長鏈非編碼RNA OR3A4在非小細胞肺癌中的作用及其機制探討
2020-01-17 02:23:40李珊珊顧文超
浙江臨床醫學 2019年12期
李珊珊 顧文超
肺癌是全球最常見的惡性腫瘤,也是導致癌癥死亡的主要原因。據估計,2018 年全球新增病例為2093876 例,死亡人數為1761007 人[1]。肺癌的主要組織學亞型是非小細胞肺癌(NSCLC),主要包括腺癌和鱗狀細胞癌,約占肺癌的80%[2]。雖然近年來NSCLC 的臨床治療取得了越來越多的進展,但NSCLC患者的總體生存時間并未顯著提高。因此,揭示非小細胞肺癌發生、發展的潛在分子機制,以開發新的非小細胞肺癌治療策略具有重要意義。近年來,非編碼RNA(non-coding RNA,ncRNA)因其豐富的調控手段在基因轉錄后調節中的作用越來越受到重視,其中長鏈非編碼RNA(lncRNA),一類長度>200 個核苷酸、編碼能力有限的非編碼轉錄產物,逐漸引起了人們的關注。研究發現lncRNA 在多種人類腫瘤中異常表達,在生物過程發揮重要作用,包括生長、細胞分化、增殖、凋亡和侵襲,發揮促癌或者抑癌基因的重要作用[3]。一些lncRNA 被報道在NSCLC 中具有致癌或抑制腫瘤的作用[4-5]。lncRNA OR3A4 是新近發現的一種lncRNA,在乳腺癌和胃癌中表達上調,促進癌癥的發生發展[6]。然而,lncRNA OR3A4 在NSCLC 中的表達、作用及作用機制尚不清楚。本研究旨在探討RNA OR3A4 在NSCLC 中的作用和機制,報道如下。
1 材料和方法
1.1 臨床標本和倫理聲明 在患者知情同意的情況下,經浦東新區人民醫院倫理委員會按照國際標準批準,從浦東新區人民醫院獲得30 對NSCLC 組織及相鄰非癌性肺組織樣本。另外,10 例NSCLC 骨轉移組織也取自浦東新區人民醫院手術的NSCLC 患者。所有患者術前均未接受放療或化療。組織標本冷凍保存于- 80℃冷凍箱。
1.2 細胞培養和轉染 人NSCLC 細胞株(A549、SPC-A1、H1299 和SK-MES-1)和HEK293T 細 胞 購自中國科學院細胞庫(上海),PC-9 細胞和人支氣管上皮(HBE)細胞購自ATCC(馬納薩斯,弗吉尼亞州,美國)。用DMEM 或RPMI1640 培養基培養NSCLC 細胞株。使用Lipofectamine 2000 按照制造商的協議進行細胞轉染。
1.3 qRT-PCR 分析 使用TRIzol?試劑從組織或細胞中提取總RNA。采用PrimeScript?RT 試劑試劑盒(TaKaRa,大連,中國)合成互補DNA,采用SYBR預混料Ex Taq II(TaKaRa)進行qRT-PCR 分析。β-actin作為標準化的對照。采用2-ΔΔCT方法計算相對基因表達水平。
1.4 Western blot 實驗 用RIPA 緩沖液制備細胞裂解液。在10%凝膠上用SDS-PAGE 分離等量的蛋白質,并轉移至聚偏氟乙烯膜上,然后用5%脫脂牛奶封閉1h。在4℃下用一級抗體將細胞膜培養過夜。使用的主要抗體如下:KLF6(Proteintech,武漢,中國)和β-actin(Proteintech)。將二抗37℃孵育1h 后,用增強的化學發光系統檢測觀察條帶。
