鈍葉柃花器官性別分化的形態學研究*
2020-03-19 07:23:48鄧洪平倪東萍程蒞登巴羅菊
林業科學 2020年2期
顧 梨 何 平,2 鄧洪平,2 倪東萍 王 茜 程蒞登 巴羅菊
(1.西南大學生命科學學院 三峽庫區生態環境教育部重點實驗室 重慶 400715;2.重慶市科學技術研究院 重慶 400715)
鈍葉柃(Euryaobtusifolia)為山茶科(Theaceae)柃木屬(Eurya)常綠灌木或小喬木,我國主要分布于陜西、湖北、湖南、貴州、四川及云南等省的海拔400~1 450 m的山地疏林、密林以及林緣路旁灌叢中。柃木屬植物是熱帶和亞熱帶常綠闊葉林灌木層優勢種之一(馬德風等,1993),其物種的穩定對維持森林生態系統有重要的作用。傳統認為柃木屬植物是嚴格的雌雄異株,但在重慶縉云山發現鈍葉柃中存在雌株、雄株以及Leaky個體等類型,Leaky個體指典型的兩性變異株在性別表達上偏離于正常雌株及雄株:有的具有兩性功能;有的僅具雌性功能,但具不同程度退化的雄蕊;有的僅具雄性功能,但不育雌蕊發育明顯(王茜等,2012;李運婷,2016;李運婷等,2016)。這一現象在自然種群中發生的頻率不高,隨著野外工作的深入,相關的報道越來越多,柿科(Ebenaceae)(Venkatasamyetal.,2007)、茄科(Solanaceae)(Martineetal.,2014;Andersonetal.,2015)等10余科中均發現了這一現象,但目前對出現這一現象的原因尚不清楚。
植物性別分化是雌蕊和雄蕊的發生及分化的過程,具體包括雌、雄性別的決定和配子體的分化、發育與成熟等階段。對于雌雄異株和雌雄同株植物來說,性別分化是原始兩性花原基的雄性或雌性選擇性敗育及相應的雄蕊或雌蕊中配子體發生分化的結果(張大勇,2004)。目前對鈍葉柃出現Leaky現象的研究僅在生理生化、傳粉、群落特征等方面(王茜等,2012;李運婷,2016;李運婷等,2016),對其花芽分化及花器官發育過程的形態學研究仍然缺乏。因此,本文采用常規石蠟切片法對鈍葉柃3種典型雌株、雄株、兩性變異株不同分化時期的花芽形態和結構進行觀察,探討3種不同性別花器官發育過程的異同,明確性別分化的關鍵時期,為探討性別分化的相關機制及植物類群之間的系統進化關系提供重要的形態學證據。
1 材料與方法
1.1 試驗材料 試驗材料采自重慶縉云山國家級自然保護區金果園(106°17′—106°24′E,29°41′—29°52′N),該區屬中亞熱帶季風濕潤性氣候,平均氣溫3.1 ℃。區域內以常綠闊葉林、針闊混交林等植被類型為主,灌木層主要優勢種為柃木屬植物。2016年2月,盛花期時選取長勢健壯且一致的鈍葉柃典型雌株、雄株、兩性變異株各12株,平均樹高2.5 m,胸圍6.4 cm,冠幅3.4 m2,進行掛牌標識。
