荷花株型相關性狀與SSR分子標記的關聯分析
2020-01-04 07:14:15楊丹杜鳳鳳常雅軍崔鍵孫林鶴劉曉靜姚東瑞
熱帶作物學報 2020年11期
楊丹 杜鳳鳳 常雅軍 崔鍵 孫林鶴 劉曉靜 姚東瑞

摘 ?要:以129份株型多樣性豐富的蓮屬(Nelumbo Adans.)群體為供試群體,觀測與株型相關的立葉高度、花柄高度、立葉短徑、立葉長徑等數量性狀。供試荷花群體株型性狀變異豐富,立葉高度、花柄高度變異系數分別達39.68%和41.69%,立葉短徑、立葉長徑變異系數分別為29.70%和31.41%。選用多態性好的98對SSR引物對129份蓮屬種質進行多位點掃描。利用STRUCTURE V2.2軟件對供試蓮屬品種進行基于混合模型的群體結構分析。利用GLM模型分析與蓮株型性狀相關聯的SSR標記。結果表明:從群體結構看,供試群體最終可被劃分為4個亞群,中國蓮傳統品種和中美雜交種屬于不同的亞群,同時有部分遺傳背景復雜的雜交種在不同亞群間分布。通過關聯分析共找到11個標記與花柄高度、立葉高度、立葉長徑、立葉短徑等4個性狀顯著相關。其中,SSR067、SSR132、SSR458標記與所觀測株型性狀均顯著相關,可能由于基因連鎖或者一因多效引起。在連續2年的重復試驗中,SSR132和SSR458標記均可與立葉長徑、立葉短徑相關聯,表明這2個標記可能受環境影響較小。
關鍵詞:荷花;株型;群體結構;關聯分析
中圖分類號:S813.3 ? ? ?文獻標識碼:A
Association Studies of Plant Architecture Traits and SSR Markers in Nelumbo Adans.
YANG Dan, DU Fengfeng, CHANG Yajun, CUI Jian, SUN Linhe, LIU Xiaojing*, YAO Dongrui*
Institute of Botany, Jiangsu Province and Chinese Academy of Sciences, Nanjing, Jiangsu 210014, China
Abstract: Totally 129 lotus accessions with different architecture were used as the experimental material. The leaf height, flower height and leaf diameter were determined. The accessed lotus population had diverse plant architecture traits. Variation coefficient for leaf height and flower height was 39.68% and 41.69% respectively, and variation coefficient for leaf length and width was 29.70% and 31.41% respectively. 129 lotus accessions were genotyped with 98 pair polymorphic SSR primers. Population structure of lotus accessions was analyzed using the STRUCTURE V2.2 software. Association analysis of lotus architecture traits was performed with the GLM model. The results indicated that lotus accessions were divided into four subgroups. The results distinctly grouped lotus accessions from Chinese lotus and Sino-American hybrids into different clusters. However, the hybrids dispersed among clusters. The results showed that 11 SSR markers were associated with leaf height, flower height, leaf length and width; three markers were associated with all the plant architecture traits, and that might be caused by multiple effects of one gene or genetic linkage. In two years of repeated tests, two markers (SSR132 and SSR458) can be associated with leaf length and width, indicating that these two markers may be less affected by environment.
