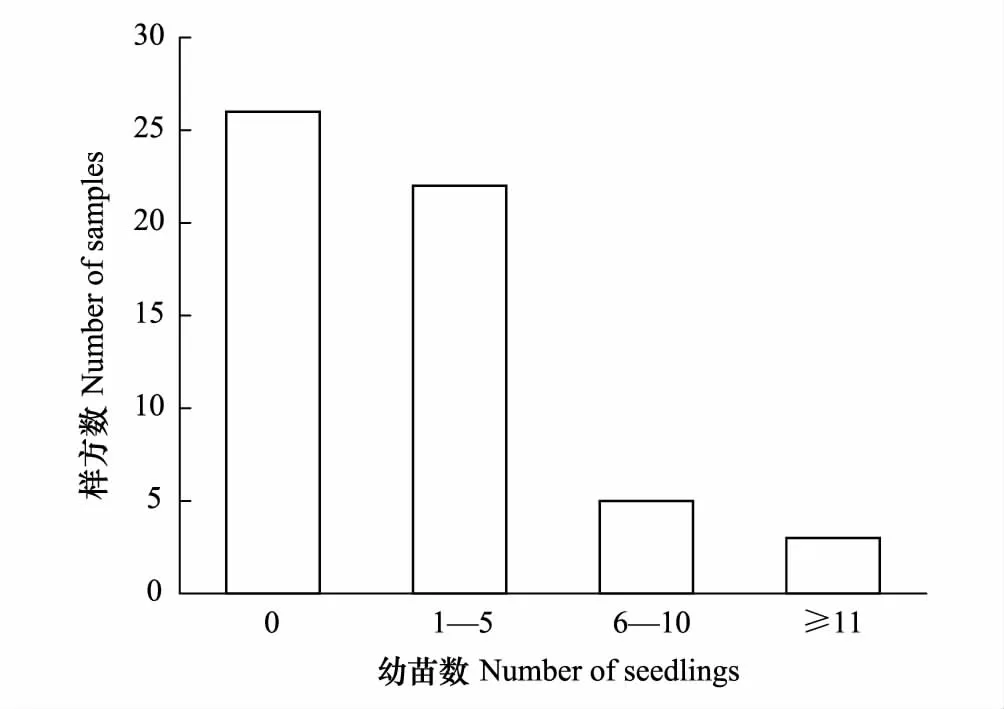
圖2 調查樣方內雅礱江冬麻豆幼苗數量分布Fig.2 Seedling distribution of S. bouffordiana in samples
雅礱江冬麻豆種群年齡結構呈紡錘形(圖1)。種群植株個體主要集中在第Ⅲ—XIV齡級,即地徑5—17 mm之間,占總植株數的80.71%。其中,地徑8—15 mm的植株數占總植株數的56.85%,說明第Ⅵ—Ⅻ齡級的植株數量充足,而地徑3—8 mm和>17 mm之間的植株數分別占總植株數的18.27%和14.72%,說明雅礱江冬麻豆種群缺乏低齡級和高齡級植株,且第Ⅰ—Ⅲ齡級的植株個體數占比之和不足9%,第XXIII—XXV級的植株個體數占比之和不足3%,表明種群有一定的更新能力,但更新后勁不足,甚為缺乏齡級兩端處的植株,種群呈現出衰退狀態。
根據調查樣方幼苗數統計顯示,約53.57%的樣方中有雅礱江冬麻豆幼苗,其中以1—5株幼苗的居多,占總樣方數的39.29%(圖2),表明研究區內幼苗雖有分布,但其個體數量仍相當缺乏,種群更新能力低下。雅礱江冬麻豆種群整體呈現出中齡數量豐富,低齡數量缺乏,高齡數量嚴重不足的紡錘形結構,種群更新不良,種群年齡結構為衰退型。
由表1可知,雅礱江冬麻豆種群V6、V17、V19、V23均等于0,說明第Ⅵ—Ⅶ、XVII—XVIII、XIX—XX、XXIII—XXIV齡級間的個體數量沒有變化,V4、V8、V10、V12、V14、V15、V16、V18、V20均大于0,說明第Ⅳ—Ⅴ、Ⅷ—Ⅸ、Ⅹ—Ⅺ、Ⅻ—XIII、XIV—XVII、XVIII—XIX、XX—XXI齡級間的個體數量呈現增長的結構動態關系,而剩余的齡級間均呈現出個體數量減少的結構動態關系,表明種群總體呈現出“衰退—增長—衰退—增長”的往復波動性結構動態關系。進一步分析其數量動態變化指數Vpi(不考慮外部干擾)和Vpi′(考慮隨機干擾)的結果顯示,Vpi=0.0799,Vpi′=0.0032,Vpi>Vpi′,雖然種群總體動態變化指數Vpi大于0,但僅為0.0799,且Vpi′值僅為0.0032,均趨向于0,表明種群波動大,種群穩定性差,抗干擾能力低。