1.2 試驗方法 2016年7月—2018年3月,隨機選取典型雌株、雄株、兩性變異株各12株,每株當年生新枝上形成的頂梢下第3-5芽位,采集結構完整、無病蟲害的花芽共30個,每3天1次。選擇其中的8~10個形態大小一致的花芽,用MODEL C-BD230解剖鏡測量花芽的長度、寬度,同時拍照記錄外部形態特征;剩余花芽根據其大小分別采用FAA固定液(甲醛∶冰醋酸∶70%酒精=5∶5∶90,體積比)固定,存于4 ℃冰箱。
固定24 h,取出固定后的花芽,針對鈍葉柃花芽包裹緊實、難以浸蠟的特點,對常規石蠟切片法(李桂琴,2002)加以改進:材料脫水之前用質量分數20%的鹽酸軟化24 h,流水沖洗1~2天后制作石蠟切片,切片厚度8~10 μm;采用蘇木精-鐵礬、番紅-固綠對染,加拿大中性樹膠封片,制成永久切片。用NIKON ECLIPSE 型顯微鏡觀察花芽縱切面及橫切面的解剖結構并拍攝照片,記錄花芽分化過程中解剖結構的變化特征。到花芽開放時,5%~25%花朵開放確定為始花期;25%~75%花朵開放確定為盛花期;75%以上的花朵開放確定為末花期。
記錄的數據采用Microsoft Office Excel 2016和SPSS 20.0進行統計分析。
2 結果與分析
2.1 鈍葉柃成花物候期及花芽解剖結構特征1) 成花物候期 對鈍葉柃成花物候進行觀測(圖1),7月底至8月上旬,3種不同性別鈍葉柃植株的當年生新枝及2年生枝葉腋內出現1~4個芽;8月下旬,苞片原基開始分化與發育,花原基出現,進入花芽分化階段;8月底至11月上旬,花芽的各輪花器官相繼分化、發育;12月中下旬至1月底,氣溫急劇下降,花芽進入休眠時期,此時雌雄蕊已分化發育完全;次年2月初,氣溫持續回升10~15天,進入始花期。雄株的始花期早于雌株花期3~4天,兩性變異株始花期晚于雌花11~14天,盛花期一般在始花期后的5~12天出現,維持7~10天,步入終花期,終花期維持5天左右。以花器官原基出現的時間以及花芽外部形態特征為劃分標準(王彩云等,2002),將花芽分化期分為苞片分化期(stage of bract differentiation,SBD)、萼片分化期(stage of sepal differentiation,SSD)、花瓣分化期(stage of petal differentiation,SPD)、雌雄蕊分化期(stage of stamen and pistil differentiation,SSPD)、雌雄蕊成熟期(stage of stamen and pistil formation,SSPF)5個時期。
鈍葉柃3種不同性別花芽在分化初期形態變化及內部結構保持一致,僅發育的時間有所差異,雌花芽發育滯后雄花芽8~10天,兩性變異花芽滯后雌花芽15~20天。
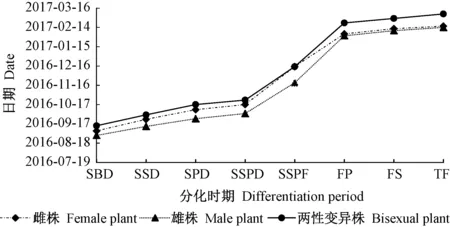
