山羊MC1R基因克隆、表達及其多態性研究
2020-03-14 13:18:32李隱俠孟春花王慧利曹少先
江蘇農業科學 2020年1期
李隱俠, 張 俊, 錢 勇, 孟春花, 王慧利, 鐘 聲, 曹少先
(江蘇省農業科學院畜牧研究所/江蘇省農業種質資源保護與利用平臺,江蘇南京 210014)
MC1R(melanocortin-1 receptor,MC1R)基因,也稱促黑素細胞激素受體(MSHR)基因,在哺乳動物中由毛色擴展(extension)位點編碼,為G蛋白偶聯受體黑素皮質素受體(melanocortin receptors,MCRs)超家族成員之一,只有1個編碼區,編碼蛋白有7個跨膜結構域[1-2]。研究表明,MC1R基因是控制動物黑色素合成的重要基因,其與編碼刺鼠信號蛋白(Agouti)的基因一起調控動物色素合成的數量和分布,從而對動物的毛色形成起到非常重要的作用[3-4]。
研究表明,MC1R基因位點的多態性與哺乳動物包括大鼠[2]、牛[5]、馬[6]、豬[7]、綿羊[3]、山羊[8]毛色變異都有很大的相關性。在家畜中,沼澤水牛與白沼澤水牛在476、618、881、930、931 bp位點上分別發生了T/C、G/C、G/A、G/A、A/G突變,導致沼澤水牛和白沼澤水牛第159位氨基酸由絲氨酸變成苯丙氨酸,第310位氨基酸由谷氨酸變成丙氨酸,第294位氨基酸由天冬氨酸變成丙氨酸,發生了非同義突變[5]。在綿羊中MC1R位點鑒定出2個等位基因ED和E+[9-10],Vage等研究表明,MC1R中 p.M73K 和p.D121N位點是控制黑色性狀顯著的等位基因位點(ED),它們的突變導致綿羊黑色表型的形成[11];在杜洛克豬中的研究表明,A240T突變與其紅色毛色相關[12],Fontanesi等在6個不同毛色的山羊中共檢測到MC1R基因5個單核苷酸突變,并且發現MC1R基因可能決定了真黑色素和褐色素的表型[13]。
研究表明,MC1R基因表達量與動物被毛顏色及膚色存在關聯。林宇紓等研究發現,MC1R基因在摩拉水牛、沼澤水牛和黃牛皮膚組織中的相對表達量均顯著高于白沼澤水平,且白沼澤水牛MC1R基因的編碼區發生氨基酸位點突變,推測白沼澤水牛體內合成的黑色素缺失而導致毛色白化[5];何蕊純等研究了MC1R基因在3種色型烏蘇里貉皮膚組織中表達發現,MC1R基因在野生型烏蘇里貉皮質組織中蛋白表達量顯著高于紅褐色和白色型,說明MC1R在烏蘇里貉皮組織中的表達量與烏蘇里色型間存在一定的相關性[14]。在羊駝中,棕色皮膚中MC1R基因的表達量遠高于白色被毛,且差異極顯著[15]。
蘇淮山羊是波爾山羊和徐淮山羊的雜交后代,采用傳統雜交育種技術和現代分子輔助育種技術相結合,培育出白色被毛、繁殖力高、生長速度快、適應性強的肉用山羊新品種,目前已橫交第2個世代[16]。本研究以蘇淮山羊和波爾山羊為研究對象,分析MC1R基因在山羊中的序列特征,挖掘在2種不同毛色山羊中MC1R基因SNPs位點差異及表達量的差異,為白色蘇淮山羊的進一步選育提供一定的理論依據。
