遼寧地區遷飛黏蟲種群動態與發生規律研究
2020-03-09 02:50:34范唯艷許國慶趙彤華
環境昆蟲學報 2020年1期
鐘 濤,范唯艷,許國慶*,趙彤華
(1. 遼寧省農業科學院植物保護研究所,遼寧省農作物有害生物控制重點實驗室,農業部沈陽作物有害生物科學觀測實驗站,沈陽 110161;2. 遼寧省農業科技成果轉化服務中心(遼寧農業博物館),沈陽 110161)
黏蟲Mythimnaseparata(Walker)又稱東方黏蟲,是我國農業生產中的重要遷飛性害蟲,其發生具有突發性、暴食性、聚集性和雜食性等顯著特點。黏蟲在我國每年發生6~8代,其中4~5次為大范圍跨區域的遷飛活動(江幸福等,2014;姜玉英等,2014)。受季風和寄主成熟度的影響,一代黏蟲借助上升氣流起飛,伴隨西南氣流向東北地區定向遷移(高月波等,2014a;高月波等,2014b;張麗等,2014)。到達遷入地上空遭遇下沉氣流降落當地,條件適宜則定殖,若環境不利,可再隨氣流繼續遷飛。黏蟲蛾晝伏夜飛,夜間可連續飛行(王玉正和張孝羲,2001;江幸福等,2005)。降落區域有蜜源植物生長,對黏蟲蛾迅速補充能量有利,從而結束遷飛轉向生殖發育(曹雅忠等,1995;李克斌等,2005;呂偉祥等,2014;王娟等,2016)。有研究表明種群中80%以上黏蟲蛾卵巢發育級別為3~4級,可認定為遷入種群,反之,為遷出種群(孫金如,1990;江幸福和羅禮智,2005;江幸福等,2014)。黏蟲喜食禾本科作物,玉米、水稻、小麥、谷子等受害重(屈麗莉和郭英偉,2014;趙彤華等,2016)。東北地區6月份麥田和玉米田郁閉程度高,溫濕度適宜,食料充足,成為一代黏蟲遷入的重要區域(王寧等,2014)。遼寧地區黏蟲寄主種類多,各地區發生普遍。黏蟲大發生年份,低齡幼蟲進入暴食期,植株葉片一夜間被食光,僅剩莖稈和葉脈,對秋糧生產威脅極大(范垂鵬等,2012;曲昌明,2013;屈麗莉和郭英偉,2014)。2012年8月遼寧省臺安縣黏蟲大暴發,玉米基本絕收。就是由于二代蛾外遷受阻,導致三代黏蟲幼蟲大量聚集造成的生物災害(晉齊鳴等,2013;王寧等,2014;張麗等,2014;叢勝波等,2015;孫嵬等,2018)。東北地區冬季氣溫低,黏蟲無法越冬,必須隨氣流南遷至南方越冬區(胡高等,2014;姜玉英等,2014;李鴻波等,2017)。7、8月份受寄主成熟度影響,二代黏蟲蛾羽化后隨氣流陸續遷出(江幸福等,2005)。羽化后1~2天是二代蛾種群外遷高峰(江幸福等,2014)。