圖1 不同性別鈍葉柃開花物候期Fig.1 Blossoming phenophase among different sexes of E.obtusifoliaSBD:苞片分化期;SSD:萼片分化期;SPD:花瓣分化期;SSPD:雌雄蕊分化期;SSPF:雌雄蕊成熟期;FP:始花期;FS:盛花期;TF:終花期。SBD:Stage of bract differentiation;SSD:Stage of sepal differentiation;SPD:Stage of petal differentiation;SSPD:Stage of stamen and pistil differentiation;SSPF:Stage of stamen and pistil formation;FP:Flowering period;FS:Full-blooming stage;TF:The final flowering.
2) 苞片分化期(SBD) 8月中旬至9月初左右,葉腋2枚小苞片原基包裹著芽尖,此時植株由營養生長轉為生殖生長,小苞片原基逐漸發育呈片狀結構,同時在中間位置分化出半球形的凸起(花原基)(圖2A、B、C)。
3) 萼片分化期(SSD) 9月初到9月中下旬,進入萼片原基分化期,2枚小苞片中間的花原基分化形成萼片原基,外層細胞分裂速度較快,萼片原基呈“鉤”狀,接著中間突起生長點細胞繼續側向分裂,伸長并向內彎曲,最終形成片狀弧形,在縱切面上僅見2個萼片原基(圖2D、E、F)。
4) 花瓣分化期(SPD) 9月中旬到10月初,在萼片原基分化的同時,中央的生長錐細胞繼續分裂形成花瓣原基,花芽進入花瓣分化期,15~17天。花瓣原基與萼片原基分裂的方式相似,生長錐細胞先進行橫向分裂,中央位置微凹,后進行側向分裂,伸長并向內彎曲,呈片狀弧形(圖2G、H、I)。
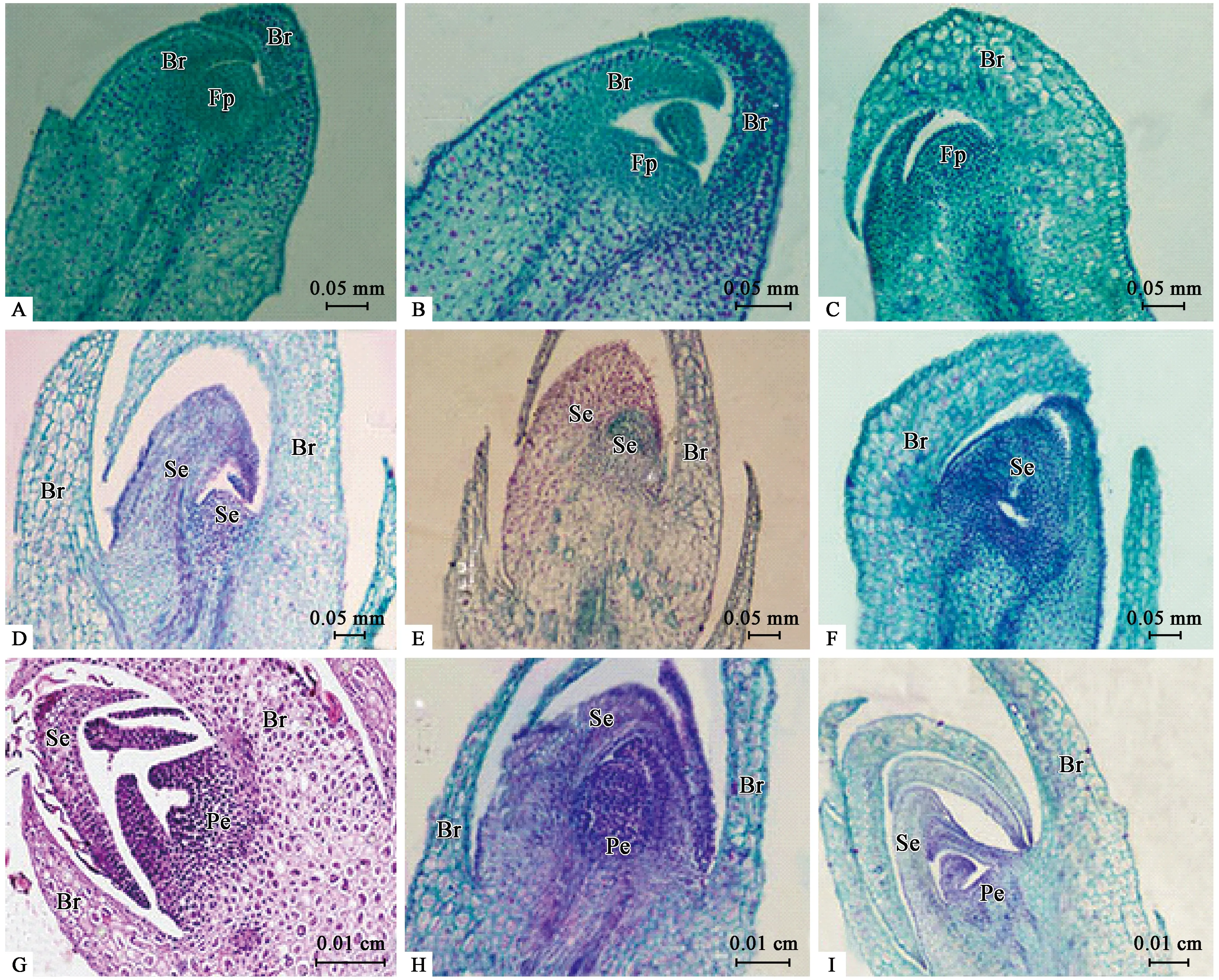
圖2 鈍葉柃花芽分化中苞片、萼片及花瓣分化時期石蠟切片Fig.2 Section observation of the development of bracts,sepals and petal during the flowering stages of E.obtusifolia.A,B,C:分別為雌花芽、雄花芽和兩性變異花芽的苞片分化期;D,E,F:分別為雌花芽、雄花芽和兩性變異花芽的萼片分化期;G,H,I:分別為雌花芽、雄花芽和兩性變異花芽的花瓣分化期。Fp:花原基;Br:苞片;Se:萼片原基及萼片;Pe:花瓣原基。A,B,C:Stage of bract differentiation in female,male and bisexual flower buds,respectively;D,E,F:Stage of Sepal differentiation in female,male and bisexual flower buds,respectively;G,H,I:Stage of petal differentiation in female,male and bisexual flower buds,respectively.Fp:Flower primordium;Br:Bracts;Se:Sepal primordia and sepals;Pe:Petal primordia.
