高原鼢鼠肝臟組織細胞周期相關基因的進化和表達
2020-02-27 03:04:32安志芳魏琳娜王志潔李蘇華徐波李永曉魏蓮魏登邦
四川動物 2020年1期
安志芳, 魏琳娜, 王志潔, 李蘇華, 徐波, 李永曉, 魏蓮, 魏登邦, *
(1.青海大學省部共建三江源生態與高原農牧業國家重點實驗室,西寧810016; 2. 青海大學醫學院,西寧810016; 3.青海大學畜牧獸醫科學院,西寧810016; 4. 青海大學生態環境工程學院,西寧810016)
細胞周期是細胞生命活動的基本過程,主要分為G1、S、G2和M期(翟中和等,2000)。細胞周期的調控是通過各個時期特異的細胞周期調控因子實現的,主要包括3大類:細胞周期蛋白(cyclin),包括CyclinA、CyclinB、CyclinD、CyclinE等;細胞周期蛋白依賴性激酶(cyclin-dependent kinases,CDKs),包括CDK2、CDK4、CDK6等;細胞周期依賴性激酶抑制因子(cyclin-dependent kinases inhibitor,CDKI),包括p21、p16、p19、p27等。Cyclin是細胞周期正調控因子,CDKI是細胞周期負調控因子,CDK的活性受Cyclin正向調節,受CDKI負向調節,Cyclin會與對應的CDK結合,形成Cyclin-CDK復合物,進而決定細胞周期進程(Coatsetal.,1996;Sherr & Roberts,1999;Balter & Vogel,2001)。
研究表明,低氧誘導細胞周期蛋白依賴性激酶抑制因子p16、p21、p27高表達,CyclinD1、CyclinE、CDK4、CDK2等低表達,進而引起細胞周期G1期阻滯(Krtolicaetal.,1998;Gardneretal.,2001;Godaetal.,2003;Cazzalinietal.,2010;Hubbietal.,2013)。細胞周期G1期阻滯是機體應對外界刺激的一種保護反應,可以提供充足的時間修復受損的DNA,從而避免突變基因遺傳給子代細胞,避免腫瘤的發生(Kastanetal.,1991;Cazzalinietal.,2010)。低氧誘導p53下游Gadd45α、14-3-3-σ等基因的高表達以及CyclinB1等的低表達,引起細胞周期G2期阻滯(Hermekingetal.,1997;Innocenteetal.,1999;Puccietal.,2000;Hammeretal.,2007)。細胞周期G2期阻滯可以保證細胞有絲分裂過程中遺傳物質分配的忠實性,保持基因組的穩定(Kastanetal.,1991;Cazzalinietal.,2010)。
地下鼠是一類終身生活在完全封閉的地下洞道中的嚙齒動物,其地下洞道嚴重缺氧(Nevo,1999,2011;Nevoetal.,2001)。大量研究表明,在長期的進化過程中,地下鼠形成了一系列適應低氧的策略(Arieli & Ar,1981;Fangetal.,2014;Shaoetal.,2015;Maliketal.,2016;Danial-Farranetal.,2017;Schmidtetal.,2017)。