黏蟲在我國的越冬區域基本上穩定在30°N以南的西南和華南地區,但隨著全球氣候變暖效應加劇,黏蟲越冬界逐漸北延(江幸福等,2014;姜玉英等,2014;叢勝波等,2015;毛永凱等,2016;李鴻波等,2017)。越冬區域的擴大勢必造成黏蟲蟲源基數進一步累積,使東北地區二、三代黏蟲的暴發概率和發生面積較過去均有明顯的增加(江幸福等,2014;姜玉英等,2014)。因此,開展該地區長期性監測工作,分析黏蟲遷入和遷出種群發生動態對黏蟲的區域防控和異地測報有著重要的指導意義。當前,遼寧地區有關黏蟲發生規律的報道較少,主要圍繞黏蟲大發生后的原因探討方面(范垂鵬等,2012;曲昌明,2013;屈麗莉和郭英偉,2014;趙彤華等,2016)。目前各地區黏蟲監測主要依靠高空探照燈、黑光燈和蟲情測報燈(江幸福等,2014;叢勝波等,2015)。因此,本研究利用田間架設的高空探照燈和黑光燈,適時開展田間調查,分析了黏蟲在遼寧沈陽和岫巖地區的種群消長動態和蟲情發生發展規律,旨在為遼寧省黏蟲的區域防控與協同治理提供科學參考。
1 材料與方法
1.1 監測地概況
監測地選擇遼寧省沈陽市(123°33′E, 41° 49′N)和岫巖縣(123° 20′E, 40° 13′N),以小麥、玉米為主要監測作物,連片種植總面積約0.67 hm2。監測地地勢平坦,栽培和管理水平一致,黏蟲每年均有不同程度發生。小麥品種為“遼春18號”,玉米品種為“丹玉4955”。
1.2 試驗材料
高空探照燈(時控)固定于遼寧省農科院試驗基地小麥田附近(叢勝波等,2015)。頻振式太陽能誘蟲燈(光控,由富巍盛科技有限公司生產,型號為FWS-DBL-2)。在岫巖縣農業技術中心聯合示范基地架設頻振式太陽能誘蟲燈1臺,固定于玉米田附近。
1.3 試驗方法
1.3.1黏蟲蛾種群發生動態監測
根據以往監測研究,黏蟲蛾本地遷入日期最早為5月中旬,二代或三代黏蟲蛾本地遷出時間最遲為9月底(李光博等,1964)。因此,高空探照燈啟用時間為4月20日,至10月31日關閉,每天20 ∶30自動亮燈,至凌晨4 ∶30關閉,監測時長8 h。燈下接一個塑料桶,內盛清水。頻振式太陽能誘蟲燈(雙燈型)于4月20日開啟,至10月31日結束。燈下各放置1個塑料盆,內盛清水。每天對燈下誘集的昆蟲進行分類、記錄,拍照,保存黏蟲蛾標本。
1.3.2黏蟲蛾卵巢發育成熟度調查