1.5 質粒構建和細胞轉染 針對(shOR3A4)和陰性對照(shCtrl)的特異性shRNA 寡核苷酸購自上海吉瑪制藥技術有限公司。將編碼OR3A4 的互補DNA 經PCR 擴增后插入pcDNA3.1(Invitrogen)中。細胞轉染采用Lipofectamine 2000 進行。
1.6 細胞增殖檢測 使用CCK-8 試劑盒評估細胞增殖能力。在96 孔板中每孔接種2000 個轉染細胞,在不同時間點(接種后1d、2d、3d 和4d)檢測細胞活力。在490nm 處用分光光度微平板閱讀器測定吸光度。平板克隆形成實驗:將轉染后的細胞接種于6 孔板(1×103/孔)中,連續培養10d。更換培養基1 次/3d。實驗結束時,用4%多聚甲醛固定細胞,結晶紫染色。在顯微鏡下計算細胞克隆數量。
1.7 細胞凋亡測定 采用Annexin V-Alexa Fluor 647/ PI 凋亡檢測試劑盒檢測細胞凋亡。轉染細胞用膠原酶消化,收集細胞,用annexin V-Alexa Fluor 647 和PI在室溫下染色15min。流式細胞儀檢測細胞凋亡率。
1.8 細胞遷移實驗 收集轉染細胞,以每孔1×105個細胞的密度接種至上腔室。下室加入含有10% FBS 的500μl 細胞培養基。12h 后,用棉簽取出細胞膜上表面的細胞。然后用甲醛固定下層細胞,用結晶紫染色30min。遷移細胞的數量在顯微鏡下計數。
1.9 細胞侵襲實驗 將各室加入稀釋后的基底基質凝膠37℃下聚合30min。轉染細胞以2×105個細胞/孔的密度接種于上腔室。下室加入含有10% FBS 的500μl 細胞培養基。這些細胞被允許進入下膜24h。隨后,用棉簽取出細胞膜上表面的細胞。然后用甲醛固定下層細胞,用結晶紫染色30min。在顯微鏡下計數細胞遷移數。
1.10 體內實驗 動物實驗是在動物研究機構委員會的批準下進行的,嚴格按照美國國立衛生研究院實驗室動物護理和使用指南中的建議。雌性athymic BALB/c 小鼠(5 周齡)購自河南實驗動物中心(鄭州,中國),在特定的無致病性條件下培養。將穩定表達shCtrl 或shOR3A4 的A549 細胞(1×106)或穩定表達Ctrl 或OR3A4 的H1299 細胞(1×106)皮下注射于裸鼠側翼兩側(n=6 只/組)。測量腫瘤體積1 次/5d,計算公式為體積=(長×寬×0.5)。25d 后小鼠被處死,并對腫瘤進行稱重。
1.11 統計學分析 采用GraphPad Prism(Version 5.0,GraphPad Software,Inc)軟件。計量資料以()表示,組間比較采用t 檢驗或方差分析(one-way ANOVA)。P<0.05 為差異有統計學意義。
2 結果