研究表明,地下鼠具有壽命長和抗腫瘤的特征,以色列鼴鼠Nannospalaxgalili和裸鼴鼠Heterocephalusglaber的壽命分別長達21年和30年,且在國外40多年的研究過程中,無論是野外采集的樣本還是實驗室養殖的樣本,沒有發現一例患有腫瘤的個體(Buffenstein & Jarvis,2002;Buffenstein,2008;Kimetal.,2011;Edreyetal.,2012;Tianetal.,2013)。研究發現,生活在濕潤低氧的玄武巖地下洞道中的以色列鼴鼠腎臟組織中的p21基因表達水平顯著高于生活在干旱高溫的白堊土壤洞道中的個體(Zhaoetal.,2016)。Miyawaki等(2015)的研究發現,裸鼴鼠會上調p16和p19的mRNA和蛋白水平。Fang等(2014)對以色列鼴鼠基因組和轉錄組的研究發現,低氧上調其腦組織中p21、p16和p19基因的表達,下調CyclinD1、CyclinG1、CyclinG2和CDK2等基因的表達。因此,地下鼠通過上調細胞周期依賴性激酶抑制因子的表達、下調細胞周期蛋白等的表達誘導細胞周期G1、G2期阻滯。
Kim等(2011)對裸鼴鼠基因組的研究發現,CyclinE1在第335號位點由丙氨酸(Ala)變異為纈氨酸(Val),該位點的變異可能與裸鼴鼠長壽抗腫瘤有關。Kim等(2011)和Miyawaki等(2015)的研究發現,裸鼴鼠中細胞周期因子p16和p19結構的變異對其功能的發揮起著重要作用。p53是一個腫瘤抑制因子,通過上調或下調其下游靶基因表達,從而調控細胞周期,進行DNA損傷修復(Agarwaletal.,1998;Vogelsteinetal.,2000)。研究發現,以色列鼴鼠中p53結構的變異對其功能的發揮有重要的作用,p53在DNA結合域172位由精氨酸(Arg)突變為賴氨酸(Lys),這種變異使得p21基因高表達,細胞周期G1期阻滯(Ashur-Fabianetal.,2004;Avivietal.,2007)。因此,地下鼠細胞周期的調控不僅與細胞周期相關基因的表達水平有關,而且與其結構的變異有關。
高原鼢鼠Myospalaxbaileyi是生活在青藏高原2 800~4 200 m地區的一種典型的地下鼠,其洞道內氧氣的含量較同地區大氣中的低20%(王祖望等,1979;樊乃昌,施銀柱,1982;劉仁華,1995)。目前,在低氧條件下,關于高原鼢鼠肝臟組織中細胞周期相關基因的表達水平,以及細胞周期因子是否在長期低氧的作用下為了適應低氧環境發生了結構上變異的研究鮮見報道。因此,本文應用生物信息學方法對p53下游細胞周期相關因子p21、CyclinD1、CDK6、CyclinE、CDK2、14-3-3-σ、Gadd45α、B99和CyclinB1進行了進化分析,并以SD大鼠Rattusnorvegicus為對照,研究了這些基因在不同海拔(3 300 m和2 260 m)條件下高原鼢鼠肝臟組織中的表達模式。
1 材料與方法
1.1 實驗動物
16只高原鼢鼠捕捉于青海省西寧市湟源區宗家溝(101°17′E,36°43′N,海拔3 300 m),隨機分為2組,每組8只。實驗動物處理方法同An等(2018)的前期研究。第1組為高海拔組(海拔3 300 m),該組為宗家溝地區(海拔3 300 m)捕捉的樣本;第2組為低海拔組(海拔2 260 m),將捕捉于宗家溝的高原鼢鼠置于西寧實驗室(海拔2 260 m)飼養8 d。
16只SD大鼠購買于甘肅省蘭州市[實驗動物生產許可證號:SCXK(甘)2018-0002,實驗動物使用許可證號:SYXK(甘)2018-0002],隨機分為 2組,每組8只。第1組為高海拔組(海拔3 300 m),將SD大鼠置于宗家溝地區(海拔3 300 m)飼養8 d;第2組為低海拔組(海拔2 260 m),將SD大鼠置于西寧實驗室(海拔2 260 m)飼養8 d。