Keywords: Nelumbo; plant architecture; population structure; association analysis
DOI: 10.3969/j.issn.1000-2561.2020.11.002
荷花是蓮科(Nelumbonaceae)蓮屬(Nelumbo)多年生水生植物,是被子植物中起源最早的物種之一,在中國已有2500多年的栽培歷史[1]。荷花作為我國十大傳統名花之一,具有很高的觀賞價值,同時兼具食用、藥用價值,是我國最具特色、栽培面積最大的水生經濟植物。蓮品種繁多,目前已超過800余種[2]。
蓮屬目前僅存中國蓮(Nelumbo nucifera)和美洲蓮(Nelumbo lutea)。中國蓮和美洲蓮不存在生殖隔離,可以互相雜交[1]。美洲蓮花色和花型單一,中國蓮形態變異大,尤其在株型、花型、花色、花態等方面多樣性非常豐富,且品種數量遠多于美洲蓮,是培育蓮新品種、研究遺傳多樣性的重要資源。但由于荷花的很多觀賞性狀如株高、花朵大小等多表現為數量性狀,是由多個基因和環境共同作用的結果,對其遺傳基礎的研究比較困難。
近年來,研究者采用RAPD、AFLP、ISSR等分子標記方法,在評估野生荷花種質或荷花不同地理群體的遺傳多樣性和親緣關系方面做了相關工作[3-7]。由于上述研究的群體和對象存在較大差異,采用的分子標記技術也不同,使聚類情況較為復雜,部分研究并未檢測出豐富的遺傳多樣性[4-5],其研究結果有待進一步商榷。2013年,中國古代蓮全基因組序列的公布,為蓮功能基因組學、新基因挖掘提供了豐富的遺傳信息[8]。Yang等[9]從我國古代蓮的基因組序列中發現86 089個SSR motifs,測試開發了500對SSR引物,并利用這些新的引物構建了亞洲蓮和美洲蓮的遺傳連鎖圖譜。這些報道在一定程度上有助于了解和掌握蓮種質資源的遺傳背景。杜鳳鳳等[10]采用SSR標記技術對42個荷花品種進行了聚類分析和群體結構分析,明確42個荷花品種的遺傳多樣性特征。楊郭陽[11]利用F1遺傳群體構建了荷花遺傳連鎖圖譜,采用復合區間作圖法檢測了株型相關QTLs位點。迄今為止,以荷花自然群體或種質資源作為材料,研究荷花株型相關性狀的SSR關聯分析未見報道。
本研究以129份株型多樣性豐富的荷花群體為供試群體,利用多態性好的98對SSR引物,在分析遺傳多樣性和群體結構的基礎上,進行SSR標記與立葉高度、花柄高度、立葉直徑等株型相關性狀的關聯分析,旨在為荷花理想株型的構建及培育微型或大型荷花新品種提供理論依據。
1 ?材料與方法
1.1 ?材料
供試材料來源于江蘇省中國科學院植物研究所荷花種質資源庫,地理坐標為北緯32°04、東經118°45;年平均溫度為15.7 ℃,年降雨量為1106.5 mm,年平均降雨天數為117 d。供試的129份中國蓮種質名稱見表1。所有品種于2017年、2018年清明左右翻池,采用種藕繁殖,連續2年7—8月盛花期觀察記錄。
1.2 ?方法
1.2.1 ?表型數據的選取與測量 ?每個品種隨機選擇3株有代表性、無病蟲害、生長狀態良好的植株進行觀察測量,觀察記錄花柄高度、立葉高度、立葉直徑等重要性狀。各指標性狀具有不同的記錄標準,統計分析時對質量性狀予以賦值,對數量性狀進行分級分類。利用SPASS和Excel軟件進行統計分析。