2.1 OR3A4 在NSCLC 細胞系和組織中表達上調 為了探討OR3A4 在NSCLC 中的作用,采用qRT-PCR檢測了30 對NSCLC 組織樣本及相鄰非癌性肺組織中OR3A4 的表達情況。OR3A4 在NSCLC 組織標本中的表達明顯高于鄰近非癌性肺組織標本(P<0.05)(見圖1A)。此外,作者收集了10 例NSCLC 骨轉移組織,并測定了OR3A4 的表達。結果顯示OR3A4 在骨轉移組織中的表達高于原發性NSCLC 組織(P<0.05)(見圖1B)。此外,與HBE 細胞比較,OR3A4 在NSCLC 細胞 株(A549、SPC-A1、PC-9、SK-MES-1 和H1299)中的表達較高(P<0.05)(見圖1C)。

圖1 OR3A4在NSCLC組織樣本和細胞系中上調。A:OR3A4在30對NSCLC和非癌性肺組織樣本中的表達的定量分析,與正常相比較,*P<0.05。B:OR3A4在30個原發性NSCLC組織和10個骨轉移組織樣本中的表達的定量分析,與原發性NSCLC比較,*P<0.05。C:定量OR3A4在NSCLC細胞系和正常HBE細胞中的表達,與HBE組比較,*P<0.05。
2.2 下調OR3A4 表達抑制NSCLC 細胞增殖、侵襲和遷移 為了研究OR3A4 在NSCLC 中的作用,使用shRNA 下調了OR3A4 在NSCLC 細胞中的表達。qRTPCR 驗證了基因沉默的有效性(見圖2A)。用CKK-8法和平板克隆形成實驗測定了A549 細胞的增殖能力。結果表明,下調OR3A4 表達可顯著抑制A549 細胞的增殖速率(見圖2B)。菌落形成實驗結果表明,與對照組比較,下調OR3A4 的A549 細胞形成的菌落明顯減少(見圖2C)。作者用流式細胞術測定了OR3A4的下調對A549 細胞凋亡的影響。如圖2D 所示,在A549 細胞中,OR3A4 敲低顯著增加了凋亡細胞的百分比。然后在體外評估了OR3A4 敲低對NSCLC 細胞轉移潛能的影響。Transwell 遷移實驗結果顯示,與對照組比較,OR3A4 敲低組的細胞遷移更少(見圖2E)。此外,基質凝膠侵入的結果實驗表明,OR3A4 敲低也抑制了A549 細胞的侵襲能力(見圖2F)。

圖2 OR3A4敲低抑制NSCLC細胞的增殖、侵襲和遷移。A:qRT-PCR分析轉染shCtrl 和shOR3A4的A549細胞中OR3A4的表達。B:采用CCK-8法測定shCtrl 和shOR3A4兩組的細胞增殖能力。C:檢測shCtrl 和shOR3A4兩組的細胞克隆形成能力。D:流式細胞術分析shCtrl和shOR3A4轉染的A549細胞的凋亡情況。E:采用Transwell遷移實驗檢測下調OR3A4表達后A549細胞的遷移能力。F:采用基質凝膠侵襲法對轉染shOR3A4的 A549細胞進行侵襲性檢測。與ShCtrl組比較,*P<0.05。
2.3 上調OR3A4 表達促進NSCLC 細胞增殖、侵襲和遷移 使用pcDNA3.0 上調了OR3A4 在NSCLC 細胞中的表達。qRT-PCR 驗證了基因沉默的有效性(見圖3A)。用CKK-8 法和平板克隆形成實驗測定了H1299細胞的增殖能力。結果表明,上調OR3A4 表達可明顯促進H1299 細胞的增殖速率(圖3B)。菌落形成實驗結果表明,與對照組比較,上調OR3A4 的H1299 細胞形成的菌落明顯增加(見圖3C)。