對沈陽和岫巖地區每天燈誘黏蟲蛾進行卵巢解剖,觀察和記錄卵巢發育級別。并根據前述對黏蟲卵巢發育級別的研究結果,判定遷入種群或遷出種群。
1.3.3一代黏蟲蛾產卵調查
在誘蟲燈監測到黏蟲蛾后,將干稻草捆扎成枯草把,固定在竹竿頂部,始終保持稻草把頂端高于周邊作物20 cm以上。在小麥田間隔10 m插一草把,共插10把。田間蛾量觀測從5月20日至6月20日結束。自5月23日開始,觀測每隔3 d在夜間19 ∶30-21 ∶30進行。間隔15 min向稻草把均勻噴灑糖酒醋液(配比3 ∶1 ∶4 ∶2)一次,在弱紅光下記錄黏蟲蛾前來取食情況。第二天上午調查草把上卵塊數,同時隨機選取10塊卵,調查卵粒數,不足10塊的,全部調查。
1.3.4二代黏蟲幼蟲發生調查
分別調查田邊雜草、小麥田、玉米田、谷子地黏蟲幼蟲發生情況。每隔2 d調查1次,發現黏蟲幼蟲,即按照對角線五點取樣法,每點調查10株,記錄齡期、蟲量、植株葉片是否被幼蟲取食等。
1.4 數據處理
利用Microsoft Excel 2003及IBM SPSS Statistics 24.0軟件對數據進行分析并繪圖。
2 結果與分析
2.1 高空探照燈監測黏蟲蛾發生動態
沈陽地區2017年5月14日利用高空探照燈首次監測到黏蟲蛾遷入(圖1)。黏蟲蛾發生有2個主要發生時期,5月下旬到6月底比較集中,根據卵巢的解剖結果,判定為一代蛾遷入期;7月中旬至8月初為二代蛾發生期。6月6-9日黏蟲蛾數量急劇增加,單日最大上燈蛾量為9頭。此后,黏蟲蛾數量下降。至7月上旬,黏蟲蛾數量又逐漸增加。至8月4日,為監測到的黏蟲蛾第二個發生高峰,單日最大上燈蛾量為5頭。10月中旬仍能監測到黏蟲蛾,但蛾量一直很低。整個監測期間共誘集黏蟲蛾91頭,其中雌蛾63頭,雄蛾28頭,雌雄性比為2.25 ∶1。一代蛾遷入種群共誘集42頭,占總誘蛾量的46.2%;二代蛾發生期共誘集36頭,占總蛾量的39.6%。三代黏蟲蛾零星上燈,僅占上燈蛾量的14.2%。應為受降雨影響迫降本地的黏蟲蛾過境種群或本地外遷受阻種群,且此階段上燈蛾量較前兩次高峰均明顯下降。10月 12日為全年最后一次監測到黏蟲蛾的時間。

圖1 2017年沈陽地區高空探照燈誘集蛾量動態變化Fig. 1 Population dynamic of Mythimna separata under vertical-pointing searchlight-traps in Shenyang (2017)