5) 雌雄蕊分化期(SSPD) 雄花芽的雌雄蕊分化:9月底至10月初,萼片和花瓣原基分化的后期,花瓣內側的生長錐持續分裂,中央位置出現雌蕊原基,兩側形成指狀的雄蕊原基(圖3B)。雄蕊原基和雌蕊原基同時分化,后期分化過程中,雌蕊原基停止分化形成半球形突起(圖3E、H)。
雄花芽的花絲花藥分化:10月初至10月上旬,萼片和花瓣繼續伸長分裂,相互疊壓,隨后進入花藥、花絲的分化,指狀雄蕊原基上部膨大,下部纖細且短,呈“棒槌”狀。膨大部分側向生長旺盛,分化為花藥,基部進一步伸長,分化為花絲(圖3H、圖4B)。
雌花芽的雌蕊分化:10月初至10月中下旬左右,花瓣原基分化末期,花芽中央染色較深部分為雌蕊原基,不產生雄蕊原基(圖3A)。雌蕊原基不斷分裂,切片可見產生2個心皮原基,心皮原基持續分裂,基部愈合形成大的突起(圖3D、G)。
兩性變異花芽的雌雄蕊分化:10月中旬至10月底,兩性變異花芽開始進入雌雄蕊的分化期,與雄花芽的分化初期相似,同時出現雄蕊原基和雌蕊原基(圖3C、F),雌雄蕊原基正常發育,雄蕊原基發育的速度較雌蕊原基慢,心皮原基遠大于雄蕊原基(圖3I)。
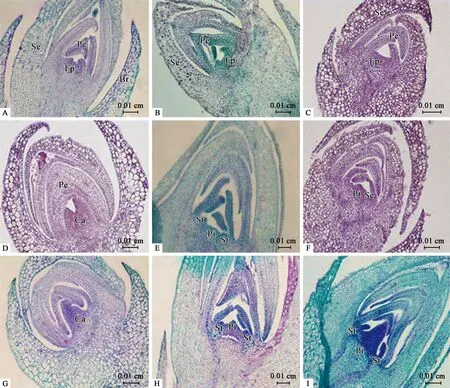
圖3 鈍葉柃花芽分化中雌雄蕊分化期石蠟切片Fig.3 Section observation of the stage of stamen and pistil differentiation during the flowering stages of E.obtusifoliaA:雌花芽中雌蕊原基的分化初期;B:雄花芽中雌雄蕊原基的分化初期;C:兩性變異花芽中雌雄蕊原基的分化初期;D:雌花芽中心皮原基的分化;E:雄花芽中雌蕊、雄蕊原基同時分化;F:兩性變異花芽中雌蕊、雄蕊原基的分化;G:雌花芽心皮原基愈合;H:雄花芽中心皮原基、花藥、花絲的分化;I:兩性變異花芽中雌、雄蕊原基的發育。Br:苞片;Se:萼片原基及萼片;Ep:花原基;Pe:花瓣原基;Pi:雌蕊原基;St:雄蕊原基;Ca:心皮原基。A:Initial differentiation of the pistil primordium in female flower buds;B:Initial differentiation of the primordium of pistil and stamen in male flower buds;C:Initial differentiation of the primordium of pistil and stamen in bisexual flower buds;D:Differentiation of the carpel primordium in female flower buds;E:Pistil and stamen primordia differentiate simultaneously in male flower buds;F:Differentiation of pistil and stamen primordia in bisexual flower buds;G:Basal union of carpel of female flower buds;H:Differentiation of carpels,anthers and filaments in male flower buds;I:Development of the primordium of pistil and stamen in bisexual flower buds.Br:Bracts;Se:Sepal primordia and sepals;Ep:Flower primordium;Pe:Petal primordia;Pi:Pistil primordium;St:Stamen primordium;Ca:Carpel primordium.
