植物類病變突變體的研究進展
2020-02-29 02:40:24劉思辰曹曉寧王海崗王君杰陳凌田翔秦慧彬喬治軍
山西農業科學 2020年2期
劉思辰,曹曉寧,王海崗,王君杰,陳凌,田翔,秦慧彬,喬治軍
(山西省農業科學院農作物品種資源研究所,農業部黃土高原作物基因資源與種質創制重點實驗室,雜糧種質資源發掘與遺傳改良山西省重點實驗室,山西太原030031)
植物類病變(lesion mimic)是指在無外界致病因素和逆境因素的影響下,植株葉片、葉鞘等部位自發產生壞死斑的現象。多種植物中均發現類病變突變體[1]。類病變突變與植物過敏性反應(hypersensitive response,HR)癥狀類似[2],可在類病變產生部位發生細胞壞死,屬于程序化細胞死亡(programmed cell death,PCD),而且許多斑點葉突變體對某些植物病原物表現出了一定的抗性[3]。因此,研究類病變突變體對了解細胞的程序性死亡以及植株抗病反應機制具有重要意義[4]。類病變突變體也是抗病種質資源的重要來源。在大麥中發現的類病變突變體mlo對白粉病生理小種都具有持久抗性[5],類病變突變體mlo在歐洲許多國家已經大面積種植。
本研究通過綜述植物類病變突變體的來源、類病變表型的形成機制及類病斑基因的研究進展,對深入研究植物類病變突變體對作物抗病的分子機理研究和栽培品種的遺傳改良都具有重要的意義。因此,類病變突變體的研究也能夠為具有較寬的抗譜、抗性持久的抗病基因提供豐富的種質資源。
1 類病變變異的來源
類病變突變體主要來源有2種,即自然變異和人工誘變。
1.1 自然變異
自然變異頻率很低,僅為10-8~10-5,此類突變體占比較小。1965年,SEKIGUCHI等發現了水稻第一個自然突變的類病變突變體sl(sekiguchi lesion),該突變性狀受1對隱性基因控制。隨后,bl1、bl2和spl1等水稻自然類病變突變體被相繼發現[6]。
1.2 人工誘變
其主要包括物理、化學和生物誘變等。物理誘變是指利用各種高能射線進行照射處理植物種子進行人工誘發變異,如水稻突變體lmi、spl3、spl5、rls1等[7]。化學誘變是利用化學誘變劑(如烷化劑、堿基類似物、抗生素等)處理植物組織,最常用的誘變劑是EMS。如lsd1、acd1、lmm1、spl11、spl29和les23等都是通過EMS誘導獲得的類病變突變體[7-8]。生物誘變主要包括T-DNA插入[9]、轉座子插入[6]等發生的變異,導致植株出現病斑表型。此外,在植物組織培養過程中體細胞再生也會導致類病變的發生。
2 類病變突變體的特征
類病變主要發生部位是葉片或葉鞘,其病斑的顏色和性狀不同,病斑出現的時期也有差異,多數出現在苗期也就是播種后2~3周(如水稻類病變突變體spl1~spl4)。但也有些突變體的斑點直到孕穗期才出現,如水稻類病變突變體sp16。根據病斑的擴展方式,類病變突變體也可分為起始型和擴散型2類[10]。起始型突變體斑點是一種局部自發性的細胞壞死現象,不向周邊蔓延擴散,如blm和M1009。擴散型突變體的病斑會擴散到整個葉片及植株,如spl1和lmm1[7]。