隨機干擾風險極大值P極大=0.04,即種群結構對隨機干擾的敏感性指數為0.04,說明雅礱江冬麻豆種群對外界隨機干擾具有較高的敏感性,表明其對環境的適應力低,而且由于種群結構低齡個體更新不良,因此雅礱江冬麻豆種群應屬于衰退型種群。
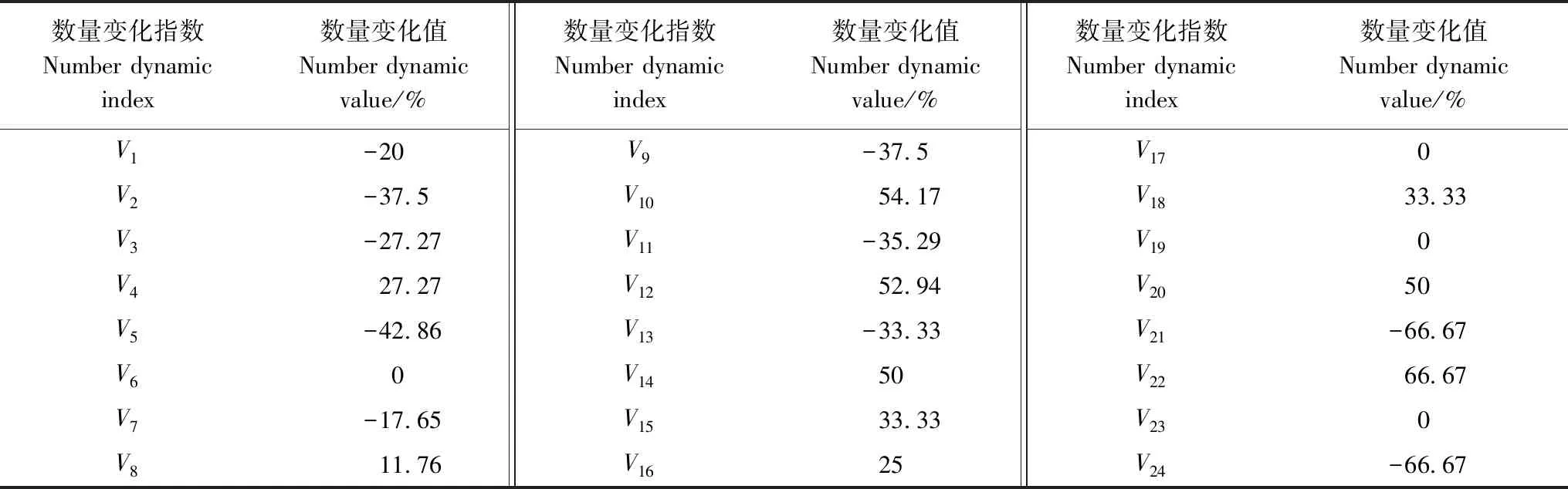
表1 雅礱江冬麻豆種群個體數量變化
Vn:種群從第n到n+1齡級的個體數量變化,V1為種群從第Ⅰ到Ⅱ齡級的個體數量變化,V2、……、V24以此類推 the individual number change of the population from age classnto age classn+1,V1means the individual number change of the population from age classⅠ to age class Ⅱ, theV2toV24are calculated in the same way
2.2 雅礱江冬麻豆種群靜態生命表
由雅礱江冬麻豆種群靜態生命表可知,隨著年齡的增大,雅礱江冬麻豆種群存活數(lx)逐漸減小(表2)。植株個體期望壽命(ex)隨著齡級的增加呈現出先增后減的狀態,其反應了植株個體的生命期望壽命,其最小期望壽命在第XXV齡級處(ex=1),最大期望壽命在第XIV齡級處(ex=7.667),說明雅礱江冬麻豆種群在較高齡級階段仍然具有較高的生命期望。種群死亡率(qx)與消失率(Kx)反映了種群數量隨著齡級增加的動態變化,雅礱江冬麻豆種群死亡率與消失率在第Ⅻ齡級前均隨齡級增加而呈增加的狀態,但在第Ⅻ齡級及其后出現劇烈波動,表明種群處于極不穩定狀態。因此,雅礱江冬麻豆種群整體呈現出極不穩定的狀態,種群發展狀況差。
經R3.3.3軟件擬合,雅礱江冬麻豆種群存活曲線的擬合方程為:
Nx=186.3488e-0.21878x(R2=0.8424,P<0.001)
Nx=1418.521x-2.10069(R2=0.998,P<0.001)