所有實驗動物均用5%戊巴比妥鈉麻醉,采集肝臟組織樣品立即置于液氮中保存,采樣過程中所涉及處理動物的措施均按照《實驗動物管理條例(GB14923-2010)》執行。
1.2 細胞周期基因序列分析
1.2.1 序列獲取從NCBI數據庫下載以色列鼴鼠、大鼠、小鼠Musmusculus、橙腹草原田鼠Microtusochrogaster、金倉鼠Mesocricetusauratus、黑線倉鼠Cricetulusgriseus、突尼斯非洲跳鼠Jaculusjaculus、裸鼴鼠、達馬拉鼴鼠Fukomysdamarensis、豚鼠Caviaporcellus、毛絲鼠Chinchillalanigera、智利八齒鼠Octodondegus、多紋黃鼠Ictidomystridecemlineatus、北美鼠兔Ochotonaprinceps、野兔Oryctolaguscuniculus、家牛Bostaurus、牦牛Bosgrunniens、綿羊Ovisaries、山羊Caprahircus、人Homosapiens和黑猩猩Pantroglodytes的細胞周期相關基因p21、CyclinD1、CDK6、CyclinE、CDK2、14-3-3-δ、Gadd45α、B99和CyclinB1的堿基及氨基酸序列。高原鼢鼠和高原鼠兔Ochotonacurzoniae細胞周期相關基因的堿基序列利用Blast程序將實驗室前期已測得的三代轉錄組數據庫中全長非嵌合序列文件以及二代轉錄組數據庫中的Trinity文件,構建本地Blast數據庫,分別用以色列鼴鼠和北美鼠兔上述周期相關基因的編碼區序列作為query文件進行Blast比對篩選,使用DNASTAR中的Lastergene程序(Burland,2000)拼接篩選出的基因片段,最終獲得完整的編碼區堿基序列,使用MEGA 7.0(Kumaretal.,2016)將所有比對以及拼接篩選出的序列進行比對,挑選與以色列鼴鼠和北美鼠兔同源性最高的一段序列作為目標基因的編碼區序列,利用基因探索者軟件將編碼區序列翻譯成氨基酸序列。
1.2.2 同源性分析選擇與高原鼢鼠親緣關系較近的2種地下鼠(以色列鼴鼠和裸鼴鼠)、大鼠、小鼠和人的p21、CyclinD1、CDK6、CyclinE、CDK2、14-3-3-δ、Gadd45α、B99和CyclinB1基因編碼區序列和氨基酸序列,用DNAMAN 9.0和MEGA 7.0進行同源性分析。
1.2.3 物種樹構建方法從NCBI數據庫中檢索并下載1.2.1中包括高原鼢鼠和高原鼠兔在內的22個物種(嚙齒目Rodentia 14個物種、兔形目Lagomorpha 3個物種、鯨偶蹄目Cetartiodactyla 3個物種和靈長目Primates 2個物種)的線粒體DNA全基因組序列,利用貝葉斯算法的MrBayes 3.2(Huelsenbeck & Ronquist 2001;Ronquist & Huelsenbeck,2003)構建貝葉斯系統進化樹。