3 類病變突變體的遺傳模式
大多數突變體為單個基因控制,主要受1~2個基因控制,在玉米中多數為顯性突變,定位玉米斑點突變的基因分布在各個染色體上。小麥中已報道的類病變突變體有7個,除了突變體C591(M8)受顯性基因控制外,其余均為隱性遺傳。迄今報道的200多例水稻斑點突變,已經對80多份類病變突變體材料進行了遺傳分析和鑒定,類病變性狀主要受1~2個核基因控制,在這些突變體中有73個受單隱性基因控制。
4 類病變基因的克隆與功能分析
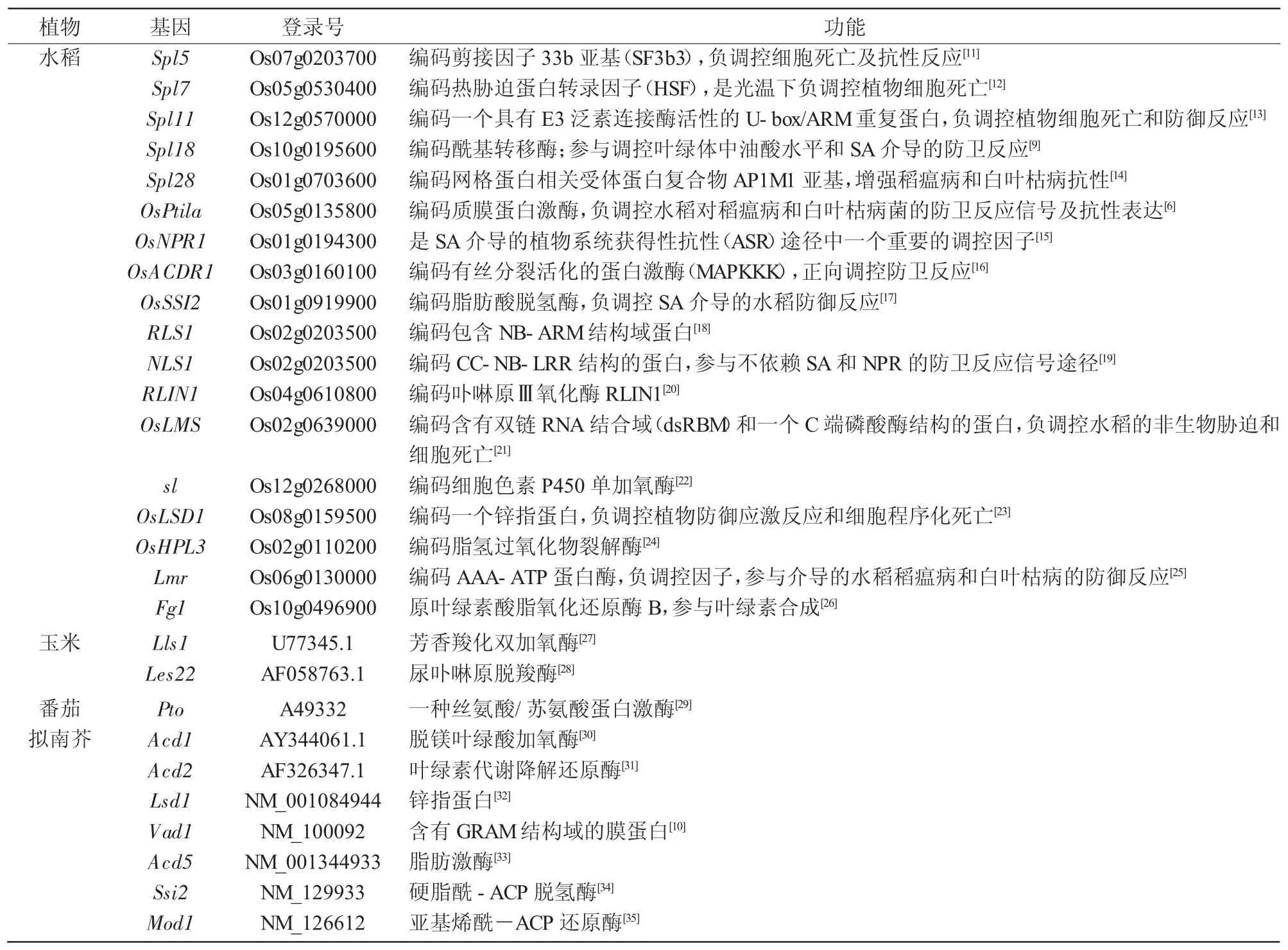
表1 植物類病變基因
自第1例類病變突變體報道以來,研究人員利用各種分子遺傳學技術,在擬南芥、玉米、大麥、水稻、番茄等的類病變突變體中克隆到多個控制類病變性狀的基因(表1)。
5 類病變突變體表型的形成機制
5.1 抗病基因的改變或功能缺失
植物抗病基因的改變啟動了過敏反應,導致植物細胞程序性死亡提前發生,誘發植物發生類病斑的癥狀。大麥抗白粉病基因Mlo的堿基突變致使其突變體自發形成壞死斑[20]。水稻NLS1基因、玉米RP1基因及番茄Pto基因的突變,會引發過敏性反應和程序性細胞死亡,形成壞死病斑。
5.2 正常代謝途徑紊亂
植物正常代謝紊亂會使植物PCD途徑受到干擾,有可能引起類病變表型的發生。主要涉及脂肪酸、氨基酸以及卟啉代謝途徑等。玉米類病變les22基因突變顯著降低尿卟啉原脫羧酶活性,誘導植株產生病變表型[5]。水稻中nls1基因突變伴隨防御反應組成性表達,其突變體中活性氧大量積累,導致類病變表型的發生[28]。水稻rlin1突變體由于編碼糞卟啉原III氧化酶的基因發生單堿基突變致使該酶活性降低,影響糞卟啉原Ⅲ氧化形成原卟啉Ⅸ的生化反應過程,導致植株出現類病變表型[21]。擬南芥SSCD1基因突變后絡氨酸降解異常,植株出現病斑表型[31]。MOU等[35]研究發現,編碼擬南芥脂肪酸合成的關鍵酶(烯酰-ACP還原酶enoyl-ACP reductase,ENR)的mod1基因發生突變后,烯酰基酰基載體蛋白還原酶的活性顯著降低,阻礙脂肪酸的合成,植株表現出類病變表型。水稻中多不飽和脂肪酸合成途徑中的關鍵酶OsSSI2基因突變導致類病變的產生[17]。
5.3 活性氧在植物體內的積累
類病變突變體壞死病斑的產生往往伴隨著活性氧物質的積累,這些富集的ROS可以直接破壞正常細胞的結構,從而導致類病變的產生。玉米les22突變體的葉片細胞壞死就是由活性氧積累直接造成的[28]。擬南芥突變體rcd1中過氧化物(O2-)和臭氧分子(O3)的過量積累導致產生壞死病斑[36-37]。水稻noe1突變體中編碼基因改變后,葉片中H2O2過量積累,最終誘導突變體植株產生類病變的表型[38]。