模型檢驗結果表明,兩種模型的擬合結果在雅礱江冬麻豆種群中均達到了極顯著水平,由于冪函數模型的R2值比指數函數模型大,故認為雅礱江冬麻豆種群存活曲線更趨近于Deevey-Ⅲ型。依據Deevey的劃分[22]:Ⅰ型呈凸曲線,多數個體都能活到該物種的生理年齡,但當達到一定生理年齡時則大量死亡;Ⅱ型呈直線,說明各年齡的死亡率相等;Ⅲ型是凹曲線,早期死亡率極高。因此雅礱江冬麻豆種群具有早期死亡率極高的特點。
以齡級為橫坐標,以存活數的對數為縱坐標,根據雅礱江冬麻豆種群靜態生命表繪制存活曲線(圖3)。由圖3可知,雅礱江冬麻豆種群存活曲線在第Ⅰ到第Ⅲ齡級幾乎成直線下降,進入第Ⅳ齡級后,逐漸趨緩,且變化率在第XV、XVI、XIX齡級處和第XXII齡級及其之后均為0,同時自第XXI齡級之后存活數的對數值也均為0,表明雅礱江冬麻豆種群低齡級死亡率高,中齡級植株存活率相對較高,當種群進入第XV、XVI、XIX齡級及第XXI齡級之后大部分植株會較快死亡。在存活曲線方面,雅礱江冬麻豆種群與瀕危物種鵝掌楸、矮牡丹、裂葉沙參、木根麥冬、長喙毛茛澤瀉種群的研究結果一致[23],均為Deevey-Ⅲ型,高死亡率集中在低齡階段,具有生殖能力的成年個體少。
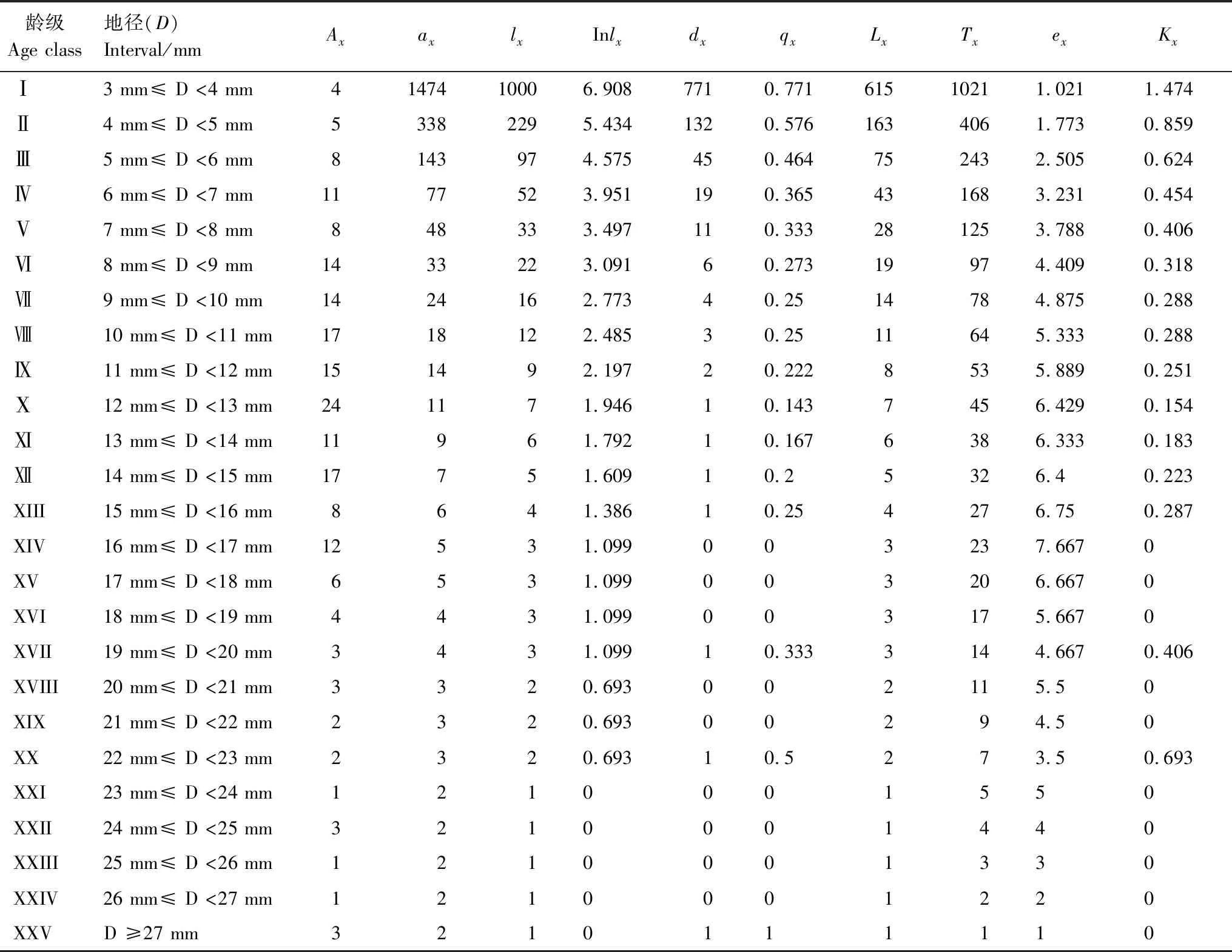
表2 雅礱江冬麻豆種群靜態生命表
Ax:x齡級內現有個體數 individual number of age classx;ax:Ax勻滑后x齡內的存活個體數 the revised data ofAx;lx:x齡級開始時標準化存活個體數 the standardized number of surviving individuals of age classx;Inlx:lx的自然對數 the natural logarithm oflx;dx:從x到x+1齡級間隔期內標準化死亡數 the standardized number of death individuals from age classxto age