作者用流式細胞術測定了OR3A4 的上調對H1299 細胞凋亡的影響。(如圖3D)所示,在H1299 細胞中,OR3A4 過表達顯著降低了凋亡細胞的百分比。然后在體外評估了OR3A4過表達對NSCLC 細胞轉移潛能的影響。Transwell 遷移實驗結果顯示,與對照組比較,OR3A4 過表達組的細胞遷移增加(見圖3E)。此外,基質凝膠侵入的結果實驗表明,OR3A4 過表達也促進了H1299 細胞的侵襲能力(見圖3F)。
2.4 下調OR3A4 表達抑制NSCLC 腫瘤生長,但上調其表達促進NSCLC 腫瘤生長 為進一步證實OR3A4在NSCLC 中的生物學作用,將穩定轉染shCtrl 或shOR3A4 的A549 細胞,或穩定轉染Ctrl 或OR3A4的H1299 細胞皮下注射到裸鼠體內。測量腫瘤體積1次/5d,并進行腫瘤注射25d 后稱重。結果表明,與shCtrl 組比較,OR3A4 的下調顯著抑制了體內腫瘤的生長,包括平均腫瘤體積(見圖4A)和體重(見圖4B);OR3A4 的上調明顯促進了體內腫瘤的生長,包括平均腫瘤體積(見圖4C)和體重(見圖4D)。

圖3 OR3A4過表達促進NSCLC細胞的增殖、侵襲和遷移。A:qRT-PCR分析轉染OR3A4過表達質粒以及空白質粒的H1299細胞中OR3A4的表達。B:采用CCK-8法測定Ctrl和OR3A4轉染的H1299細胞的細胞增殖能力。C:檢測Ctrl 和OR3A4兩組的細胞克隆形成能力。D:流式細胞術分析Ctrl和OR3A4轉染的H1299細胞的凋亡情況。E:采用Transwell遷移實驗檢測過表達OR3A4表達后H1299細胞的遷移能力。F:采用基質凝膠侵襲法對轉染OR3A4過表達質粒的 H1299細胞進行侵襲性檢測。與Ctrl組比較,*P<0.05。
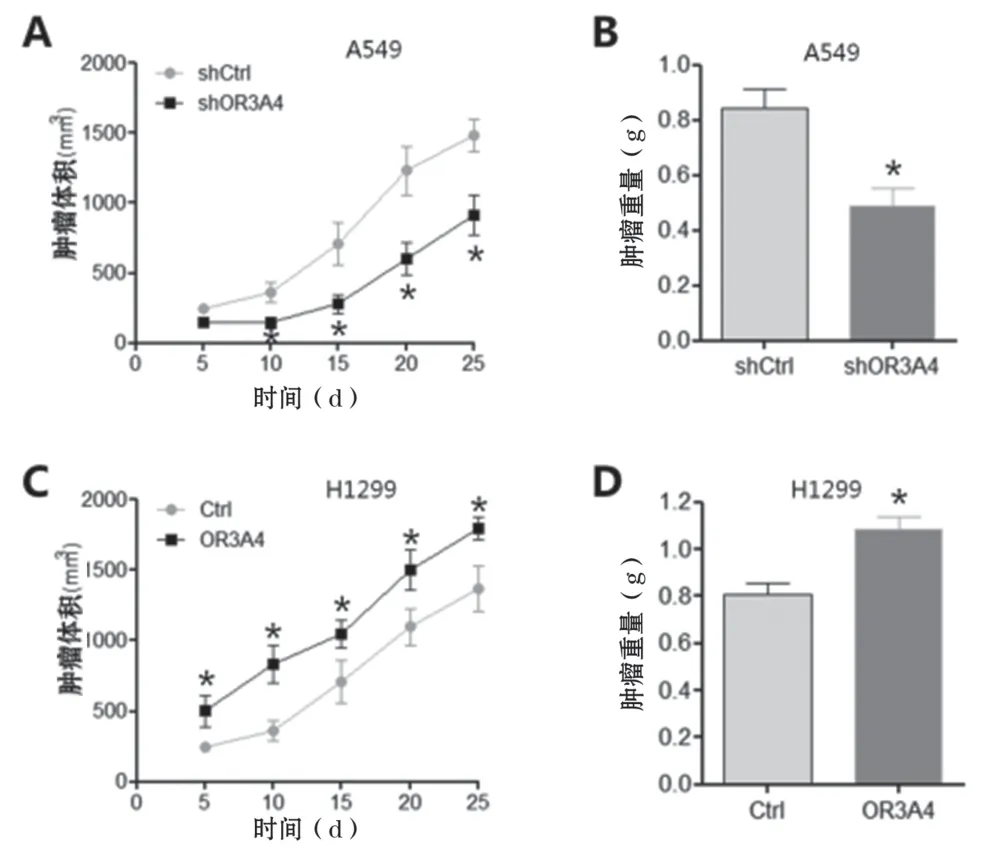
圖4 下調OR3A4抑制NSCLC腫瘤生長,上調OR3A4則促進NSCLC腫瘤生長。A:將穩定表達shCtrl或shOR3A4的A549細胞皮下注射于裸鼠,測量腫瘤體積1次/5d。B:腫瘤重量。C:將穩定表達Ctrl或OR3A4的H1299細胞皮下注射于裸鼠,測量腫瘤體積1次/5d。D:腫瘤重量。與Ctrl組比較,*P<0.05。
2.5 OR3A4 通過抑制KLF6 表達影響NSCLC 細胞生物學功能 已有報道OR3A4 通過抑制KLF6 促進卵巢癌的轉移[7]。KLF6 在NSCLC 中具有抑癌作用[8]。因此,作者首次研究OR3A4 是否也調控NSCLC 中KLF6的表達,以及OR3A4 在NSCLC 中的致癌作用是否依賴于KLF6 的調控。采用qRT-PCR 和western blot 檢測KLF6 在OR3A4 穩定過表達的H1299 細胞和OR3A4敲低的A549 細胞中的表達。結果顯示KLF6 過表達下調了PXN mRNA 和蛋白水平,而敲低OR3A4 表達后上調PXN mRNA 和蛋白水平。另外,在OR3A4 過表達的H1299 細胞中轉染KLF6 過表達載體明顯抑制了OR3A4 對細胞增殖、侵襲和遷移的促進作用,而在shOR3A4 敲低的H1299 細胞中轉染KLF6 敲低載體顯著抑制了OR3A4 對細胞增殖的抑制作用。提示OR3A4 在NSCLC 中的促癌作用至少部分依賴于KLF6的負向調控。