6) 雌雄蕊成熟期(SSPF) 10月中下旬至12月下旬,雌花芽中,心皮原基基部膨大,形成子房原基的小室,中部縱向凹陷形成空腔(圖4A、D),頂部愈合向上分化成花柱;隨后,子房基部內側分化形成中軸胎座,胎座兩側分化出胚珠原基(圖4E),最終形成倒生胚珠;花柱繼續縱向伸長,頂端形成柱頭,子房逐漸膨大呈囊狀,胚珠逐漸成熟(圖4F)。雄花芽中,花絲繼續伸長,花藥形成花粉囊(圖4G、H)。在兩性變異花芽中,雌雄蕊皆發育完全(圖4C),也有少數花芽中雄蕊或雄蕊原基發育不正常(圖4I)。
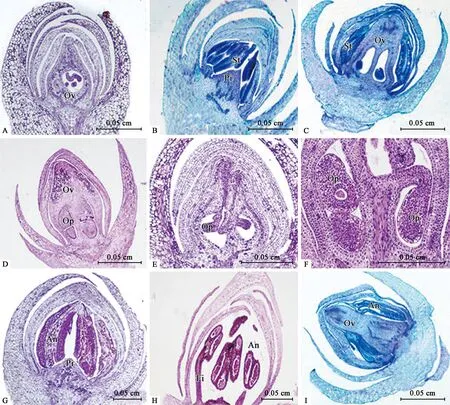
圖4 鈍葉柃花芽分化中雌雄蕊成熟期石蠟切片Fig.4 Section observation of the stage of stamen and pistil formation during the flowering stages of E.obtusifoliaA:雌花芽中子房的形成;B:雄花芽中雄蕊原基的分化后期;C:兩性變異花芽中雌、雄蕊的發育;D,E:雌花芽中子房的發育;F:雌花芽中胚珠的發育;G,H:雄花芽花藥、花絲發育;I:兩性變異花芽中退化雌蕊。Pi:雌蕊原基;St:雄蕊原基;Ov:子房;Op:胚珠原基及胚珠;An:花藥;Fi:花絲。A:Ovary formation in female flower buds;B:Anaphase of stamen primordium in male flower buds;C:Development of pistil and stamen in bisexual flower buds;D,E:Ovary development in female flower buds;F:Ovule development in female flower buds;G,H:The anther and filament development in male flower buds;I:Pistil degradation in bisexual flower buds.Pi:Pistil primordium;St:Stamen primordium;Ov:Ovary;Op:Ovules and ovule primordia;An:Anthers;Fi:Filaments.
2.2 花芽分化外觀形態的變化特征 如圖5所示,鈍葉柃3種不同性別花芽在分化過程中,雌花芽、兩性變異花芽的長寬比均呈先上升后下降的趨勢,在雌雄蕊分化期(SSPD)分別達到最高1.27、1.36,此時花器官處于分化的最高峰,后期由于子房、花藥不斷膨大,長寬比有所下降。雄花芽在萼片分化期(SSD)前,花芽縱向生長較橫向生長迅速,而隨著雄蕊原基的不斷發育,橫向生長速度加快,長寬比逐漸下降,到雌雄蕊成熟期最低(1.09)。從整個分化時期看,兩性變異花芽與雌花芽的長寬比動態變化相似,但兩性變異花芽的長寬比始終高于雌花芽。
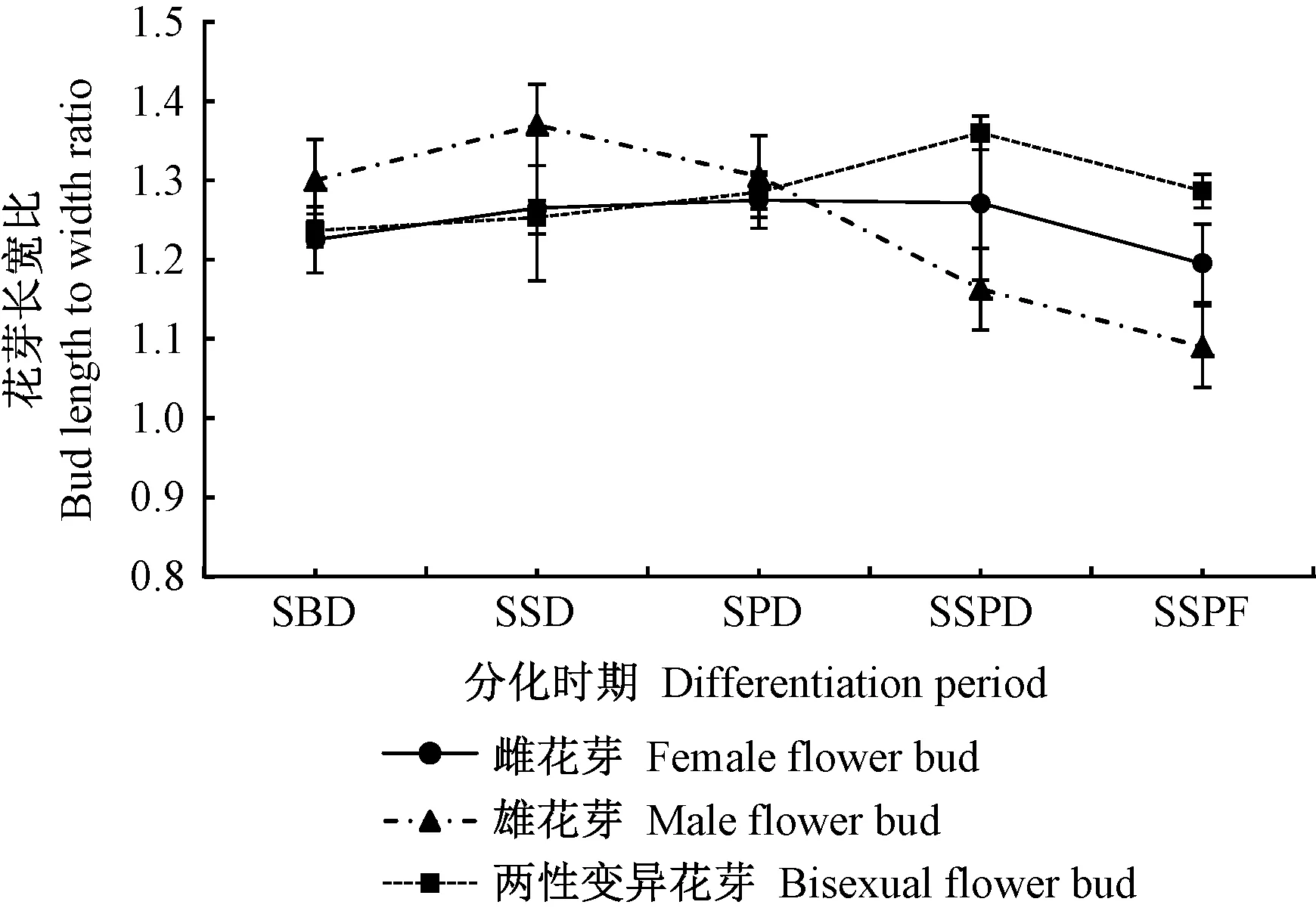