classx+1;qx:從x到x+1齡級間隔期間死亡率 mortality from age classxto age classx+1;Lx:從x到x+1齡級間隔期間還存活個體數 the number of surviving individuals from age classxto age classx+1;Tx:從x齡級到超過x齡級的個體總數 the total individuals number of age classxand age classes elder thanx;ex:進入x齡級個體的生命期望或平均期望壽命 life expectancy or average life expectancy of individuals in the age classx;Kx:消失率 disappearance rate
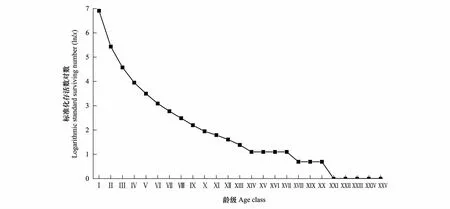
圖3 雅礱江冬麻豆種群的存活曲線Fig.3 Survival curve of S. bouffordiana population
2.3 雅礱江冬麻豆種群生存
由圖4可知,雅礱江冬麻豆種群的生存率隨齡級增大表現出先急劇減小而后趨緩的狀態,累計死亡率呈現出先快速遞增而后趨緩的態勢,二者互補,但種群生存率遠小于累計死亡率,表明種群處于衰退狀態。生存率曲線和累計死亡率曲線在前3個齡級呈現出急劇升降變化,在第Ⅴ齡級后逐漸趨緩,這可能和種群在低齡級的高死亡率有關,大量低齡級個體死亡,種群呈現衰退特征。在第Ⅵ齡級處,種群生存率為0.016,累計死亡率為0.984,種群已進入衰退期,表明雅礱江冬麻豆種群在生存早期已處于衰退狀態。
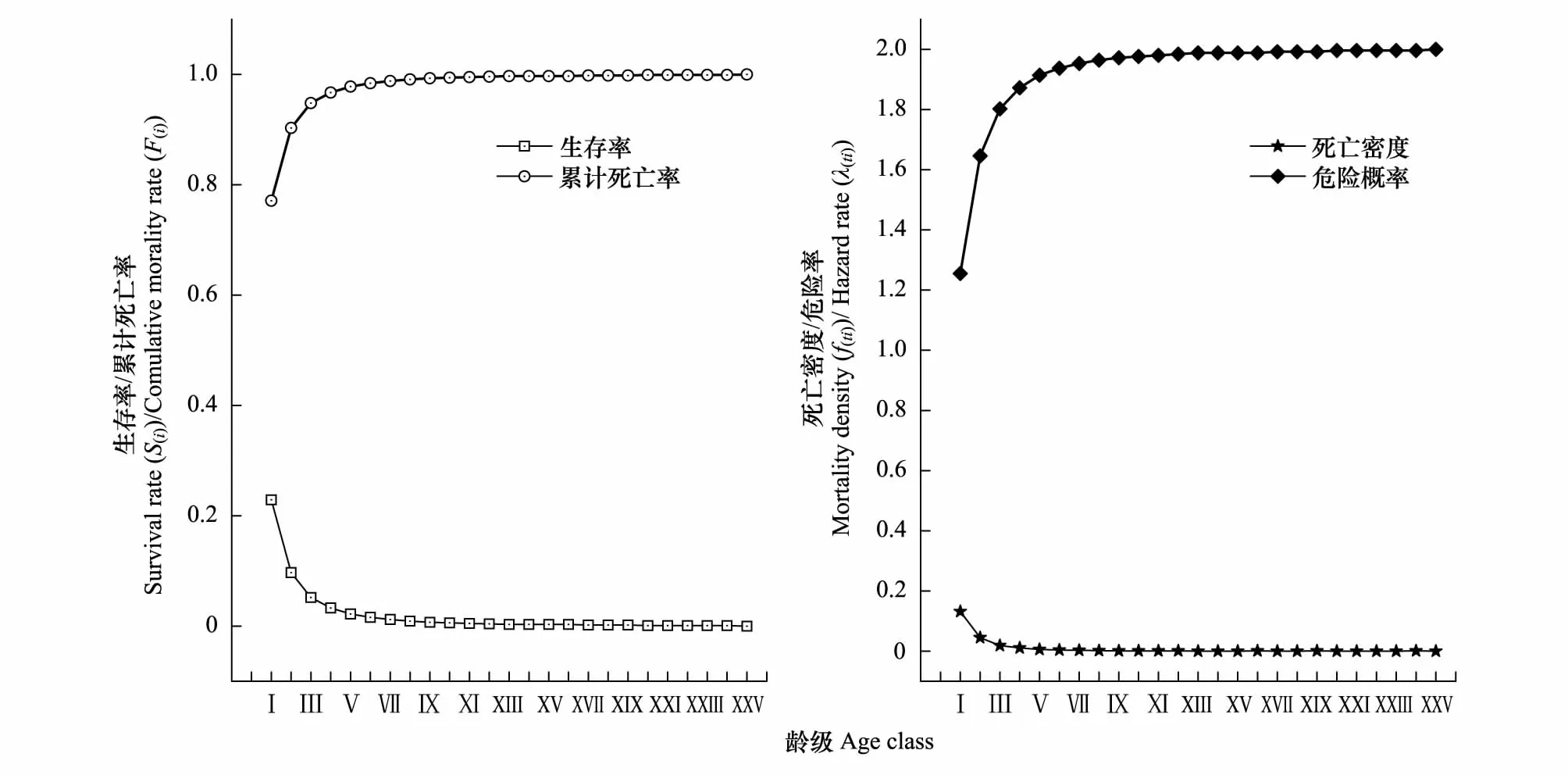
圖4 雅礱江冬麻豆種群生存率、累計死亡率、死亡密度和危險率函數曲線Fig.4 Survival rate, cumulative mortality rate, mortality density and hazard rate functional curves of S. bouffordiana population