采用PAUP 4.0(Swofford,2002)和Modeltest 2.3(Darribaetal.,2012)篩選最優模型,以赤池信息量準則(Alaike information criterion,AIC)(Bozdogan,1987;Brooks,1989)為標準進行最優模型的篩選與確定,采用馬爾科夫鏈蒙特卡羅(Markov chain Monte Carlo,MCMC)運算(Gamerman & Lopes,2006),以隨機樹為起始,當運行2條和4條(1條冷鏈和3條熱鏈)MCMC時,分歧頻率的標準差穩定到小于0.01為止。在貝葉斯系統發育樹構建的過程中,共進行106代的MCMC運算,設置每100代間隔進行一次抽樣,舍棄起始老化樣本數(burn-in)占總數的25%(Wangetal.,2016)。利用TreeGraph 2.7.1作圖(St?ver & Müller,2010)。
1.2.4 選擇壓力分析分別將22個物種的p21、CyclinD1、CDK6、CyclinE、CDK2、14-3-3-δ、Gadd45α、B99和CyclinB1基因編碼區序列用ClustalX 1.81進行比對,比對后的結果利用MEGA 7.0進行格式轉換,應用PAML 4.8中的CODEML程序(Zhangetal.,2005;Yang,2007),選用該程序中改進的基于最大似然法的分支位點模型(“test2”)對基因序列進行正向選擇分析。將高原鼢鼠設為前景支,其余支系設為背景支,先用model A檢測前景支中是否存在顯著的正選擇位點,檢測標準為貝葉斯經驗貝葉斯值大于0.95;再將控制文件(codeml.ctl)中的fix omega和omega值都設定為1,作為Null A進行第二次運算,提取2次運算得到的lnL值,分別記為lnL1和lnL0,計算其加倍差值2×ΔlnL。最后利用PAML 4.8中的Chi2程序,基于2×ΔlnL值計算模型的后驗概率P值(df=1),當P<0.05時,可認為此模型得到的結果較可靠。
1.2.5 趨同進化分析選用1.2.1中的22個物種,對p21、CyclinD1、CDK6、CyclinE、CDK2、14-3-3-δ、Gadd45α、B99和CyclinB1基因的氨基酸序列進行地下鼠趨同進化分析。利用DNAMAN 9.0進行序列比對,使用PAML 4.8中的CODEML程序(Yang,2007)對每組蛋白序列進行了祖先序列重建。用重建后祖先位點的后驗概率評估重建結果的準確性。由于祖先位點多態性會干擾后續分析,因此舍棄后驗概率小于0.9的位點。應用converg 2(Zhang & Kumar,1997)檢驗趨同進化位點的顯著性;應用MEGA 7.0中的ClustalW模式對22個物種的氨基酸序列進行比對,去掉所有空格,并按指定格式做出系統發育樹,然后輸入樹文件和序列文件并應用Jones-Taylor-Thornton 距離矩陣模型和泊松校正模型分別計算,參數使用默認值,舍棄P>0.05的結果(Zhang & Kumar,1997)。
1.2.6 變異位點對基因功能影響的評估由于SIFT數據中與高原鼢鼠親緣關系最接近的物種為小鼠,因此從NCBI和Ensembl數據庫中下載得到小鼠p21、CyclinD1、CDK6、CyclinE、CDK2、14-3-3-δ、Gadd45α、B99和CyclinB1的蛋白ID,以該氨基酸序列作為query序列,采用“Sorting Tolerant From Intolerant(SIFT)”的algorithm程序評估氨基酸變異位點對該基因功能的影響,其中參數的設置使用默認值(Kumaretal.,2009)。
1.3 高原鼢鼠細胞周期基因表達水平測定