3 討論
在過去的十年中,各種lncRNA 被發現在癌癥的發病機制中發揮著重要的作用[9-10]。在本研究中,作者發現OR3A4 在NSCLC 組織和細胞株中表達上調。OR3A4 過表達促進NSCLC 細胞增殖、遷移和侵襲,而下調OR3A4 表達則抑制NSCLC 細胞增殖、遷移侵襲和腫瘤生長。另外,進一步證明了OR3A4 在NSCLC 中的作用可能通過抑制KLF6 表達。結果表明OR3A4 在NSCLC 中發揮致癌作用。
本資料數據表明,OR3A4 的表達上調可能有助于NSCLC 的進展,這在其他癌癥中也有類似的發現。Li等[11]證明OR3A4 通過調控AGGF1/akt/mTOR 通路參與肝細胞癌血管生成,證實OR3A4 是肝癌進展和血管生成的促進劑。在胃癌中,過表達OR3A4 促進胃癌細胞生長,血管生成和轉移,并且證明了OR3A4 通過調節PDLIM2、MACC1、NTN4、GNB2L1 的活化,影響胃癌細胞的生物學功能[6]。OR3A4 在卵巢癌組織中的表達水平明顯高于癌旁組織。此外,OR3A4 基因在體外敲除后,細胞遷移和侵襲受到明顯抑制[7]。此外,在乳腺癌中,OR3A4 在乳腺癌組織和細胞中表達上調,并與臨床病理特征相關。下調OR3A4 表達通過影響細胞周期和凋亡抑制乳腺癌細胞的增殖,并通過EMT 抑制細胞轉移,過表達OR3A4 取得相反實驗結果。結果表明,OR3A4 是一種潛在的診斷生物標志物和治療目標的各種癌癥。
大量研究表明,KLF6 可以作為腫瘤抑制因子,調控增殖、遷移、分化、凋亡、細胞周期和血管生成等多種基本細胞功能,從而抑制腫瘤的發生[12]。KLF6通過下調間質標志物和抑制MMP-9 活性,抑制口腔癌的遷移和侵襲。KLF6 通過轉錄抑制E2F1 抑制透明細胞腎細胞癌的轉移。Guo 等[7]指出OR3A4 通過抑制KLF6 促進卵巢癌的轉移。另外有研究表明,KLF6在NSCLC 中表達下調,并通過誘導NSCLC 細胞凋亡抑制腫瘤生長,這可能提示KLF6 是NSCLC 的抑癌因子[8]。本研究中,在NSCLC 細胞中過表達OR3A4 顯著降低KLF6 表達,而降低OR3A4 表達則明顯促進KLF6 表達。另外結果證明NSCLC 細胞中轉染KLF6可以顯著減輕OR3A4 對NSCLC 細胞增殖、侵襲和遷移能力的影響。說明OR3A4 可能通過抑制KLF6 表達促進NSCLC 細胞增殖、侵襲和遷移。
綜上所述,OR3A4 至少部分地通過KLF6 沉默促進NSCLC 細胞的增殖、遷移和侵襲。本研究為OR3A4 在NSCLC 進展中的作用提供了新的證據,并為NSCLC 的診斷和治療提供了一個潛在的標志物。
猜你喜歡
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