圖5 鈍葉柃花芽長寬比動態變化Fig.5 The flower bud length to width ratio of E.obtusifolia
鈍葉柃花芽在分化過程中外部形態特征隨著發育時期的不同會發生相應的改變,不同性別花芽之間也存在差異。在苞片分化期,下部寬胖頂端尖,呈三角形,扁平,綠色,表面有透明的柔毛,長0.645~0.745 mm、高0.470~0.572 mm(圖6A、E、I),長寬比1.22~1.30;萼片分化期,體積膨大,呈橢圓形或近圓形,綠色褪盡,呈深紫紅色,表面粗糙,白色柔毛增長,花芽長1.044~1.467 mm、寬0.874~1.035 mm(圖6B、F、J);花瓣分化期,花芽圓胖,明顯伸長、變寬,2枚小苞片緊包,呈綠色三角形,頂端深褐色,皆被透明長柔毛,長1.160~1.565 mm、寬0.891~1.125 mm(圖6C、G、K)。3種不同性別花芽在前3個時期的外部形態和色澤方面的變化一致,但體積方面雄花芽>兩性變異花芽>雌花芽。
雌雄蕊分化期,雄花芽頂端圓鈍,迅速增大增長,表面綠色加深,紫紅色逐漸褪去,長1.889~2.429 mm、寬1.671~2.106 mm,長寬比1.16(圖6H,圖7E)。雌花芽頂端漸尖,相對雄花芽較小,橢圓形,紫紅色逐漸褪去,綠色加深,被白色長毛,花芽長1.577~1.645 mm、寬1.035~1.241 mm(圖6D,圖7 A)。兩性變異花芽,與雌花芽相似(圖6L,圖7I)。
雌雄蕊成熟期,雄花芽變得更加圓潤,長寬比降至1.09,雌花芽和兩性變異花芽繼續增長加寬,萼片、花瓣也逐漸發育完全,覆瓦狀排列,花藥由白色變為淺黃色,子房綠色逐漸加深。此時花芽外部紫紅色完全褪盡,由嫩綠色逐漸變成黃綠色或棕綠色,2枚小苞片頂端出現深褐色,雌花芽、兩性變異花芽5枚萼片大小相近,深紫紅色,而雄花芽呈現綠色(圖7B、F、J);5枚花瓣,白色,花藥由無色透明轉變成深黃色,子房翠綠色,花柱微帶橘色(圖7C、G、K)。
1月底至2月初,進入始花期,雌花芽花瓣反卷,柱頭三裂,子房、花柱綠色透明(圖7D);雄花雄蕊9~11枚,呈“燈籠”型(圖7H);兩性變異花芽,雄蕊群不定數,雌蕊中胚珠不定數(圖7K、L)。

圖6 鈍葉柃花芽分化早期階段的形態變化Fig.6 Morphological changes at early stage of flower bud differentiation of E.obtusifoliaA,B,C,D:分別為雌花芽的苞片分化期、萼片分化期、花瓣分化期、雌蕊分化初期;E,F,G,H.分別為雄花芽的苞片分化期、萼片分化期、花瓣分化期、雌雄蕊分化初期;I,J,K,L:分別為兩性變異花芽的苞片分化期、萼片分化期、花瓣分化期、雌雄蕊分化初期。