2018年監測黏蟲蛾首次遷入日期為5月11日。一代蛾遷入期為5月中下旬至6月上旬,高峰蛾量出現在5月底至6月上旬;6月下旬至7月上旬為二代蛾發生期。5月30日黏蟲蛾達到18頭峰值(圖2)。此后,黏蟲蛾數量開始波動。6月20日,上燈蛾量明顯增加,為監測到的第二個發生峰期,單日最大上燈蛾量為12頭。2018年共誘集黏蟲蛾212頭,其中雌蛾148頭,雄蛾64頭,雌雄性比為2.31 ∶1。一代蛾共誘集126頭,占總誘蛾量的59.4%;二代蛾發生期共誘集71頭,占總蛾量的33.5%。三代蛾發生期從7月24日至8月25日,僅占上燈蛾量7.1%。
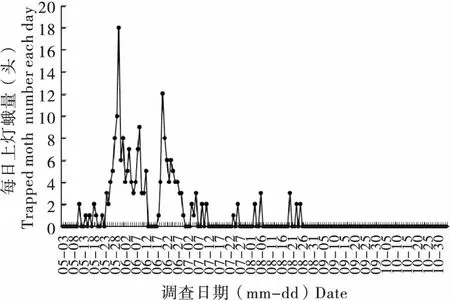
圖2 2018年沈陽地區高空探照燈誘集蛾量動態變化Fig.2 Population dynamic of Mythimna separata under vertical-pointing searchlight-traps in Shenyang (2018)
2.2 黑光燈監測黏蟲蛾量發生動態
2017年岫巖地區利用黑光燈監測黏蟲蛾,也監測到2個發生期,一代蛾遷入時期為5月底至6月上旬,二代蛾發生期為7月上旬至8月上旬。利用黑光燈首次監測黏蟲蛾日期為5月30日。5月 30日至6月9日為黏蟲蛾遷入高峰,單日最大上燈蛾量3頭。此后,黏蟲蛾數量下降。7月8日再次監測到有黏蟲蛾上燈活動,至8月13日為黏蟲蛾第二個發生期,單日最大上燈蛾量2頭(圖3)。整個黑光燈監測期間共誘集黏蟲蛾25頭,其中雌蛾20頭,雄蛾5頭,雌雄性比為4 ∶1。一代蛾遷入種群共誘到13頭,占總誘蛾量的52.0%;二代蛾發生期共誘集12頭,占總蛾量的48.0%。
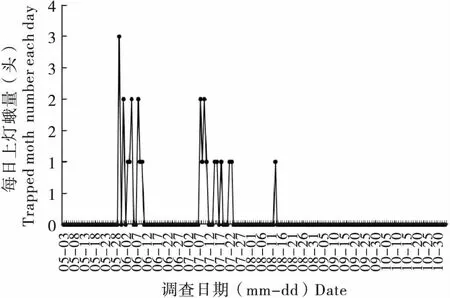
圖3 2017年岫巖地區黑光燈誘集蛾量動態變化Fig.3 Population dynamic of Mythimna separata under black light lamp in Xiuyan (2017)

圖4 2018年岫巖地區黑光燈誘集蛾量動態變化Fig.4 Population dynamic of Mythimna separata under black light lamp in Xiuyan (2018)