利用總RNA抽提試劑盒(天根,中國)提取高原鼢鼠和SD大鼠肝臟組織總RNA,核酸蛋白含量檢測儀測定A260/A280值(1.8
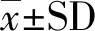

表1 熒光定量引物序列Table 1 Quantitative PCR primers used in this study
2 結果
2.1 高原鼢鼠細胞周期基因同源性比對結果
同源性比對發現,高原鼢鼠細胞周期相關基因p21、CyclinD1、CDK6、CyclinE、CDK2、14-3-3-σ、Gadd45α、B99和CyclinB1編碼區及氨基酸序列與以色列鼴鼠同源性最高,達到90%以上(表2)。
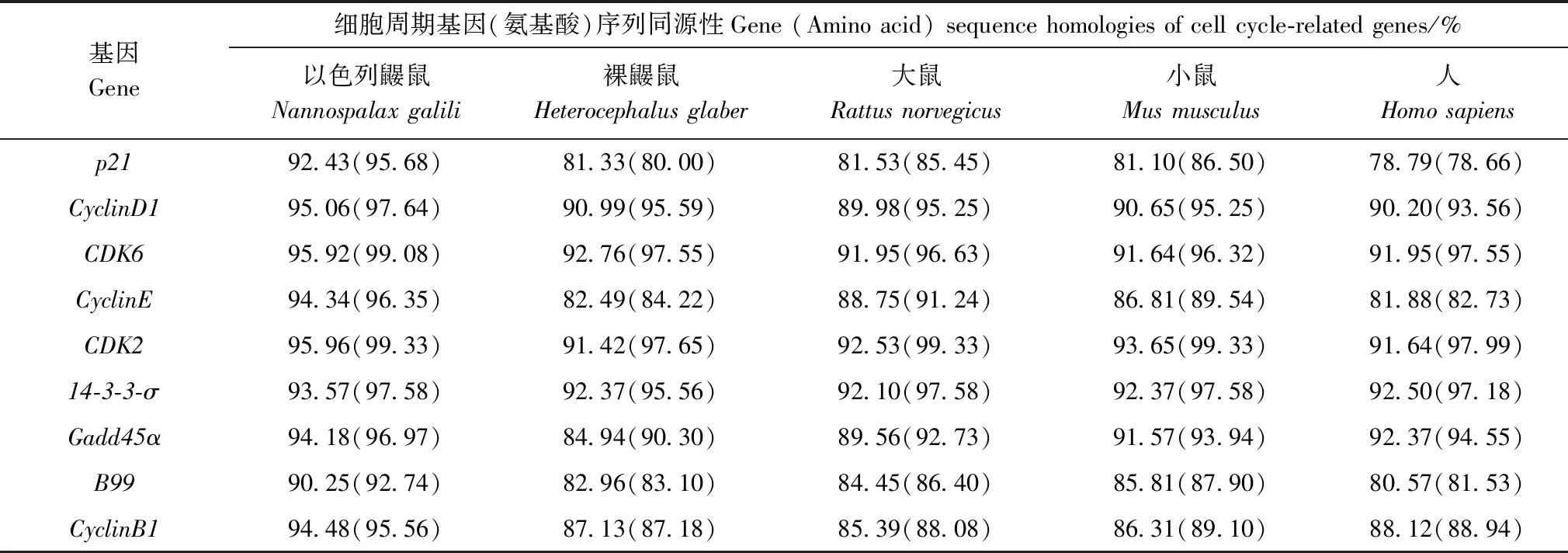
表2 高原鼢鼠與其他物種細胞周期基因、氨基酸序列同源性比較Table 2 Gene, amino acid sequence homologies of cell cycle-related genes between Myospalax baileyi and other species
2.2 物種樹的構建
根據選取的22個物種的線粒體DNA全基因組序列構建物種進化樹。DAMBE飽和度檢測結果顯示,指標分數(index score,ISS)低于臨界分數(critical score,TSS.C)(ISS=0.685,TSS.C=0.830,P<0.01),說明核酸替換未達到飽和,適合建樹。最佳DNA進化替代模型采用with gamma-distributed rate variation across sites和a proportion of invariable sites的GTR模型。所得的貝葉斯樹各支的支持率都大于85%(圖1),可以用于后續研究。

圖1 22種哺乳動物的mtDNA系統進化樹Fig. 1 Phylogenetic tree of 22 mammal species based on mtDNA
2.3 高原鼢鼠細胞周期相關因子選擇壓力分析
基于圖1構建的22個物種進化樹,檢測高原鼢鼠細胞周期相關因子p21、CyclinD1、CDK6、CyclinE、CDK2、14-3-3-σ、Gadd45α、B99和CyclinB1是否具有正向選擇位點。結果表明,CDK2有2個潛在的正向選擇位點,分別為第330號位點的蘇氨酸(Thr)和第341號位點的異亮氨酸(Ile);B99有2個潛在的正向選擇位點,為第168號位點的谷氨酰胺(Gln)和第447號位點的絲氨酸(Ser);Gadd45α有2個潛在的正向選擇位點,分別為第8號位點的絲氨酸(Ser)和第14號位點的賴氨酸(Lys);CyclinB1有1個潛在的正向選擇位點,為第396號位點的精氨酸(Arg),但是似然比檢驗法顯示這些位點差異不顯著(2ΔlnL=0,P>0.05)(表3)。
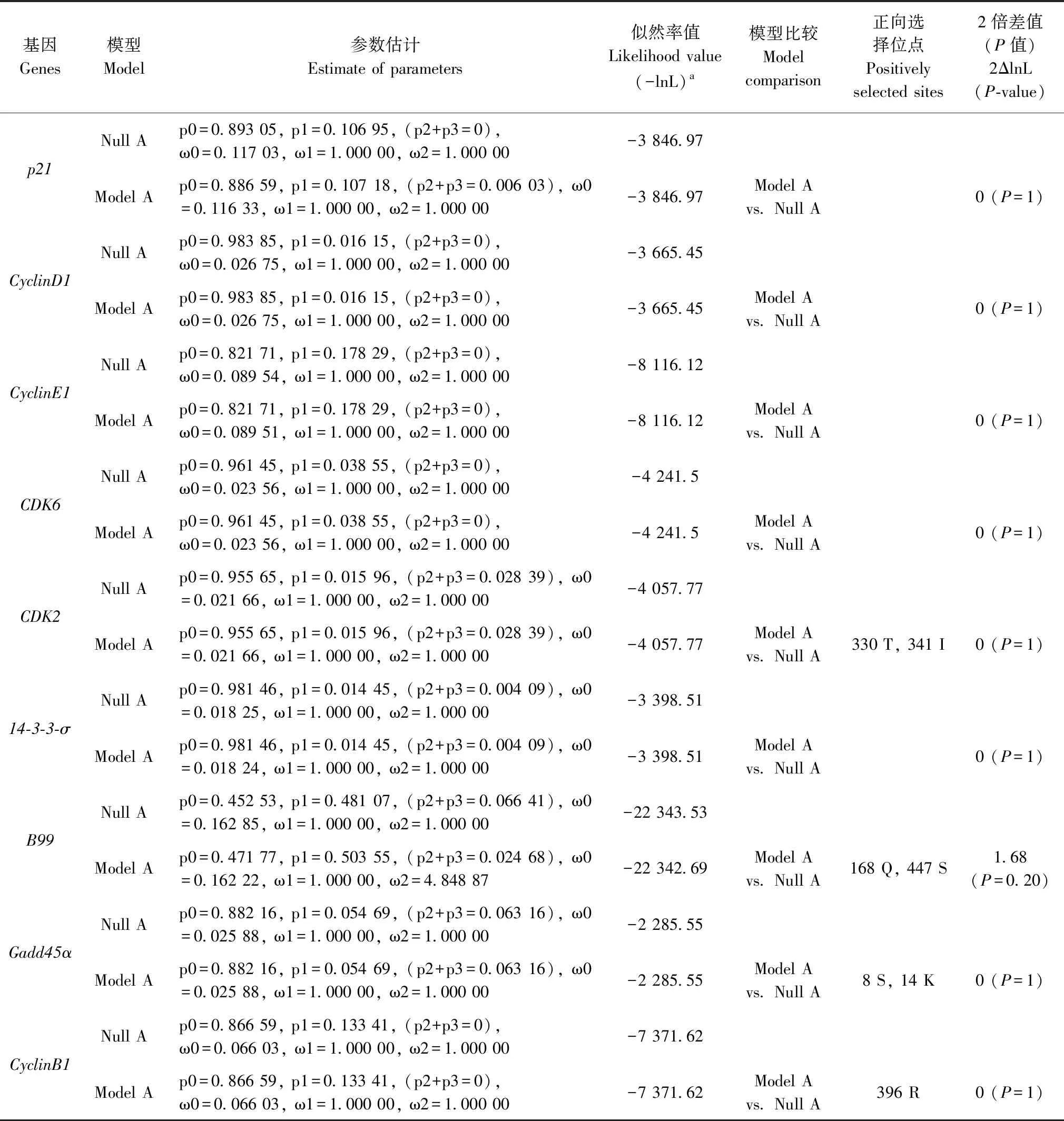
表3 高原鼢鼠細胞周期基因選擇壓力似然比檢驗Table 3 Likelihood ratio test of branch-site models for cell cycle-related genes in Myospalax baileyi
2.4 細胞周期相關因子趨同進化分析