種群死亡密度曲線和危險概率曲線與其生存率和累計死亡率變化趨勢相似,均在前3個齡級急劇變化,但種群危險概率曲線一直顯著大于死亡密度曲線,在進入第Ⅳ齡級后均變化趨緩,其中危險概率曲線的整體變化更顯著,死亡密度曲線在第XXV齡級達到最小值(0),危險概率曲線在第XXV齡級達到最大值(2)。由4 個生存函數曲線表明,雅礱江冬麻豆種群具有前期薄弱,中期、后期衰退顯著的特點,種群生存力較低,處于衰退狀態。
2.4 雅礱江冬麻豆種群數量的時間序列
雅礱江冬麻豆種群數量動態變化的時間序列預測是以其種群各齡級株數為原始數據,按照一次移動平均法預測出各齡級在未來2、4、9、6、8、12、20和24個齡級時間后的個體數。從表3分析可知,經歷2、4、6個齡級時間,種群從第Ⅻ齡級開始之后均有不同程度的增加,經過8和12個齡級時間,從第Ⅻ齡級開始減少,而后趨于穩定,說明一旦過了一定的生理年齡,雅礱江冬麻豆種群就開始逐漸走向衰退。種群前期,在第Ⅵ齡級經過2、4和6個齡級時間后個體數分別下降了21.43%、28.57%和42.86%;種群前中期,在第Ⅺ齡級經過2、4、6和8個齡級時間后個體數下降了16.67%、25.00%、37.50%和41.67%;而種群中期,在第Ⅻ齡級經過12個齡級時間后個體數下降了29.41%。表明雅礱江冬麻豆種群嚴重缺乏低齡個體。綜合其整個發展趨勢來看,雅礱江冬麻豆種群正常更新難以維持,未來仍然會繼續衰退。
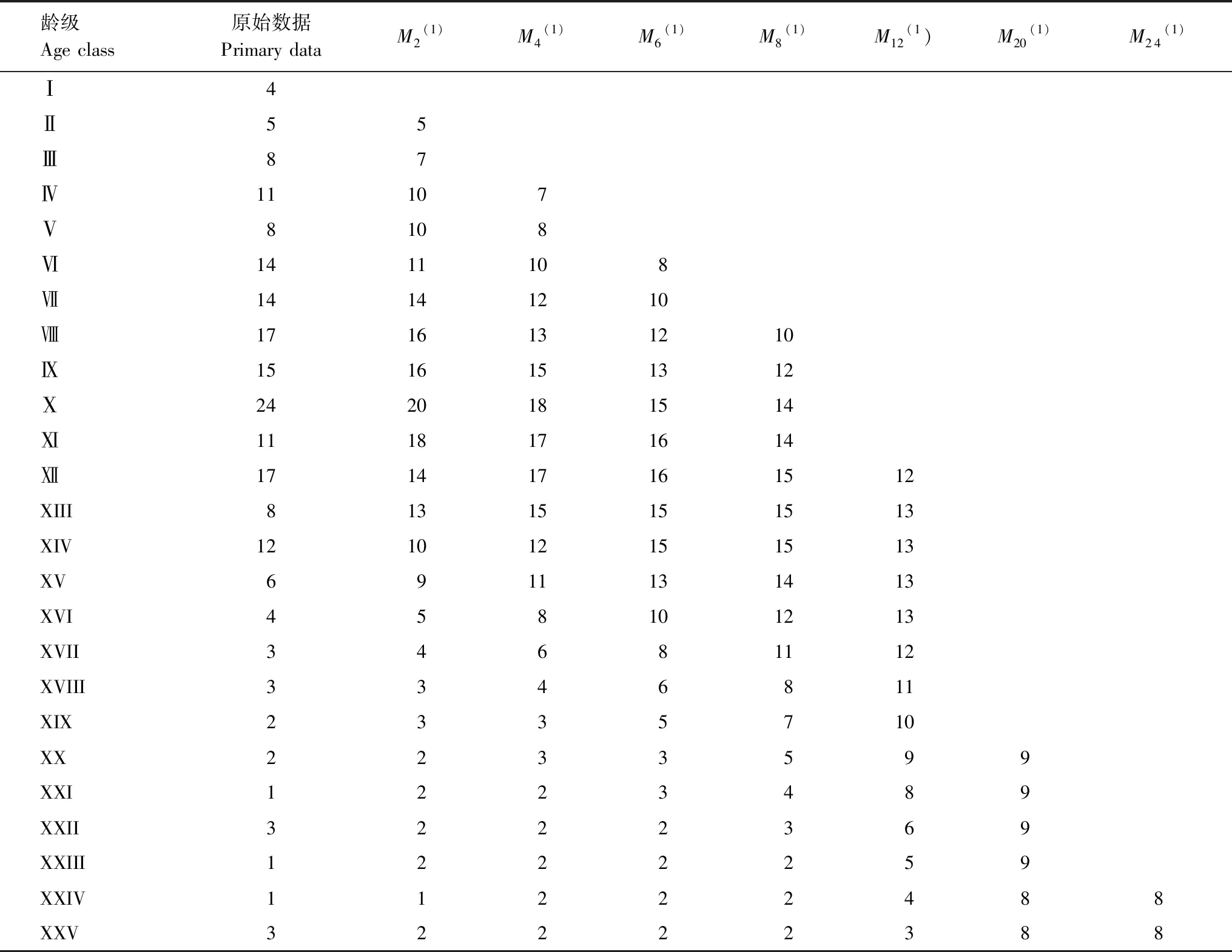
表3 雅礱江冬麻豆種群動態變化的時間序列分析
Mt(1):經過2、4、6、8、12、20和24齡級時間后t齡級的種群大小the population size of age class t after the time of 2, 4, 6, 8, 12, 20, 24 age class;(1):移動平均法中的一次移動平均法標識the mark of one-time moving average in moving average method