A,B,C,D:Stage of bract differentiation,sepal differentiation,petal differentiation and initial pistil differentiation in female flower buds,respectively;E,F,G,H:Stage of bract differentiation,sepal differentiation,petal differentiation and initial pistil and stamen differentiation in male flower buds,respectively;I,J,K,L:Stage of bract differentiation,sepal differentiation,petal differentiation and initial pistil and stamen differentiation in bisexual flower buds,respectively.

圖7 鈍葉柃花芽分化后期階段的形態變化Fig.7 Morphological changes at late stage of flower bud differentiation of E.obtusifolia A:雌花芽雌蕊分化后期;B:3種不同性別花芽雌雄蕊成熟初期(由左至右:兩性變異花芽、雌花芽、雄花芽);C:圖J中雌花芽解剖圖;D:雌花芽始花期;E:雄花芽雌雄蕊分化后期;F:3種不同性別花芽的雌雄蕊成熟中期(由左至右:兩性變異花芽、雌花芽、雄花芽);G:圖J中雄花芽解剖圖;H:雄花芽始花期;I:兩性變異花芽雌雄蕊分化后期;J:3種不同性別花芽雌雄蕊成熟后期(由左至右:兩性變異花芽、雌花芽、雄花芽);K:圖J中兩性變異花芽解剖圖;L:兩性變異花芽始花期。A:Late pistil differentiation in female flower buds;B:Early maturation of pistil and stamen in flower buds(left-right:bisexual,female,male flower buds);C:Anatomy of female flower buds in Fig.J;D:Female flower buds in early blooming;E:Late pistil and stamen differentiation in male flower buds;F:Middle maturation of pistil and stamen in flower buds(left-right:bisexual,female,male flower buds);G:Anatomy of male flower buds in Fig.J;H:Male flower buds in early blooming;I:Late pistil and stamen differentiation in bisexual flower buds;J:Late maturation of pistil and stamen in flower buds(left-right:bisexual,female,male flower buds);K:Anatomy of bisexual flower buds in Fig.J;L:Bisexual flower buds in early blooming.