1.2.2 ?基因組DNA的提取和SSR分子標記 ?采集供試樣品健康無病害的幼嫩葉片,利用組織/細胞基因組DNA提取試劑盒(Aidlab)提取各樣品的基因組DNA;然后采用1%瓊脂糖凝膠電泳和OneDrop分光光度計檢測DNA質量和濃度;再將其濃度稀釋至50 ng/μL,于?20 ℃保存備用。基于已報道[8-9]的荷花遺傳連鎖圖譜和基因組信息,分別從遺傳圖譜的不同染色體上,選用多態性好且已開發的荷花SSR引物140對,隨機選擇5份供試荷花DNA進行擴增,篩選出多態性好、條帶清晰的98對引物用于全部荷花品種基因組DNA樣品的擴增。采用ABI 3730XL對擴增產物進行毛細管電泳檢測,內參Liz 500(Ic?lo?ning)。采用GeneMarker軟件對下機數據進行panel的編寫及條帶的判讀。
1.2.3 ?SSR數據處理與軟件分析 ?采用POPGENE 1.31軟件計算位點數(Na)、有效位點數(Ne)、Shannon信息指數(I)、觀測雜合度(Ho)、Neis期望雜合度(He)[12]。利用PowerMarker V3.25軟件基于Neis遺傳距離構建UPGMA聚類圖[13]。利用STRUCTURE v2.2軟件分析基于混合模型的群體結構[14]。
采用TASSEL v2.1軟件度量SSR位點間的連鎖不平衡水平,計算其標準不平衡系數(D)和決定系數(r2),繪制LD配對檢測的矩陣圖[15-16]。在表型性狀及群體結構和連鎖不平衡水平分析的基礎上,再次利用TASSEL v2.1軟件[17]的GLM(General Linear Model)程序,以各個體的Q值作為協變量進行群體矯正,分別利用花柄高度、立葉高度、立葉直徑(長徑和短徑)等性狀數據對SSR分子標記變異進行回歸分析,估算標記位點等位變異表型效應。
2 ?結果與分析
2.1 ?株型相關性狀的統計分析
選擇種質圃株型差異較大的129份荷花品種進行性狀統計。與株型相關的數量性狀分布情況及多樣性統計結果見圖1,立葉高度變異范圍為7.5~141 cm,花柄高度變異范圍為18.5~160 cm,變異系數分別達39.68%和41.69%。立葉短徑、立葉長徑變異系數分別為29.70%和31.41%。由此可知,供試荷花群體株型的性狀變異豐富。
2.2 ?基于SSR擴增的遺傳多樣性分析
利用多態性好、條帶清晰的98對SSR引物對129份蓮屬種質進行擴增,擴增產物經毛細管電泳熒光檢測、測序獲得等位基因數據,進一步分析其遺傳多樣性參數。98對引物共擴增得到537個位點(即總多態性位點數為537),每對引物擴增多態性位點2~16個,平均每對引物擴增位點數為5.48個。有效位點數(Ne)最大達7.77(引物SSR065),最小為1.16(引物SSR400),平均值為2.58。Shannon信息指數(I)最大為2.32(引物SSR065),平均值為1.04;觀測雜合度(Ho)最大為1.00(引物SSR412、SSR240);Neis期望雜合度(He)最大為0.87(引物SSR065)。引物SSR078的觀測雜合度(Ho)和Neis期望雜合度(He)均較大,分別為0.99和0.83。引物SSR463的Ho和He也均較大,分別為0.99和0.83(表2)。