22個物種細胞周期相關因子p21、CyclinD1、CDK6、CyclinE、CDK2、14-3-3-σ、Gadd45α、B99和CyclinB1(由于NCBI中無牦牛p21和CyclinB1基因序列,分別選用家牛對應序列:NP_001092428.1、NP_001039337.1)在地下鼠中的趨同進化分析顯示,高原鼢鼠和以色列鼴鼠p21在第27號位點由谷氨酸(Glu,E)變異為丙氨酸(Ala,A)(圖2:A);CyclinD1第20、24、143和266號位點分別由蘇氨酸(Thr,T)、天冬酰胺(Asn,N)、亮氨酸(Leu,L)和亮氨酸(Leu,L)變異為絲氨酸(Ser,S)、蘇氨酸(Thr,T)、異亮氨酸(Ile,I)和谷氨酰胺(Gln,Q)(圖2:B);CyclinE第86、92、125、126和402號位點分別由亮氨酸(Leu,L)、蘇氨酸(Thr,T)、谷氨酸(Glu,E)、賴氨酸(Lys,K)和賴氨酸(Lys,K)變異為纈氨酸(Val,V)、賴氨酸(Lys,K)、天冬氨酸(Asp,D)、天冬酰胺(Asn,N)和谷氨酰胺(Gln,Q)(圖2:C);CyclinB1第105、135和370號位點分別由纈氨酸(Val,V)、谷氨酸(Glu,E)和絲氨酸(Ser,S)變異為天冬酰胺(Asn,N)、天冬氨酸(Asp,D)和半胱氨酸(Cys,C)(圖2:D),而CDK6、CDK2、14-3-3-σ、Gadd45α和B99在高原鼢鼠和以色列鼴鼠中沒有趨同進化位點。

圖2 最大似然法構建的p21(A)、CyclinD1(B)、CyclinE(C)和CyclinB1(D)系統發育樹和進化位點Fig. 2 Maximum-likelihood tree of the p21(A), CyclinD1(B), CyclinE(C) and CyclinB1(D) gene sequences and the convergent sites
系統發育樹后的序列分別表示氨基酸和對應的堿基序列, 加粗及下劃線的氨基酸表示高原鼢鼠和以色列鼴鼠共有的趨同進化位點
Amino acids and codons of convergent sites are shown in the right part of each panel, and amino acids inMyospalaxbaileyiandNannospalaxgaliliare highlighted in bold and underline
2.5 高原鼢鼠細胞周期相關因子變異位點對其功能影響的評估
SIFT評估結果表明,高原鼢鼠與以色列鼴鼠細胞周期相關因子的趨同進化位點中,只有p21第27號位點丙氨酸(Ala,A)和CyclinB1第105號位點天冬酰胺(Asn,N)的變異對其功能有顯著影響,其余變異位點對基因功能均沒有影響(表4)。
表4 高原鼢鼠細胞周期基因氨基酸序列中 突變位點對其功能的影響Table 4 The effects of mutation sites on the function of cell cycle-related genes in Myospalax baileyi
2.6 高原鼢鼠和SD大鼠細胞周期基因mRNA表達水平