2018年黑光燈監測到一代黏蟲蛾遷入時期為5月 15日至6月14日,二代蛾發生期為7月15日至7月26日(圖4)。5月31日至6月11日為黏蟲蛾遷入高峰期,單日最大上燈蛾量5頭。整個監測期間黑光燈共誘集黏蟲蛾60頭,其中雌蛾49頭,雄蛾11頭,雌雄性比為4.45 ∶1。一代蛾遷入種群共誘到31頭,占總誘蛾量的51.7%;二代蛾發生期共誘到16頭,占總蛾量的26.7%。
2.3 黏蟲蛾種群性質分析
2017年監測期間,沈陽和岫巖兩地放置的誘蟲燈共收集黏蟲蛾116頭,解剖發現其中一代雌蛾卵巢3級為34頭,4級為5頭;二代蛾卵巢1級為22頭,2級為3頭(圖5)。該結果表明2017年遼寧沈陽和岫巖地區一代蛾均為遷入種群,二代蛾為遷出種群。
2018年監測期間,沈陽和岫巖兩地放置的誘蟲燈共收集黏蟲蛾272頭,其中一代雌蛾卵巢3級為64頭,4級為11頭;二代蛾卵巢1級為79頭,2級為43頭(圖6)。該結果表明2018年遼寧沈陽和岫巖地區一代蛾均為遷入種群,二代蛾則為遷出種群。
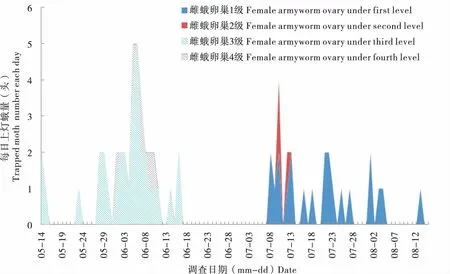
圖5 2017年監測黏蟲蛾種群卵巢發育級別分析Fig.5 Ovary development of Mythimna separata under light in Shenyang and Xiuyan (2017)
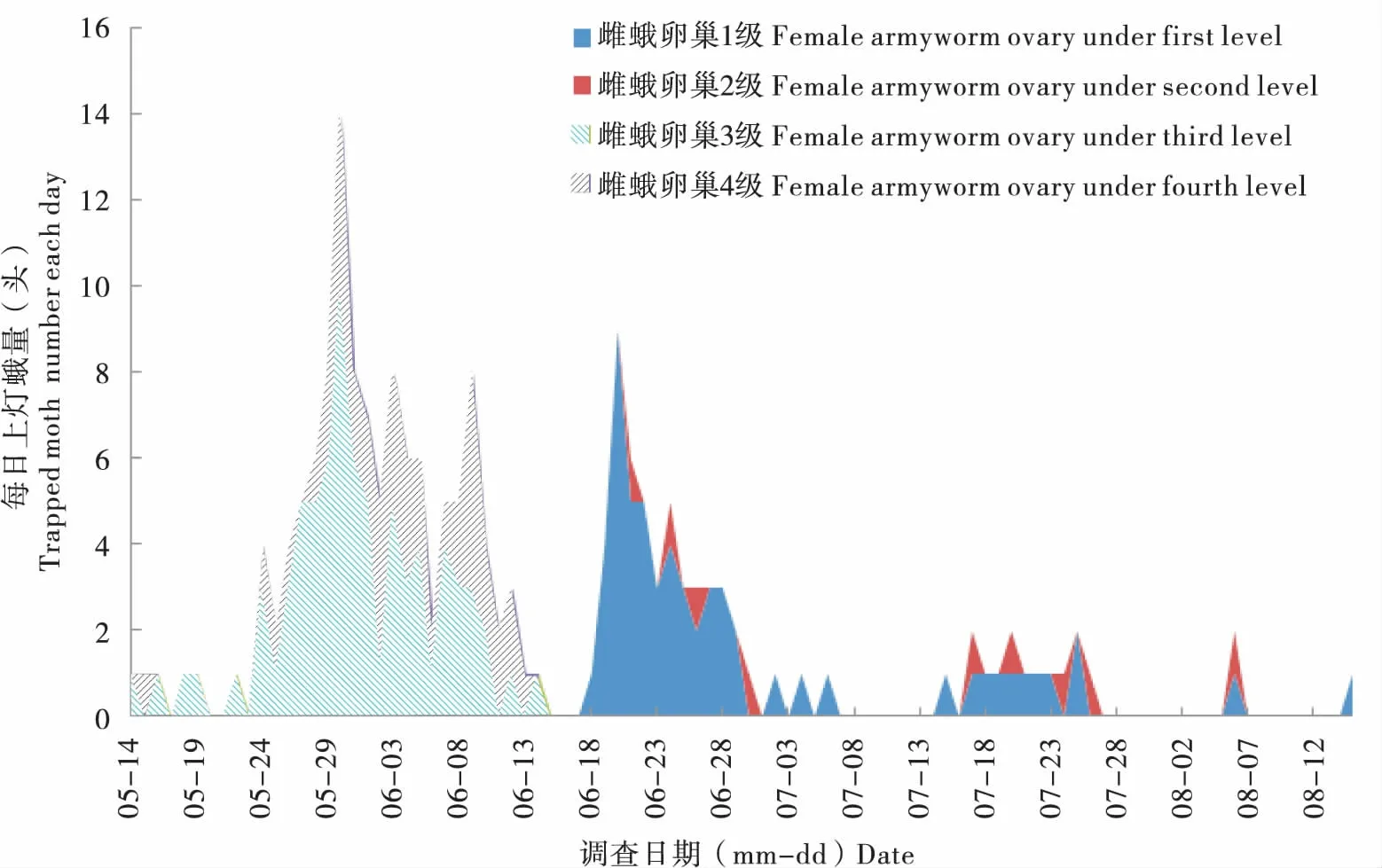
圖6 2018年監測黏蟲蛾種群卵巢發育級別分析Fig.6 Ovary development of Mythimna separata under light in Shenyang and Xiuyan (2018)
2018年沈陽地區氣象監測數據顯示(圖7),無論降水量及降水頻次都較2017年明顯增多。當降雨天氣發生時,日均氣溫多保持在20℃上下,本地氣象和環境條件對黏蟲蛾降落和定殖都十分有利。由于2017年5月下旬初降水少(5月22日至25日),且多為短時降雨,因此黏蟲蛾隨下沉氣流降落本地機會較2018年同期降水(5月22日至24日;5月27日至6月1日)要少很多。這樣就能合理地解釋了2018年燈誘粘蟲蛾量較2017年明顯增多的現象。2018年,二代蛾遷出期間強降雨(6月28日;7月6日;7月8日)對黏蟲蛾外遷活動非常不利,同時東北地區北部南遷種群受強降雨影響迫降沈陽地區。2018年8月份,降雨活動頻繁,氣溫迅速下降(20~25℃),這些原因共同加劇了三代黏蟲幼蟲暴發成災的風險。2017年8月間降雨情況類似,但是由于前期一代遷入黏蟲蛾量基數小,且各村鎮防治及時,因災損失程度低。