1 材料與方法
1.1 試驗材料
130只白色蘇淮山羊和100只波爾山羊由江蘇省農業科學院六合動物實驗基地統一飼養管理,營養及管理水平保持一致。每個個體采集耳組織樣,置液氮中帶回實驗室,-80 ℃ 保存。組織樣DNA的提取采用酚/三氯甲烷抽提法,TE緩沖液溶解后-20 ℃保存備用。RNA提取試劑盒提取RNA,TaKaRa一步法逆轉錄cDNA備用。
1.2 PCR擴增
根據GenBank數據庫中山羊MC1R基因序列(序列號為FM212940),采用Primer Premier 5.0軟件設計引物P1,擴增蘇淮山羊MC1R基因編碼區全序列。以蘇淮山羊DNA池為模板進行PCR擴增,引物退火溫度和片段長度見表1。PCR反應總體系20 μL,含模板DNA 60 ng、Taq聚合酶(5 U/μL)0.2 μL、dNTP(10 mmol/L)0.5 μL、引物(100 μmol/L)各 0.5 μL、MgC12(25 mmol/L)1.4 μL、10×緩沖液2 μL,添加滅菌雙蒸水至 20 μL。PCR反應程序:94 ℃預變性5 min;94 ℃變性30 s,退火30 s,72 ℃延伸30 s,35個循環;72 ℃延伸10 min。
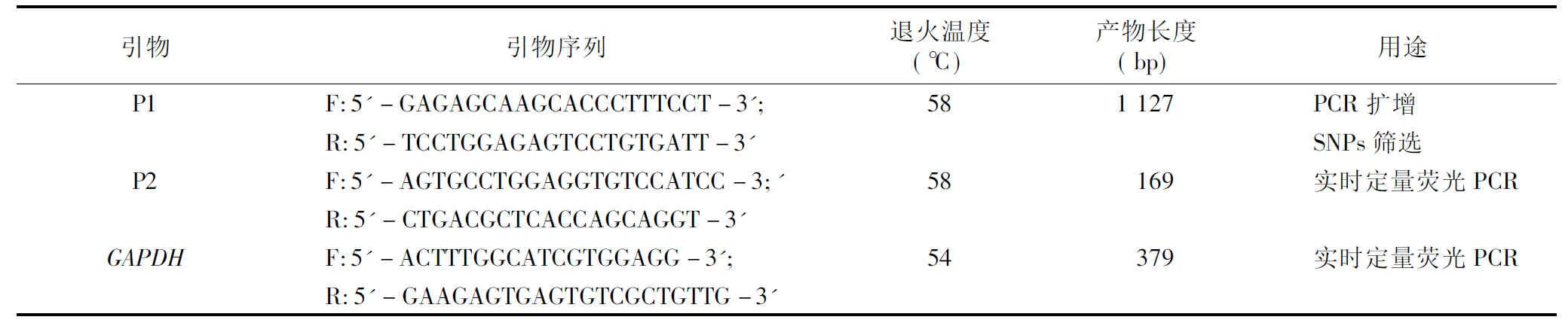
表1 引物序列及PCR反應參數
1.3 實時定量熒光PCR
根據GenBank數據庫中山羊MC1R基因序列(序列號為FM212940)和綿羊GAPDH(3-磷酸甘油醛脫氫酶,序列號為AF030943),采用Primer Premier 5.0軟件設計引物P2和GAPDH,以看家基因GAPDH為內參,實時定量熒光PCR方法檢測MC1R基因在2種山羊耳組織中的表達水平。反應總體系20 μL,含模板DNA 60 ng、SYBR Premix ExTaqⅡ 10 μL、引物(100 mol/L)各0.8 μL,添加滅菌雙蒸水至20 μL。PCR反應程序:95 ℃預變性 5 min;95 ℃變性10 s,60 ℃ 30 s,35個循環。
1.4 SNP篩選
PCR擴增產物用1%瓊脂糖凝膠電泳檢測,切膠后用DNA膠回收試劑盒回收目的DNA片段,直接送往上海捷瑞生物公司測序。根據序列峰圖,篩選出SNPs。
1.5 數據分析
利用SPSS 11.5軟件中的One-Way ANOVA統計不同組織中表達量的差異水平。
2 結果與分析
2.1 蘇淮山羊MC1R基因擴增及序列分析
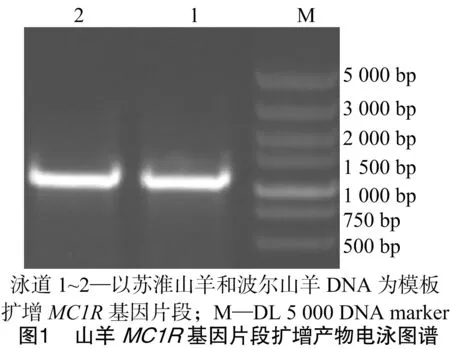