熒光定量PCR結果表明,與低海拔條件(2 260 m)相比,在高海拔條件(3 300 m)下,高原鼢鼠肝臟組織中細胞周期基因p21的表達水平極顯著高于低海拔(2 260 m)下的表達水平(P<0.01),p21下游基因CyclinD1、CDK6、CyclinE和CDK2的表達水平顯著低于低海拔(2 260 m)下的表達水平(P<0.05),而在SD大鼠肝臟組織中上述基因的表達水平差異都沒有統計學意義(P>0.05)。高原鼢鼠和SD大鼠肝臟組織中細胞周期基因14-3-3-δ、Gadd45α、B99和CyclinB1表達水平間的差異無統計學意義(P>0.05)(圖3)。在不同海拔條件下,高原鼢鼠肝臟組織中細胞周期相關基因p21、CyclinD1、CDK6、CyclinE、CDK2、14-3-3-σ、Gadd45α、B99和CyclinB1的表達水平均極顯著高于SD大鼠(P<0.01)(圖3)。
3 討論
G1期是細胞周期的第一階段,在這時期細胞開始合成生長所需的RNA、蛋白質、糖類、脂質等,同時細胞體積顯著增大,屬于細胞的生長期(翟中和等,2000)。研究表明,低氧誘導細胞周期蛋白依賴性激酶抑制因子p16、p21、p27高表達,CyclinD1、CyclinE、CDK4、CDK2等低表達,進而引起細胞周期G1期阻滯(Krtolicaetal.,1998;Gardneretal.,2001;Godaetal.,2003;Cazzalinietal.,2010;Hubbietal.,2013)。細胞周期G1期阻滯是機體應對外界刺激的一種保護反應,可以提供充足的時間修復受損的DNA,從而避免突變基因遺傳給子代細胞,避免腫瘤的發生(Kastanetal.,1991;Cazzalinietal.,2010)。研究表明,p21基因的高表達是引起細胞周期G1期阻滯的直接原因(Cazzalinietal.,2010)。p21作為細胞周期負調控因子,在其N端第21~26位及第49~72位的氨基酸分別與細胞周期蛋白和細胞周期蛋白依賴性激酶結合,抑制Cyclin-CDK復合物的活性,進而引起視網膜母細胞瘤蛋白不能發生磷酸化,阻止轉錄因子E2F的釋放,使得參與細胞周期的CyclinD1、CyclinE等不能表達,阻止細胞從G1期進入S期,導致細胞周期G1期阻滯(Sherr & Roberts,1995,1999;Gartel & Radhakrishnan,2005)。本研究結果表明,在高原鼢鼠肝臟組織中,與G1期調控相關的基因p21表達水平隨海拔的升高顯著升高,p21下游基因CyclinD1、CyclinE、CDK6和CDK2的表達水平隨海拔的升高顯著降低,而在SD大鼠中沒有差異;在高原鼢鼠和SD大鼠肝臟組織中,與G2期調控相關的基因Gadd45α、B99、14-3-3-δ和CyclinB1的表達水平隨海拔改變不發生變化。因此,高原鼢鼠通過長期的低氧適應,在高海拔條件下,p21基因的高表達抑制了下游CyclinD1、CyclinE、CDK6和CDK2基因的表達,從而誘導細胞周期G1期阻滯,提供了充足的時間修復受損的DNA,保證了DNA復制的準確性。
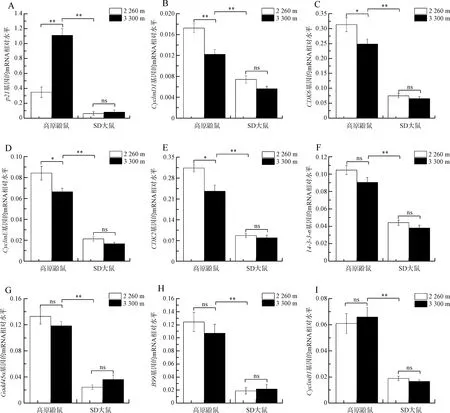
圖3 高原鼢鼠和SD大鼠肝臟組織中細胞周期基因在不同海拔條件下的mRNA表達水平
Fig. 3 Quantification of the mRNA levels of cell cycle-related genes in the liver ofMyospalaxbaileyiandRattusnorvegicusunder different altitudes
*P<0.05,**P<0.01, ns.P>0.05