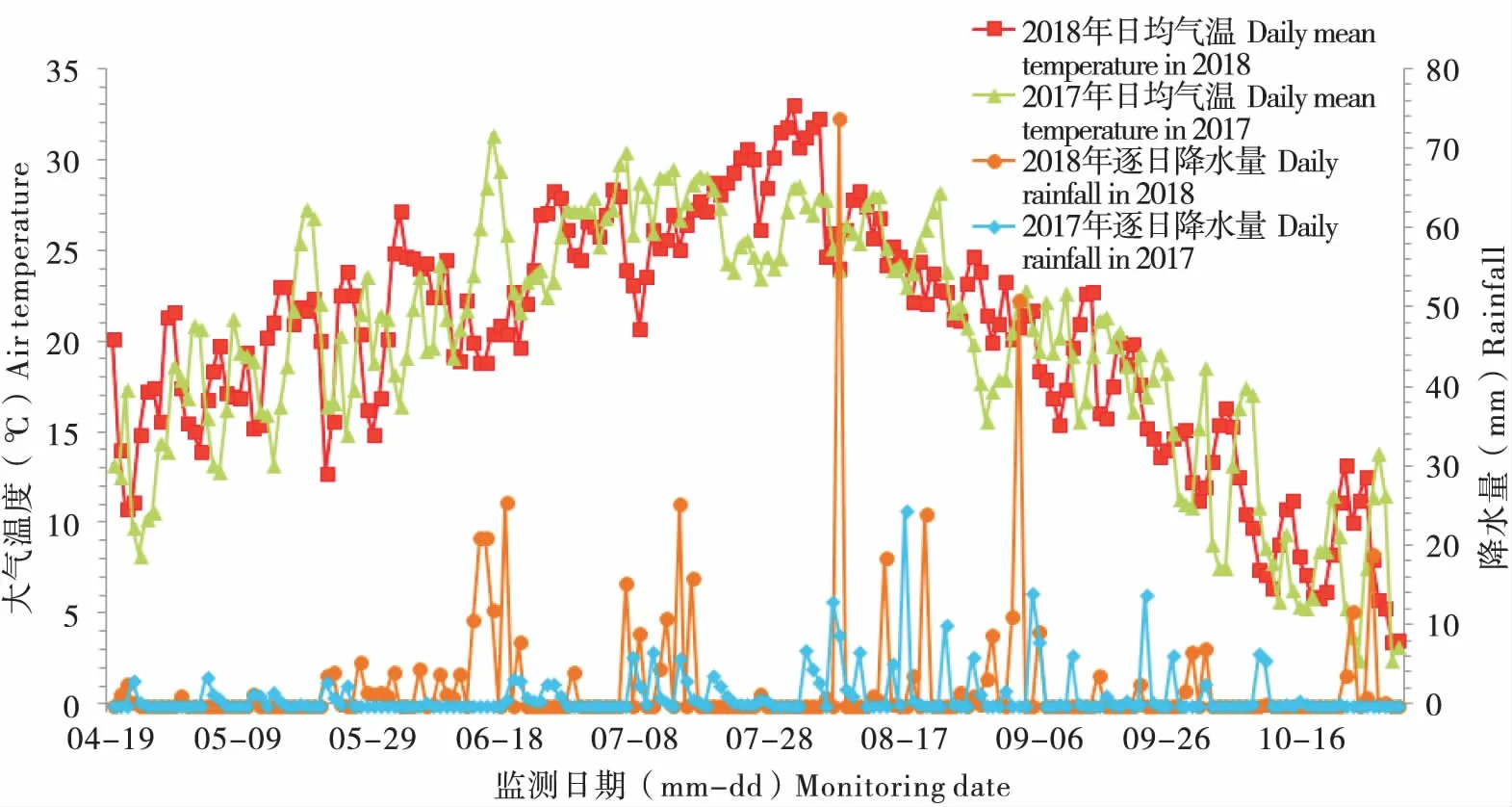
圖7 2017-2018年沈陽田間氣象監測數據Fig.7 Meteorological data of Shenyang between 2017 and 2018
2.4 黏蟲幼蟲種群發生監測
2017年6月5日開展麥田踏查,百步驚蛾量在57頭左右。6月6-9日,田間黏蟲蛾數量明顯增多。6月10日田間雜草上始見二代黏蟲幼蟲為害。玉米田受旱情影響,拔節期受到抑制,葉片卷曲,田間未調查到黏蟲幼蟲為害;而小麥已經抽穗灌漿,田間郁閉程度高,濕度較大,利于黏蟲蛾潛藏和產卵;此時,谷子地種子剛破土萌芽,田間基本處于裸露狀態,未調查到黏蟲幼蟲發生。麥田由于定期采取化學防治措施,麥蚜等害蟲基數受壓制較低,黏蟲存活蛾量每平方米僅為1頭以下,而且田間雜草上黏蟲幼蟲發生量也輕。夜間目測枯草把(噴灑糖醋液)誘蛾12頭/10把,黏蟲蛾全部來自附近麥田中。調查枯草把發現5塊卵,平均單塊卵137.6粒。
2018年5月30日開展麥田踏查,百步驚蛾達到120頭左右。6月11日麥田和雜草上始見二代黏蟲幼蟲為害。6月15日麥田調查,平均每株上黏蟲幼蟲發生量在2~4頭,蟲齡在1~3齡不等。此時玉米田和谷子地均未調查到黏蟲幼蟲為害,地頭雜草上也未見幼蟲取食。夜間目測枯草把(噴灑糖醋液)誘蛾26頭/10把,黏蟲蛾主要來自附近麥田。調查枯草把發現17塊卵,平均單塊卵為124.1粒。對比不同類型田二代黏蟲幼蟲發生量,可以發現不同年份麥田黏蟲幼蟲發生量均顯著高于玉米和谷子田;麥田雜草區最先受害,大量取食后幼蟲就近轉移麥田為害(表1)。2018年三代黏蟲幼蟲均高密度為害玉米田,谷子田受災也較2017年同期更重(表2)。
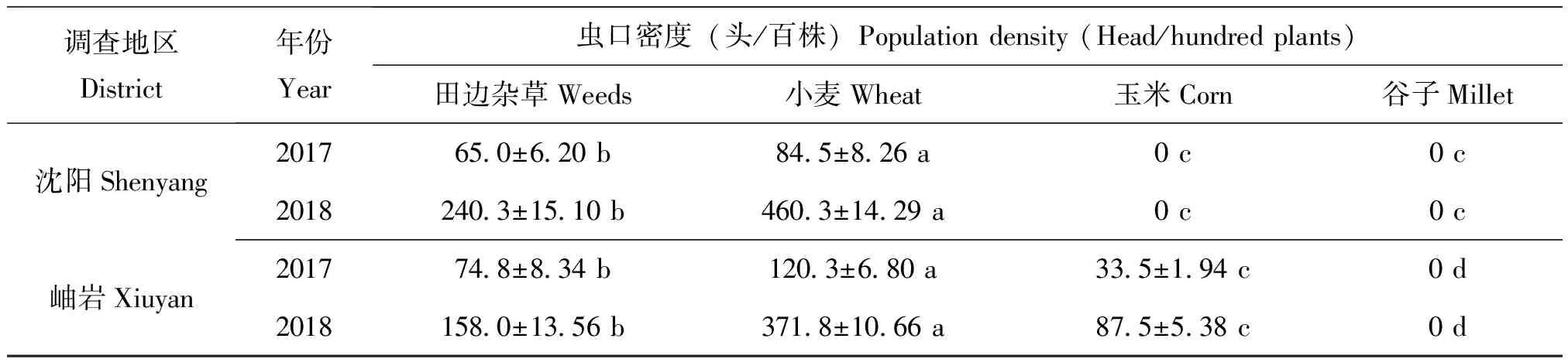
表1 二代黏蟲幼蟲田間調查結果(6月6-11日)
注:表內同行數據(平均值±標準誤)后小寫字母表示不同類型田黏蟲幼蟲蟲口密度的差異顯著性(P<0.05)。
Note: Values (mean±SE) within a row followed by lowercase letters indicated significant differences among the population density of larvae separately (P<0.05).