3 討論
植物種群結構特征能夠體現種群內部個體的發展過程,其徑級結構能很好地反應種群動態變化[24]。生命表是判斷植物種群發展趨勢的重要指標之一,對植物種群的生命表和存活曲線的分析可以反映種群現實狀況、展現植物種群與環境的競爭關系,對珍稀瀕危植物的保護和利用具有重要意義[25]。本研究得出雅礱江冬麻豆種群年齡結構為紡錘形衰退型,動態指數顯示種群波動大、穩定性差、抗干擾能力低,且其對生存環境適應力低、具有較高的敏感性。靜態生命表顯示雅礱江冬麻豆種群整體呈現出極不穩定的狀態,種群發展狀況差。其存活曲線為Deevey-Ⅲ型,早期死亡率極高。生存函數曲線表明雅礱江冬麻豆種群具有前期薄弱,中期、后期衰退顯著的特點,種群生存力較低,處于衰退狀態。時間序列分析顯示雅礱江冬麻豆種群嚴重缺乏低齡個體,種群正常更新難以維持,未來仍然會繼續衰退。因此,雅礱江冬麻豆種群為衰退種群,急需進一步弄清雅礱江冬麻豆種群的衰退原因和衰退程度,也需對雅礱江冬麻豆種群進行生物化學與分子生物學、生態學和地理學等方面的綜合研究。
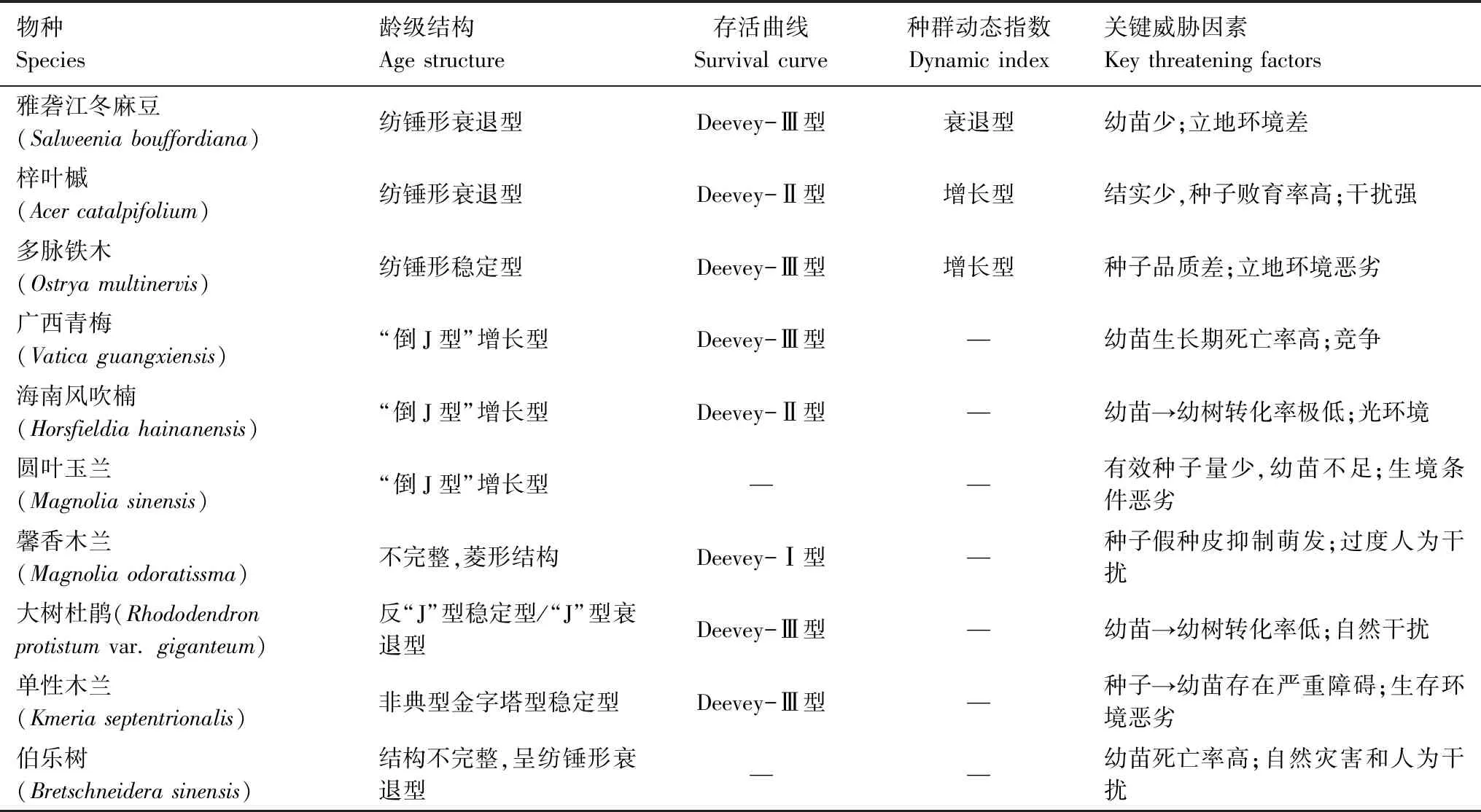
通過對極小種群梓葉槭(Acercatalpifolium)[26]、多脈鐵木(Ostryamultinervis)[27]、廣西青梅(Vaticaguangxiensis)[28]、海南風吹楠(Horsfieldiahainanensis)[29]、圓葉玉蘭(Magnoliasinensis)[30]、馨香木蘭(Magnoliaodoratissma)[31]、大樹杜鵑(Rhododendronprotistumvar.giganteum)[32]、單性木蘭(Kmeriaseptentrionalis)[33]、伯樂樹(Bretschneiderasinensis)[34]和雅礱江冬麻豆種群的種群結構與動態特征比較發現(表4):(1)從齡級結構上看,梓葉槭、大樹杜鵑、伯樂樹與雅礱江冬麻豆種群均呈現出衰退型;(2)從存活曲線上看,多脈鐵木、廣西青梅、大樹杜鵑、單性木蘭與雅礱江冬麻豆種群均為Deevey-Ⅲ型