以蘇淮山羊DNA池為模板對MC1R基因進行PCR擴增,擴增產物經1%瓊脂糖凝膠電泳檢測發現擴增效果良好(圖1),切膠回收后直接進行測序。測序結果顯示,擴增片段長度與預期結果一致,全長1 127 bp。
將擴增獲得序列與引物源序列進行比對,通過DNA STAR軟件分析得到蘇淮山羊MC1R基因的編碼區全序列954 bp,編碼317個氨基酸殘基。通過DNA MAN序列比對發現,2個品種山羊MC1R基因編碼區核苷酸序列和氨基酸序列的一致性分別為99.79%和98.77%;核苷酸c.676 A>G突變和 c.701A>G 突變分別導致了氨基酸由谷氨酸到賴氨酸的變化和甘氨酸到天冬氨酸的改變(圖2)。蘇淮山羊MC1R基因與人類、小鼠、大鼠、牛、豬和綿羊的核苷酸一致性分別為 85.57%、79.65%、81.62%、96.05%、86.63%和98.55%,氨基酸的一致性分別為81.60%、75.15%、78.22%、95.09%、82.07%和98.16%,與家禽類核苷酸和氨基酸的一致性為73.52%和61.66%,說明MC1R基因序列在哺乳動物中的保守性較好。2個不同山羊品種MC1R基因序列中A、T、C、G 4種堿基的平均含量分別為14.8%、22.3%、36.6%、26.3%,其中A+T為37.1%,G+C為62.9%,可見AT含量遠低于GC含量。

2.2 哺乳動物MC1R系統發育分析
以家禽為外類群,采用MEGA 4.1軟件中NJ法構建了哺乳動物MC1R基因編碼蛋白(表2)的系統發育樹, 結果(圖3)發現聚類結果與經典分類學結果基本一致。2個山羊品種與7個哺乳動物聚在一起,而外類群家禽自成一類;在哺乳動物中,偶蹄目中的2個山羊品種、1個綿羊品種與牛聚在一起[自舉率(BP)=100%],后再與豬聚為一類(BP=72%);嚙齒目中的大鼠和小鼠聚為一類(BP=100%),而靈長目中的人類和黑猩猩聚為一類(BP=100%)。在偶蹄目中,牛科中的2個山羊品種與綿羊先聚在一起(BP=90%)后與牛聚為一類(BP=100%),豬科中的豬單獨聚為一類。在靈長目中,人科中的人和黑猩猩聚為一類(BP=99%)。
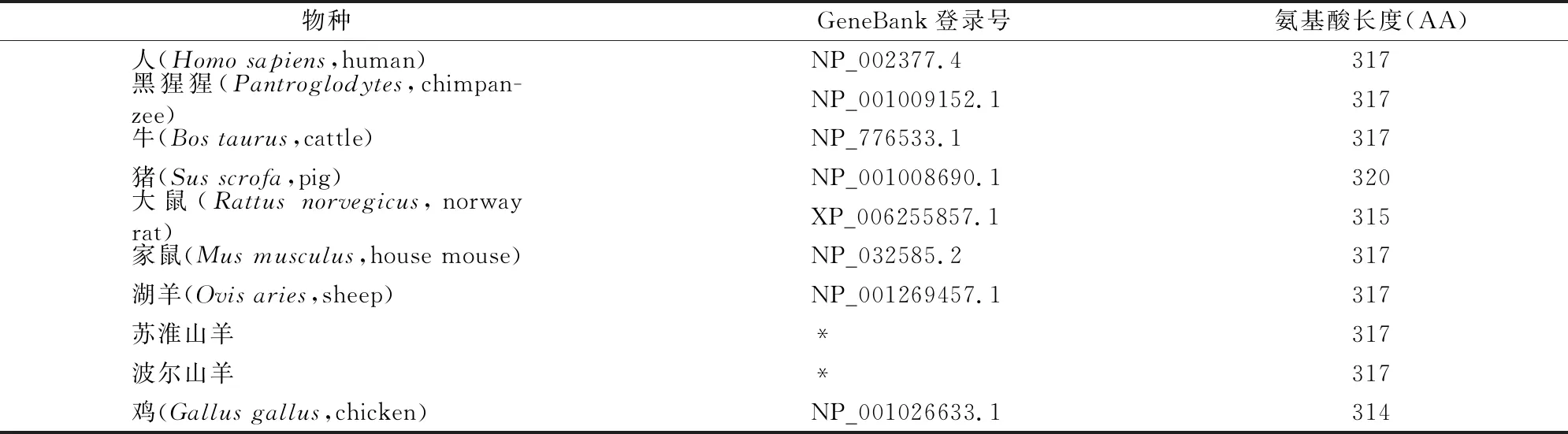
表2 部分哺乳動物MC1R氨基酸序列信息

2.3 MC1R基因在山羊耳組織中表達水平
為進一步研究MC1R基因在山羊毛色中的作用,用實時定量熒光PCR方法檢測MC1R基因在白色蘇淮山羊和紅棕色波爾山羊耳組織中的表達水平。由圖4可知,紅棕色耳朵的波爾山羊的MC1R表達水平高于蘇淮山羊,但是差異并不顯著(P>0.05)。

2.4 山羊MC1R基因編碼區SNPs篩選和分析
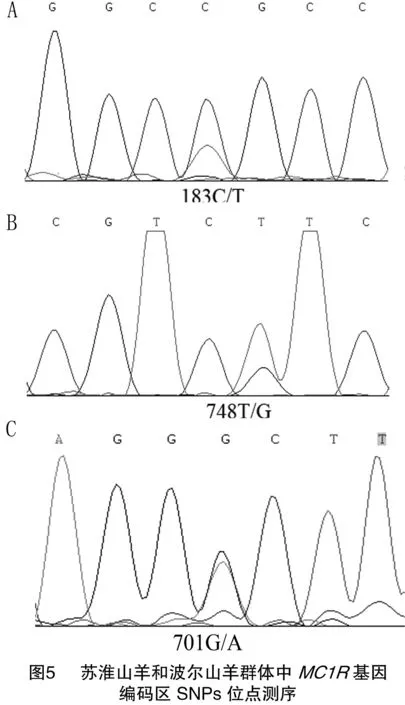