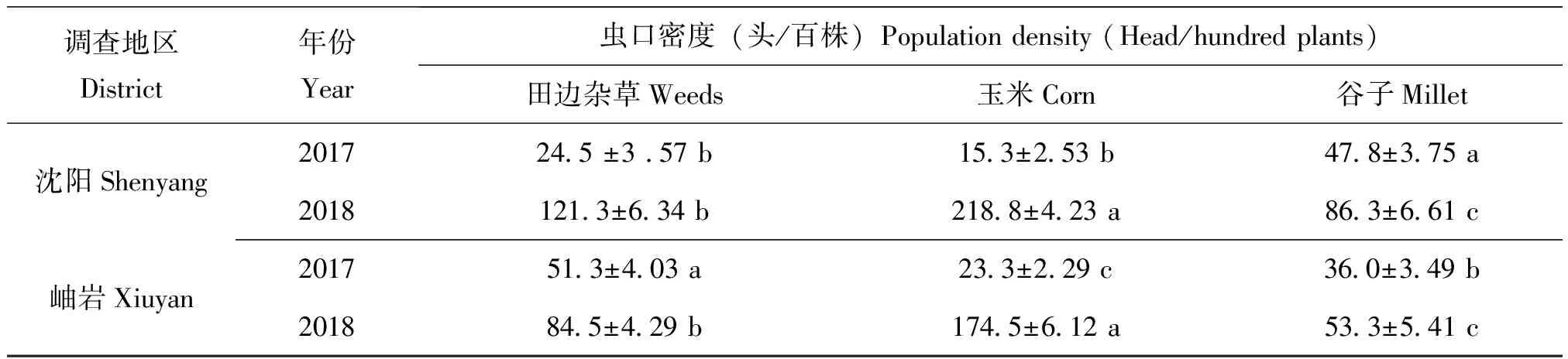
表2 三代黏蟲幼蟲田間調查結果(8月5-10日)
注:表內同行數據(平均值±標準誤)后小寫字母表示不同類型田黏蟲幼蟲蟲口密度的差異顯著性(P<0.05)。
Note: Values (mean±SE) within a row followed by lowercase letters indicated significant differences among the population density of larvae separately (P<0.05).
3 結論與討論
本研究利用高空探照燈和黑光燈分別對沈陽和岫巖地區2017-2018年遷飛黏蟲蛾活動進行了連續監測,其中高空探照燈監測黏蟲蛾動態顯示出明顯的波峰和波谷特征,證明遷飛至本地區上空的黏蟲蛾存在頻繁遷飛活動或飛行行為。有研究表明東北地區一代黏蟲蛾的遷入蟲源來自山東地區,尤其是長島觀測站監測到極大蟲量,黏蟲蛾隨西南暖濕氣流跨海遷入我國東北地區(張云慧等,2013;姜玉英等,2016)。黏蟲蛾遷入時期受當地降雨等氣象條件影響大,遷入沈陽和岫巖的時間相對集中,大致在5月30日至6月9日這段時間,與當地同期降雨時段完全吻合,這些發現與前人研究基本一致(范垂鵬等,2012;趙彤華等,2016)。在沈陽東陵地區,高空探照燈監測證實黏蟲蛾每年發生2~3個峰,其中三代蛾發生不規整,應為滯留本地活動的黏蟲蛾種群,規模不大。高空探照燈對空照射距離可達500 m,是目前監測黏蟲種群遷入時期的最為有效手段(呂偉祥等,2014;叢勝波等,2015;姜玉英等,2016)。受限于設備維護成本,本研究僅在沈陽地區設置1臺高空探照燈。岫巖利用黑光燈監測證實黏蟲蛾同樣每年發生2~3個峰,但監測結果多為遷入后定殖種群的飛行活動。
沈陽和岫巖監測到黏蟲蛾遷入時期基本一致,遷出時期不同年份間稍有所差別。高空探照燈監測到的黏蟲蛾發生時間要早于黑光燈,監測黏蟲蛾發生期也更持久一些。沈陽地區6月間發生的黏蟲蛾為遷入種群,7月下旬至8月上旬二代蛾羽化后,形成南遷種群陸續遷出。這些結果與前人研究結果大體相近(趙彤華等,2016;孫嵬等,2018)。2017年遼寧地區黏蟲偏輕發生,百株幼蟲量不足10頭。以沈陽為例,2017年6月間雨水偏少,旱情嚴重,對遷入本地的一代蛾的定殖活動將產生不利影響。盡管7月份降雨天數有所增多,但多為短時強降雨,持續高溫對黏蟲蛾上燈活動同樣產生了不利影響。8月份突降特大暴雨,引發遼寧多地洪澇災害,農作物受災,黏蟲受雨水沖刷種群密度下降。同時玉米接近成熟,葉片趨近老化,本地羽化的黏蟲蛾陸續遷出,也是本地區監測到的黏蟲蛾發生量偏少的原因。同年岫巖地區誘蛾量同樣偏少,農民防治及時,沒有造成較大危害。