,而梓葉槭、海南風吹楠種群為Deevey-Ⅱ型(除馨香木蘭種群為Deevey-Ⅰ型外),這與張文輝等對十種瀕危植物種群總結出來的存活曲線結論基本一致[23];(3)從種群動態指數上看,梓葉槭和多脈鐵木為增長型,而雅礱江冬麻豆種群均為衰退型;(4)對種群關鍵威脅因素進行分析,表明種群均與種子問題或者幼苗問題有關,而外因與干擾、立地環境和競爭有關。所以,這些極小種群和雅礱江冬麻豆種群均表現出脆弱性高的共性。
表4 極小種群和雅礱江冬麻豆的種群結構與動態特征比較分析
Table 4 The comparison in population structures and dynamic characteristics between extremely small populations andS.bouffordianapopulation
物種Species齡級結構Age structure存活曲線Survival curve種群動態指數Dynamic index關鍵威脅因素Key threatening factors雅礱江冬麻豆(Salweenia bouffordiana)紡錘形衰退型Deevey-Ⅲ型衰退型幼苗少;立地環境差梓葉槭(Acer catalpifolium)紡錘形衰退型Deevey-Ⅱ型增長型結實少,種子敗育率高;干擾強多脈鐵木(Ostrya multinervis)紡錘形穩定型Deevey-Ⅲ型增長型種子品質差;立地環境惡劣廣西青梅(Vatica guangxiensis)“倒J型”增長型Deevey-Ⅲ型—幼苗生長期死亡率高;競爭海南風吹楠(Horsfieldia hainanensis)“倒J型”增長型Deevey-Ⅱ型—幼苗→幼樹轉化率極低;光環境圓葉玉蘭(Magnolia sinensis)“倒J型”增長型——有效種子量少,幼苗不足;生境條件惡劣馨香木蘭(Magnolia odoratissma)不完整,菱形結構Deevey-Ⅰ型—種子假種皮抑制萌發;過度人為干擾大樹杜鵑(Rhododendron protistum