2.3 ?UPGMA聚類分析
根據SSR分析數據計算各品種間的Neis遺傳距離,并進行UPGMA聚類分析,結果見圖2。其中‘姬妃蓮(119)、‘嵊縣粉蓮(117)、‘點額妝(126)、‘平頭紫(121)、‘普者黑白荷(81)等傳統品種最先被分類(圖2中的藍色部分),近年引入美洲蓮基因后培育出的雜交蓮品種‘六朝玉葉(80)、‘江南煙雨(78)、‘珠峰翠影(79)、‘秦淮翡翠(75)、‘洋貴人(76)、‘俊愉蓮13B(33)、‘金陵女神(83)、‘金陵暢想(96)、‘江南碧菡(59)、‘琴韻(70)、‘大師(77)等聚在一起(圖2中的紅色部分)。聚類結果出現了很多不同的亞群,且未發現與傳統形態學分類直接相關。分析其原因,一是蓮屬大多數品種是通過自然雜交和對實生苗連續多年的人工選育而來,其遺傳背景復雜、親本信息不明;二是在長期的歷史演化和人為育種選擇的過程中,中國蓮群體演化出了多個亞群。
2.4 ?群體遺傳結構分析
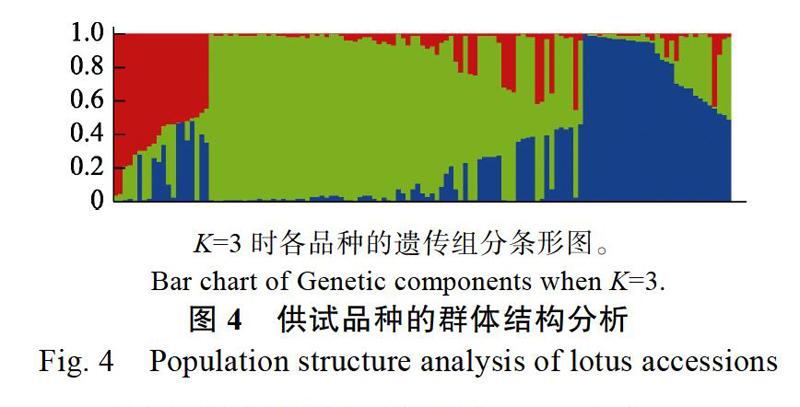
利用STRUCTURE V2.2軟件對供試蓮屬品種進行基于混合模型的群體結構分析。設定推測亞群數K=1~10,5個重復,根據運行結果計算LnP(D)及ΔK值[18],分別繪制LnP(D)和ΔK隨K值變化的散點圖。如圖3所示,LnP(D)隨K值的增大緩慢增大,并在K=10時有所降低,ΔK值在K=3時達到最大值。如圖4所示,當K=3時,即推定供試群體具有3種遺傳組分,設定閾值為0.60,供試群體被分成4個亞群,分別為P1、P2、P3及混合群M1。P1亞群共包括10個品種,其中包括‘大師‘伯里王子等雜交蓮品種。P2亞群共67個品種。P3亞群共包括25個品種,其中包括‘平頭紫‘姬妃蓮‘解放紅蓮等傳統品種。剩余27個品種被分為混合群M1。
2.5 ?蓮屬重要性狀相關聯的SSR標記
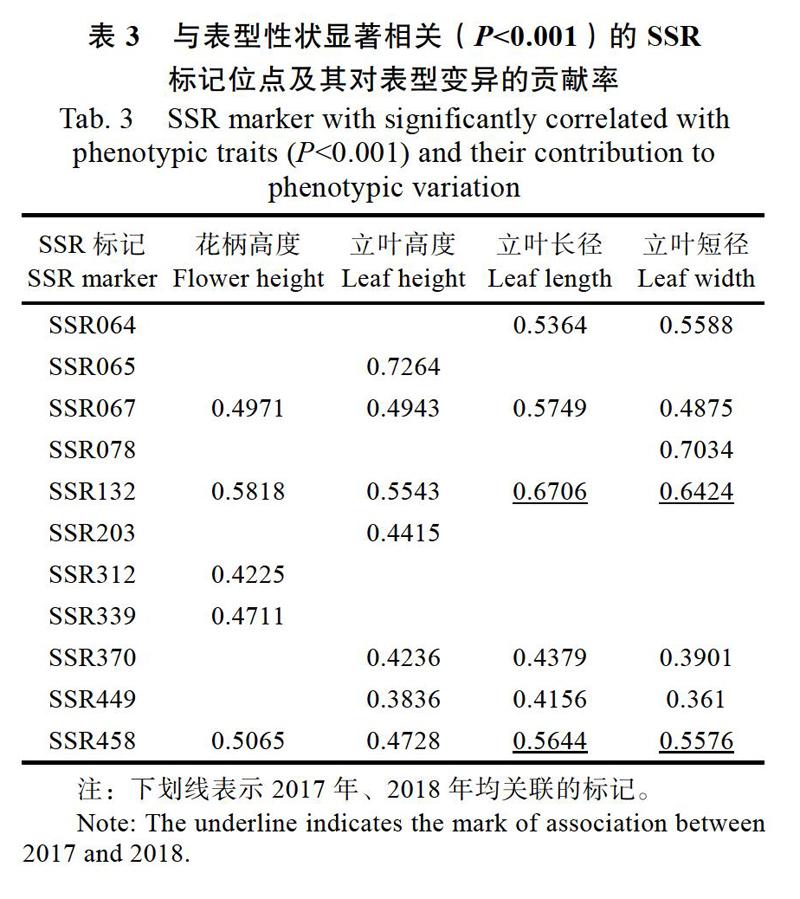
以供試的129份種質對應的Q值為協變量,利用GLM模型分析與蓮表型性狀相關聯的SSR標記位點,結果顯示共找到11個標記與花柄高度、立葉高度、立葉長徑、立葉短徑等4個性狀顯著相關(P<0.001),其中SSR067和SSR132分別與株型相關的4個性狀均顯著關聯。有5個標記與花柄高度關聯,其中表型變異貢獻率最大的是SSR132(0.5818);有7個標記與立葉高度關聯,其中表型變異貢獻率最大的是SSR065(0.7264);有6個標記與立葉直徑(長徑和短徑)同時關聯,其中表型變異貢獻率最大的是SSR132(0.6706和0.6424)。分析結果發現,同一標記可與多個性狀相關聯,如SSR067、SSR132、SSR458標記均與花柄高度、立葉高度、立葉長徑、立葉短徑等性狀相關聯,可能由于基因連鎖或者一因多效引起的。在連續2年的重復試驗中,SSR132和SSR458標記均可與立葉長徑、立葉短徑相關聯,這2個位點受環境影響可能較小(表3)。