以2個山羊品種(波爾山羊和蘇淮山羊)DNA池為模板,利用引物P1對MC1R基因編碼區進行PCR擴增,擴增產物經瓊脂糖凝膠電泳檢測回收后直接進行測序。根據測序峰圖,在2個山羊品種MC1R基因編碼區中共發現3個SNP位點,蘇淮山羊群體中發現2個SNPs位點,即183C/T和748T/G(圖5-A、B),波爾山羊群體中發現了1個SNPs位點,701G/A(圖5-C),其中748T/G位點突變引起了苯丙氨酸到頡氨酸的突變,701G/A位點突變引起了甘氨酸到天冬氨酸的改變。
3 結論與討論
MC1R基因是1個編碼7次跨膜蛋白的G蛋白受體,被認為是動物毛色和皮膚顏色變化的重要候選基因,可與內源配體促黑激素或者刺鼠信號蛋白結合,調節動物的毛色和膚色[4,17]。
本研究以白色被毛的蘇淮山羊和紅棕色頭頸部的波爾山羊為研究對象,蘇淮山羊是利用波爾山羊和徐淮山羊雜交后代,采用傳統雜交育種技術和現代分子輔助育種技術相結合,培育出白色被毛、繁殖力高、生長速度快、適應性強的肉用山羊新品種[16]。本研究得到蘇淮山羊和波爾山羊MC1R基因編碼區長度954 bp,編碼317個氨基酸殘基,同源性比對發現MC1R基因在哺乳動物間相對保守,與綿羊的氨基酸序列一致性最高,為98.16%,牛的次之(95.09%),與雞的同源性僅為61.66%。在蘇淮山羊和波爾山羊品種間核苷酸的一致性和氨基酸的一致性都很高,分別為99.79%和98.77%;核苷酸c.676的A>G突變和c.701A>G突變分別導致氨基酸由谷氨酸到賴氨酸的變化和甘氨酸到天冬氨酸的改變。波爾山羊的紅棕色耳朵和蘇淮山羊白色耳朵是1對不同的毛色性狀,而本研究發現蘇淮山羊和波爾山羊間MC1R基因編碼區有2個可以引起氨基酸改變的SNPs位點存在,這可能與二者之間毛色不同的性狀相關。
以前的研究表明,MC1R基因的表達量與動物的被毛和膚色存在一定的關聯[18]。本研究在2種山羊不同被毛的耳組織中MC1R基因表達水平也存在差異,紅棕色波爾山羊的表達水平高于白色蘇淮山羊表達水平,雖然差異不顯著,但也可在一定水平上反映不同毛色間MC1R基因表達水平是不同的,這與林宇紓等的研究結果[5]一致。這些結果間接說明MC1R基因可能參與蘇淮山羊白色被毛形成的分子調控。
到目前為止,關于MC1R多態性與動物毛色關聯性的研究很多,在哈薩克綿羊中發現MC1R基因T218A突變(p.M73K)的AB基因型與不同被毛顏色顯著相關[18];Fontanesi等在對波爾山羊的研究中發現,MC1R基因第226個核苷酸位置由賴氨酸向谷氨酸的轉變與波爾山羊頭部和頸部的紅色毛有關[13];本研究中在波爾山羊和蘇淮山羊群體MC1R基因編碼區篩選到3個SNPs位點,其中183C/T和748T/G只存在于白色被毛的蘇淮山羊群體中,701G/A位點僅存在于棕色頭部的波爾山羊群體中,在白色被毛的蘇淮山羊群體中未見該突變位點,說明波爾山羊中701G/A位點可能與其棕色頭部被毛有關,這些位點是否與山羊毛色形成有關聯還需要更多毛色的山羊進行進一步的驗證。