細胞周期調控不僅與細胞周期相關基因的表達水平有關,而且與其結構的突變有關。p21是細胞周期負調控因子,主要有2個功能結構域,N端第21~26位及第49~72位的氨基酸分別與Cyclin和CDK結合,抑制Cyclin-CDK復合物的活性;C端第124~164位氨基酸與增殖細胞核抗原(proliferating cell nuclear antigen,PCNA)結合,使得PCNA不能與DNA聚合酶δ結合,抑制DNA的復制(Luoetal.,1995;Johnson & Walker,1999)。研究發現,p21第31號位點由極性不帶電的絲氨酸(Ser)變異為極性帶正電的精氨酸(Arg),這會使得p21蛋白結構發生變化,該位點的變異與乳腺癌、宮頸癌等的易感性有關(Lukasetal.,1997)。在人類乳腺癌中,p21第94號位點由極性帶正電的精氨酸(Arg)變異為非極性的色氨酸(Trp),該位點的變異使得p21抑制CDK的能力減弱,但是與PCNA結合的能力沒有發生改變(Balbínetal.,1996)。本研究的SIFT評估結果表明,高原鼢鼠p21第27號位點的變異對其功能有顯著影響,該位點由極性帶負電的谷氨酸(Glu)變異為非極性的丙氨酸(Ala),位于Cyclin結合域附近,其改變了該區域原本帶負電的局部環境,可能引起p21對Cyclin和CDK的結合力增強,從而使得p21阻滯細胞周期的功能增強。
CyclinB1屬于細胞周期蛋白,在其第201~288位有一個高度保守的細胞周期蛋白盒序列,該結構域與CDK1氨基末端的PSTAIR結構形成CyclinB1-CDK1復合物,該復合物被稱為有絲分裂促進因子;CyclinB1位于第42~50位的氨基酸序列為降解盒結構,參與細胞周期蛋白的自身降解(Glotzeretal.,1991;Nobleetal.,1997);此外,CyclinB1還有2個控制CyclinB1-CDK1復合物胞質-核穿梭的結構域,分別是位于第155~170位的核定位區域(nuclear localization signal,NLS)和N端第78~127位的胞質滯留結構域(cytoplasmic retention signal,CRS),NLS結構域的作用是將CyclinB1-CDK1復合物運送至細胞核內,CRS結構域是確保有絲分裂前CyclinB1-CDK1復合物在細胞質中的定位(Robbinsetal.,1991;Pines & Hunter,1994)。研究表明,位于CRS結構域中絲氨酸(Ser)磷酸化位點突變為丙氨酸(Ala)時,CyclinB1活性消失,而當突變為谷氨酸(Glu)時,可以增強其活性(Lietal.,1997)。SIFT評估結果表明,高原鼢鼠CyclinB1位于CRS結構域中的第105號位點的變異對其功能有顯著影響,該位點由非極性的纈氨酸(Val)變異為極性的天冬酰胺(Asn),這種變異可能會導致該點附近的靜電勢能發生變化,影響CyclinB1-CDK1復合物從細胞核運往細胞質,從而增強細胞周期阻滯作用。
綜上所述,高原鼢鼠經過長期的低氧適應,通過上調p21基因的表達抑制下游CyclinD1、CyclinE、CDK6和CDK2基因的表達,導致細胞周期G1期阻滯,從而提供充足的時間修復受損的DNA,保證了DNA復制的準確性;同時高原鼢鼠肝臟組織中細胞周期的調控不僅與細胞周期基因的表達水平有關,而且可能與細胞周期因子p21的第27號位點和CyclinB1的第105號位點的變異有關。