5.4 植株體內激素失調
激素信號在類病變表型發生的過程中起著重要的作用,主要是乙烯、水楊酸和茉莉酸,這幾種激素代謝失調均有可能導致類病變的發生。水稻OsSSI2基因的突變導致內源水楊酸含量和防御因子WRKY45表達量的上調[17]。水稻中敲除OsEDR1基因影響乙烯的合成,破壞體內的激素平衡,從而誘發產生類病變壞死病斑[16]。擬南芥突變體hrl1中乙烯、水楊酸過量也會導致病變表型的產生。上述結果暗示,植物激素可能參與植物類病變表型的發生。
5.5 生長環境因素
生長條件同樣可以誘導植物類病變表型的產生。玉米類病變突變體les22的類病變表型受光照的誘導[28]。劉道峰等[39]用錫紙遮光處理lmi突變體葉片,發現遮光部位無壞死斑點形成,表明lmi突變體細胞受光照誘導。MATEO等[40]構建了lsd1cao雙突變體探討病斑形成與光系統II的關系,推測lsd1類病變的表達與光合系統II有關。王建軍等[41]對6個水稻lrd類病變突變體進行不同溫度梯度處理,發現低溫明顯促進突變體lrd35和lrd40類病變發生,突變體lrd31在中溫下出現壞死斑。JAMBUNATHAN等[42]研究發現,擬南芥cpnl突變體屬于濕度敏感型突變體,通過調控Ca2+流向負向調控細胞死亡,產生類病變表型。
6 類病變突變體的抗病性研究
目前,已鑒定的很多類病變突變體或多或少都表現出增強的抗病性,不同的突變體抗病性存在較大的差異[42]。水稻突變體spl1、spl5、spl9和spl11表現出稻瘟病抗性增強,spl21、lmes1、hm197等12個突變體為白葉枯病抗性增強,突變體lmm1則是紋枯病抗性增強。這些突變體的抗病譜狹窄,其在育種上的應用并不多。一些具有廣譜抗性的突變體有非常高的實際應用價值,例如,在大麥中發現的類病變突變體mlo,對白粉病具有廣譜性抗性,在歐洲許多國家已經大面積種植。水稻中也有具有廣譜抗性的類病變突變體如spl11、lrd40、spl5等,為水稻抗病育種提供了非常重要的材料來源。
7 展望
類病變突變體是研究植物對病原菌的防御機制和程序性細胞死亡的理想材料。通過對類病變突變體研究,發現類病變的研究不僅涉及程序性細胞死亡、抗病抗逆生理以及HR反應產生的活性氧與自由基等方面,還與光信號及溫度誘導信號等有著密切的聯系。植物對病原菌的廣譜抗性通常在其類病變突變體中表現出來,因此,類病斑突變體是植物育種中重要的抗病種質資源。然而,部分類病變突變體還伴隨著株高降低、產量下降、分蘗能力減弱、早衰、雄性不育等農藝性狀的改變[42-43],使得類病變突變體中表現出來的廣譜抗性直接應用到農業生產中存在困難。因此,發掘對農藝性狀影響小、并具有廣譜抗性的類病變突變體材料在抗病育種上具有重大意義。
猜你喜歡
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