3 ?討論
群體遺傳結構的解析對于種質資源的利用非常重要。迄今為止,很多園藝植物的群體結構已有相關報道[19-21]。在本研究中,UPGMA聚類分析表明供試蓮群體內部存在不同亞群,其遺傳背景較為復雜,STRUCTURE軟件群體結構分析進一步證實了這一結果。群體結構分析可較好的區分傳統品種與亞美雜交蓮品種。在最大的亞群P2中,其遺傳組分較為復雜,可能由于地理分割或者歷史演化在群體內部產生了多個亞群。此外,有27個品種被分為混合群M1,可能與長期雜交育種過程中的基因漸滲有關[10]。
荷花的株型、花期等性狀易受環境影響,與氣候和栽培條件均有較大關系,這些性狀的復雜性容易影響不同年份的關聯分析結果。Yang等[22]利用AFLP、SRAP和SSR標記挖掘荷花花朵觀賞性狀和花期相關聯的位點,結果發現14個標記可以和連續兩年的花瓣數相關聯,而對于始花期和花期性狀來講,很多標記在兩年的數據觀測中,都僅有一年與目標性狀顯著關聯,推測花期性狀可能受環境影響較大。在本研究中,一共找到11個標記與荷花花柄高度、立葉高度、立葉長徑、立葉短徑等4個性狀顯著相關。有2個標記可以連續兩年與立葉長徑、立葉短徑相關聯,而花柄高度、立葉高度這2個性狀均只有一年可以與標記顯著關聯。這一結果可能由于氣候或環境因素造成不同年份關聯分析結果的差異。雖然以上發現的株型相關聯位點在一定程度上受環境因素的影響,但這些位點的挖掘為蓮的株型改良提供了依據,在今后的研究中,可采用高密度遺傳圖譜對這些重要位點進一步驗證。
參考文獻
[1] 王其超, 張行言. 中國荷花品種圖志: 續志[M]. 北京: 中國建筑工業出版社, 1999.
[2] 張行言, 陳龍清, 王其超. 中國荷花新品種圖志[M]. 北京:中國林業出版社, 2011.
[3] Fu J, Xiang Q, Zeng X B, et al. Assessment of the genetic diversity and population structure of lotus cultivars grown in China by amplified fragment length polymorphism[J]. Journal of American Society Horticultural Science, 2011, 136: 339-349.
[4] Chen Y Y, Zhou R C, Lin X C, et al. ISSR analysis of genetic diversity in sacred lotus cultivars[J]. Aquatic Botany, 2008, 89(3): 311-316.
[5] Hu J H, Pan L, Liu H G, et al. Comparative analysis of genetic diversity in sacred lotus (Nelumbo nucifera Gaertn.) using AFLP and SSR markers[J]. Molecular Biology Reports, 2012, 39(4): 3637-3647.