3 討論
花芽分化是復雜的形態建成和生理生化過程,是有花植物發育的關鍵時期,也是植物體內外多重因子共同作用、相互協調的結果(郜愛玲等,2010;張波等,2017)。鈍葉柃花芽分化劃分為苞片分化期、萼片分化期、花瓣分化期、雌雄蕊分化期、雌雄蕊成熟期。不同研究者對同科同屬近緣種花芽分化時期的劃分不盡相同,此劃分結果與王湘南等(2011)、朱雯等(2016)分別對油茶(Camelliaoleifera)和廣寧紅花油茶(Camelliasemiserrata)劃分的花芽分化時期相類似,而與施雁飛等(2015)對茶(Camelliasinensis)的劃分結果存在較大差異,前者與孫建云等(2005)的觀點一致,將雌雄蕊成熟階段納入花芽形態分化的范圍,認為花藥、花絲的分化以及子房(胚珠、胚囊)、花柱、柱頭的分化和發育是雌雄蕊原基繼續分化的結果,是非常重要的階段。
鈍葉柃花芽形態分化始于8月,至12月中下旬基本完成5個階段的分化,其中雌雄蕊成熟期歷時相對較長,達35~40天。發育完成后,花芽仍處于休眠狀態,到次年的2—3月進入始花期,而在同屬細枝柃(Euryaloquaiana)中并未出現這一現象(巴羅菊,2017);花芽進入始花期的早晚可能與開花前一段時間內的平均氣溫、日照長短密切相關,楊康明等(1989)也表明影響杭州桂花(Osmanthusfragrans)開花的主要氣象因子是開花前10天的平均氣溫以及最低氣溫。
真花學說提出,被子植物的花是由類似于裸子植物本內蘇鐵目(Bennettitales)兩性孢子葉球上排列的胚珠和花粉器官演化而來(陳之端等,1997;冷琴等,2001),植物的性別差異主要取決于花器官的發育,單性花是在兩性花的基礎上其雌性或雄性器官退化進化而來(Aryaletal.,2014)。劉亭亭(2017)認為不同種類的花雖然在結構上存在差異,但其花原基發生的早期階段可能是一致的,而結構相似的花在花器官發育的某些階段也可能存在較大差異。本研究也發現,3種不同性別花芽在不同的分化時期其內部結構變化既存在相同之處,也有所差異。在雌雄蕊原基產生之前3種不同性別花芽的外部形態(長寬比、色澤)及內部組織結構的變化基本一致,花器官成熟時萼片和花瓣都為覆瓦狀排列,與山茶科植物其他類群的研究結果相吻合(張逸,2012);而在雌雄蕊分化期至雌雄蕊成熟期3種不同性別花芽各具特點,雌花芽中只有雌蕊原基生長發育,雄花芽和兩性變異花芽的雌雄蕊原基基本同時出現,雄花芽中雌蕊原基發育一段時間后停止分化,這可能是植物進化中兩性花向單性花進化的結果,此發育方式與細枝柃花器官分化類似,而與張瑞菊(2006)描述的柃木屬植物不符。兩性變異株的花芽中,少數花芽出現雄蕊或雄蕊原基發育不正常,這也體現鈍葉柃在進化過程中的不穩定,可能是控制花器官發生的某些基因在進化過程中,受環境、激素等因子選擇性表達的結果。近年來,相關研究已經取得了一定的成果,在某些物種中,阻滯發生在花芽分生組織的細胞分化階段,使其不能產生花器官原基(雄蕊或心皮),如大麻(Cannabissativa)花中不存在任何異性器官的痕跡(Calderon-Urrea,1993)。而在某些物種中,這種阻滯發生在花器官發育后期,從而留下退化的異性器官,比如番木瓜(Caricapapaya)雄花中仍然有心皮的存在,雌花中無雄蕊(Mitchelletal.,2005),與本文研究結果一致。
鈍葉柃雌花芽、兩性變異花芽為中軸胎座、倒生胚珠,與近緣屬代表種豬血木(Euryodendronexcelsum)、厚葉紅淡比(Cleyerapachyphylla)、厚皮香(Ternstroemiagymnanthera)的結果一致;但雄蕊原基的發育方式與近緣屬代表種有所差異,鈍葉柃中雄蕊原基單個發育分化成花藥和花絲,與細枝柃類似,而細齒葉柃(Euryanitida)、豬血木、厚皮香的雄蕊以雄蕊束的方式分化(巴羅菊,2017;張瑞菊,2006),這一差異可作為區分不同類群的依據。
4 結論
對鈍葉柃雌花芽、雄花芽和兩性變異花芽的分化和發育進行了詳細的細胞形態學觀察,不同性別花芽分化時期可劃分為苞片分化期、萼片分化期、花瓣分化期、雌雄蕊分化期、雌雄蕊成熟期,花芽性別分化的關鍵時期為雌雄蕊分化期,其兩性變異花芽的外部形態變化與雌花芽一致,內部結構發育方式與雄花芽相似,雌雄蕊發育完全,但發育過程也存在不穩定性,少數雌、雄蕊出現不同程度的退化,只具有單性花功能。研究結果為今后探討鈍葉柃性別分化的控制機制以及性別遺傳和性別鑒定提供理論依據。