var. giganteum)反“J”型穩定型/“J”型衰退型Deevey-Ⅲ型—幼苗→幼樹轉化率低;自然干擾單性木蘭(Kmeria septentrionalis)非典型金字塔型穩定型Deevey-Ⅲ型—種子→幼苗存在嚴重障礙;生存環境惡劣伯樂樹(Bretschneidera sinensis)結構不完整,呈紡錘形衰退型——幼苗死亡率高;自然災害和人為干擾
根據種群結構與動態特征比較分析表明,雅礱江冬麻豆種群與極小種群存在多方面的相似,但其生物學地位卻存在差距。確定瀕危植物種群的生物學地位是植物保護工作的第一步[35],而雅礱江冬麻豆種群的生物學地位還未引起足夠的重視。任何特有種都可能受到其有限分布的威脅[36],種群滅絕的概率與其種群大小呈負相關[35]。雅礱江冬麻豆屬于小種群,為衰退型結構,種群更新后勁乏力,生存力較低,且已處于瀕危狀態,面臨滅絕的可能。研究發現,人類活動是造成物種瀕危的首要因素[35],而人類活動無處不在,隨著人類活動的不斷擴張,雅礱江冬麻豆種群的生存將面臨著更為嚴峻的形勢。因此,需要相關部門加快對雅礱江冬麻豆的研究步伐和盡快確定其生物學地位并對其進行定級,確保其得到有效保護。
就地保護是瀕危植物解危的主要措施[23],其能使保護植物避免諸多外界干擾。本文建議對雅礱江冬麻豆種群進行就地保護,在雅礱江冬麻豆集中分布的區域設立野外保護小區,使其擁有一個修養生息的生存空間,每個保護小區必須針對所保護的物種特點制定相應的保護管理辦法,加強保護點原生境的保護管理力度,加強巡護、監管,嚴禁盜伐、開荒、放牧、火燒、種植等改變原生境生態的行為,嚴格保護原生生境,防止人為干擾、自然災害、獸類侵害的發生,保障原生種群的穩定和逐步繁衍壯大。
雅礱江冬麻豆種群年齡結構呈紡錘形衰退型結構、存活曲線為Deevey-Ⅲ型、動態指數顯示為衰退型、對外界干擾具有較高的敏感性,屬于亟需進行生物學定位的瀕危物種。使物種出現敏感性的不穩定環境因素是造成物種陷入危機的主要原因,甚至可能會引起瀕危物種的滅絕,所以保護雅礱江冬麻豆幼苗定居和存活以及為其營造穩定的生存環境是維持雅礱江冬麻豆種群存續和穩定的重要措施。