[6] Li Z, Liu X Q, Gituru R W, et al. Genetic diversity and classification of Nelumbo germplasm of different ori-gins by RAPD and ISSR analysis[J]. Scientia Horticulturae, 2010, 125(4): 724-732.
[7] Yang M, Han Y N, Xu L M, et al. Comparative analysis of genetic diversity of lotus (Nelumbo) using SSR and SRAP markers[J]. Scientia Horticulturae, 2012, 142: 185- 195.
[8] Ming R, VanBuren R, Liu Y L, et al. Genome of the long-living sacred lotus (Nelumbo nucifera Gaertn.)[J]. Genome Biology, 2013, 14(5): 131-143.
[9] Yang M, Han Y N, VanBuren R, et al. Genetic linkage maps for Asian and American lotus constructed using novel SSR markers derived from the genome of sequenced cultivar[J]. BMC Genomics, 2012, 13(1): 1471-2164.
[10] 杜鳳鳳, 劉曉靜, 常雅軍, 等. 基于SSR標記的荷花品種遺傳多樣性及群體結構分析[J]. 植物資源與環境學報, 2016, 25(1): 9-16.
[11] 楊郭陽. 荷花遺傳連鎖圖譜的構建及株型相關性狀QTLs分析[D]. 武漢: 華中農業大學, 2018.
[12] Yeh F C, Yang R C, Boyle T. Popgene Version 1.31 Quick User Guide[M]. Canada: University of Alberta, and Centre for International Forestry Research, 1999.
[13] Liu K, Muse S V. PowerMarker: An integrated analysis environment for genetic marker analysis[J]. Bioinformatics, 2005, 21(9): 2128-2129.
[14] Evanno G, Regnaut S, Goudet J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study[J]. Molecular Ecology, 2005, 14(8): 2611-2620.
[15] Gaut B S, Long A D. The lowdown on linkage disequilibrium[J]. Plant Cell, 2003, 15(7): 1502-1506.
[16] Flint-Garcia S A, Thornsberry J M, Buckler E S. Struc-ture of linkage disequilibrium in plants[J]. Annual Review of Plant Biology, 2003, 54: 357-374.
[17] Bradbury P J, Zhang Z W, Kroon D E, et al. TASSEL: Software for association mapping of complex traits in diverse samples[J]. Bioinformatics, 2007, 23(19): 2633-2635.
[18] Pritchard J K, Stephens M, Donnelly P. Inference of population structure using multilocus genotype data[J]. Genetics, 2000, 155(2): 945-959.
[19] Gawenda I, Schrder-Lorenz A, Debener T. Markers for ornamental traits in Phalaenopsis orchids: Population structure, linkage disequilibrium and association mapping[J]. Molecular Breeding, 2012, 30(1): 305-316.
[20] Song B H, Windsor A J, Schmid K J, et al. Multilocus patterns of nucleotide diversity, population structure and linkage disequilibrium in Boechera stricta, a wild relative of Arabidopsis[J]. Genetics, 2009, 181(3): 1021-1033.
[21] Zhang F, Ge Y Y, Wang W Y, et al. Genetic diversity and population structure of cultivated bromeliad accessions assessed by SRAP markers[J]. Scientia Horticulturae, 2012, 141: 1-6.
[22] Yang M, Zhu L, Xu L, et al. Population structure and association mapping of flower-related traits in lotus (Nelumbo Adans.) accessions[J]. Scientia Horticulturae, 2014, 175: 214-222.
收稿日期? 2019-11-11;修回日期? 2020-03-23
基金項目 ?農業農村部景觀設計重點實驗室開放基金項目(No. KF201802);江蘇省農業科技自主創新資金項目[No. CX(16)1024]。
作者簡介? 楊? 丹(1996—),女,碩士研究生,研究方向:植物資源學。*通信作者(Corresponding author):劉曉靜(LIU Xiaojing),E-mail:liuxiaojingcau@126.com;姚東瑞(YAO Dongrui),E-mail:shuishengzu@126.com。