2018年5月下旬,雷雨天氣較為集中,導致黏蟲蛾隨下沉氣流遷入沈陽地區。5月底田間調查時,黏蟲蛾已經普遍避入麥田,一代蛾發生量較大。6月中上旬雷雨天氣頻發,導致黏蟲蛾北遷種群持續遷入本地區。7月、8月中上旬降雨集中,本地種群外遷受阻,滯留本地繁殖。冷空氣南下活動也導致黑龍江、吉林南遷種群在遼寧地區迫降,因此2018年監測的黏蟲蛾量較往年均明顯增多。同時三代黏蟲幼蟲為害風險加大。這些結果與2018年東北地區二代、三代黏蟲發生偏重,局地黏蟲幼蟲高密度發生預測結果十分吻合(姜玉英等,2018)。對2018年沈陽和岫巖兩地誘集黏蟲的蟲源性質分析,一代蛾中卵巢發育3和4級占77.7%,二代蛾卵巢發育1和2級占86.2%,符合對遷入和遷出種群的認定。黏蟲蛾降落區域相對集中,聚集程度高,產卵危害程度大(羅禮智等,1995;高月波等,2014b)。雌蛾偏好禾本科雜草產卵,毗鄰雜草田塊往往受害較重(李瓊等,2008;姜玉英等,2014)。即使同一塊田受害也不均勻,田邊地頭受害較重。調查發現沈陽地區5月下旬春麥田處于孕穗期,田間郁閉程度高,濕度大,麥田成為一代黏蟲蛾遷入后棲息的首選場所。小麥成熟后,玉米田成為黏蟲蛾主要繁殖場所(叢勝波等,2015)。
研究表明溫濕度適宜、蟲源和食料充足是三代黏蟲暴發的重要原因(曹雅忠等,1995;姜玉英等,2014)。黏蟲暴發受環境氣象條件影響很大,降雨偏多年份黏蟲往往大發生(屈麗莉和郭英偉,2014)。2018年是繼2012年、2013年三代黏蟲大暴發后又一個高發年份。針對黏蟲暴發性危害特點,開展異地測報是實現全國黏蟲協同治理的最佳解決方案,比如華中或黃淮海麥區一代黏蟲羽化和遷出時間可為華北、東北地區黏蟲監測提供準確的遷入時期預報,東北地區要重點監測二代黏蟲蛾遷出時期(江幸福等,2014;毛永凱等,2016)。改善農田生態環境和增強天敵資源保護對黏蟲防治工作也至關重要(李文強等,2017),玉米與花生、大豆間作可以減輕大發生年份秋黏蟲為害(Midegaetal., 2018)。隨著遷飛黏蟲在我國不同區域和本地蟲源頻繁基因交流,黏蟲抗性問題突顯(陳曉燕等,2017)。國外已有黑化黏蟲為害轉基因玉米的報道(Silvaetal., 2016)。這恰好說明沈陽和岫巖地區開展黏蟲監測工作的必要性、重要性以及長期性,各觀測站應準確追蹤黏蟲遷入和遷出時期,結合異地測報信息網絡平臺在黏蟲高峰來臨前及時開展科學防治,以減少黏蟲引發的災害損失。
